Inhaled Argon Impedes Hepatic Regeneration after Ischemia/Reperfusion Injury in Rats

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
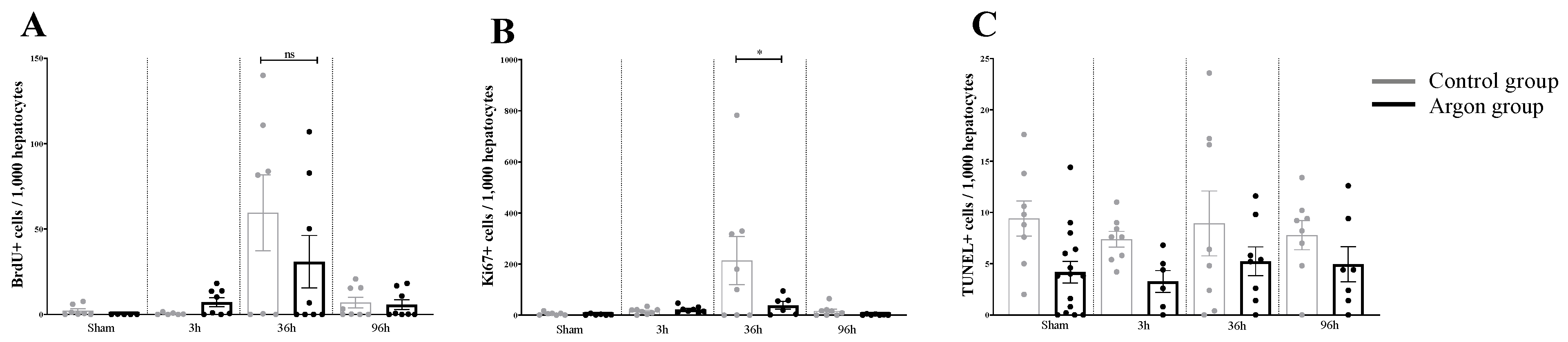
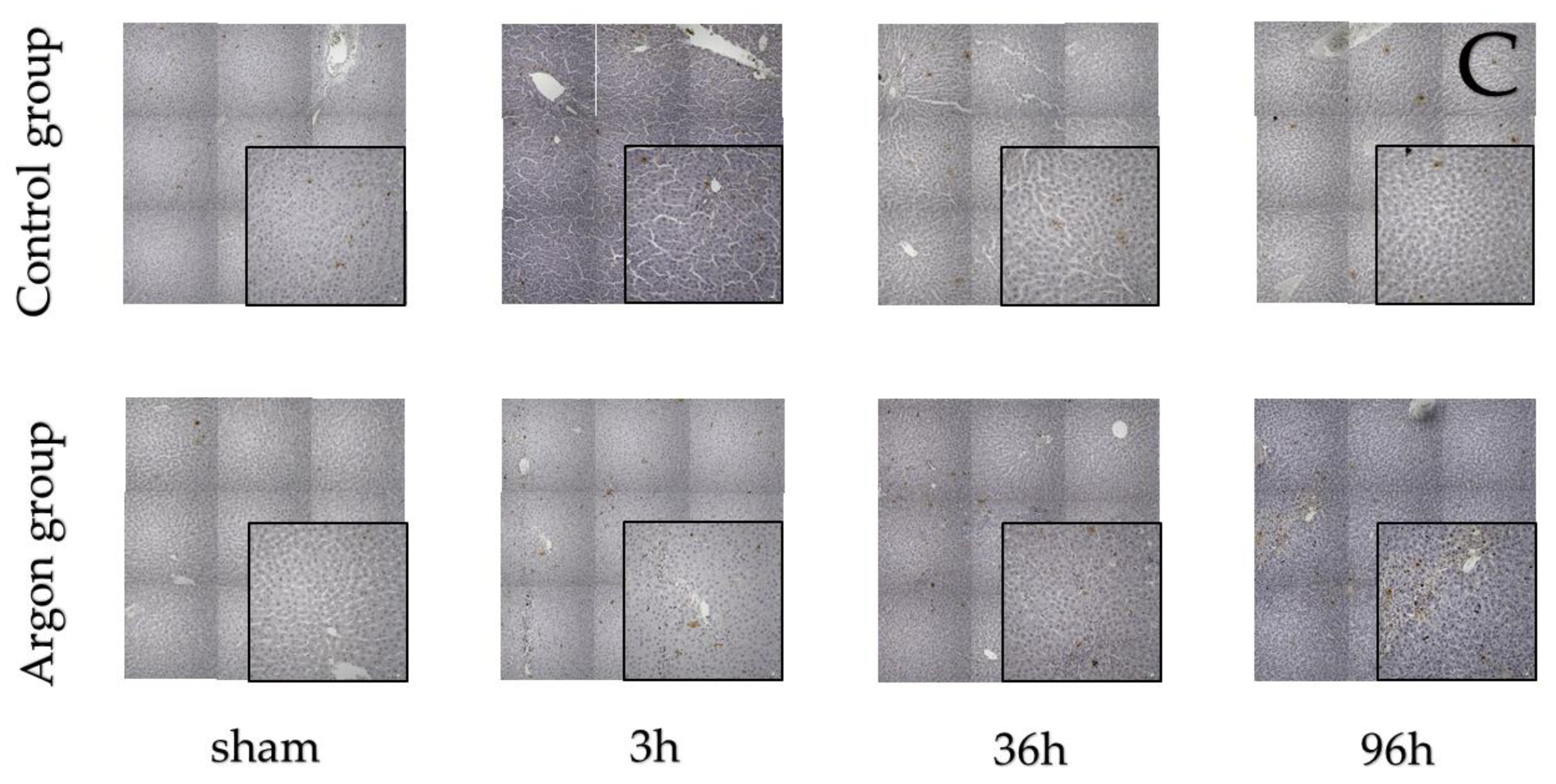
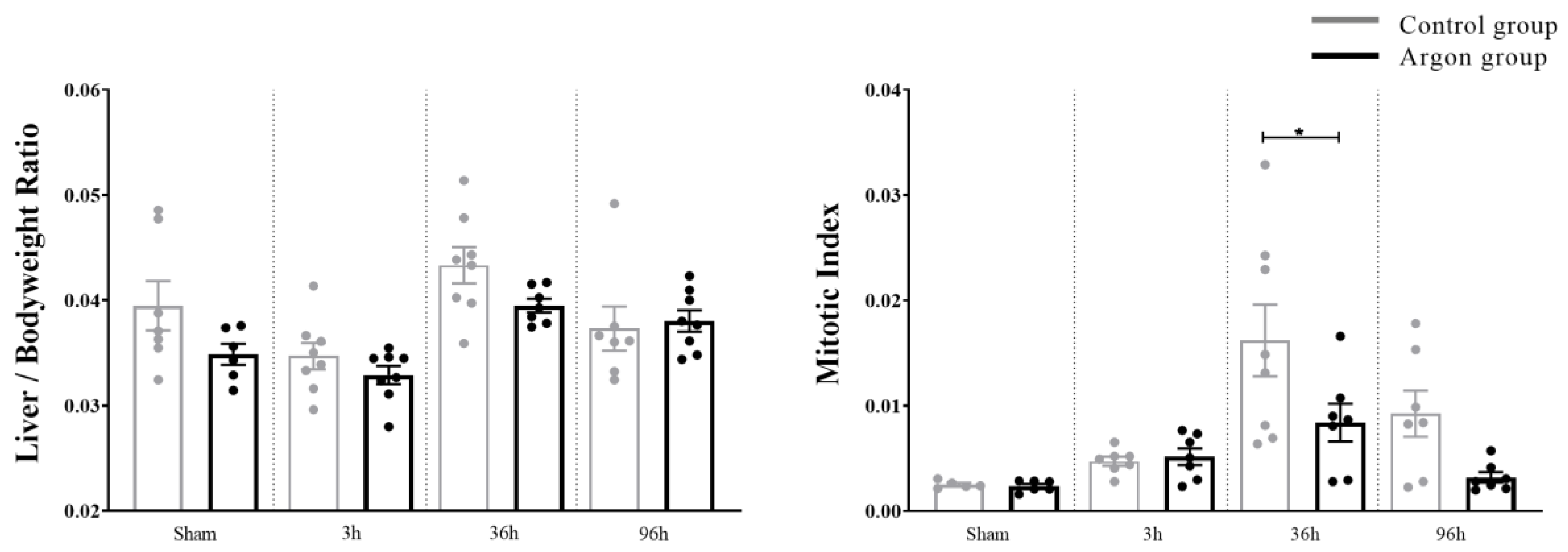
2.1. Effect of Argon on Hepatocytes Proliferation and Apoptosis after I/R
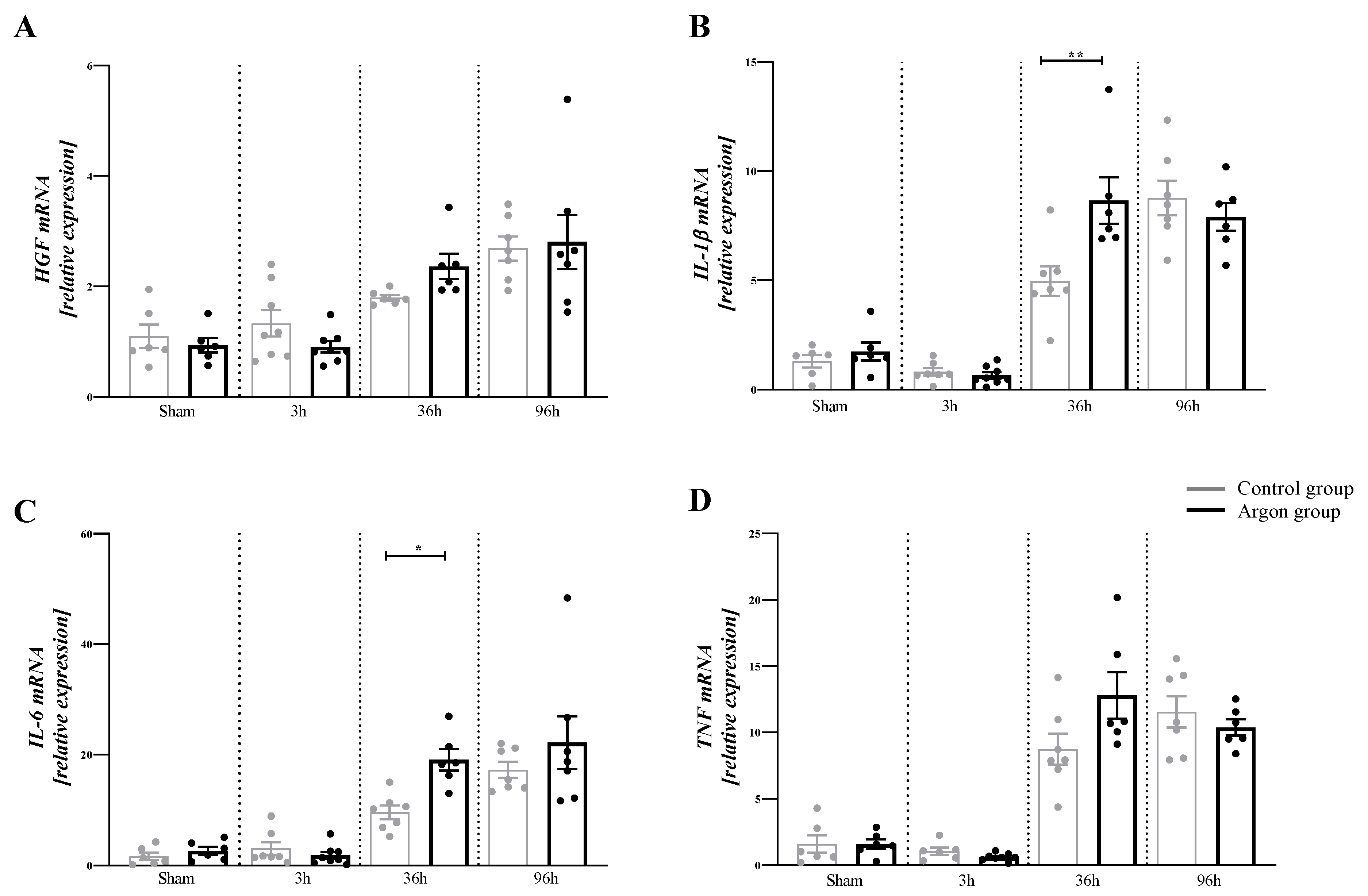
2.2. Argon Modulates the Expression of Cytokines
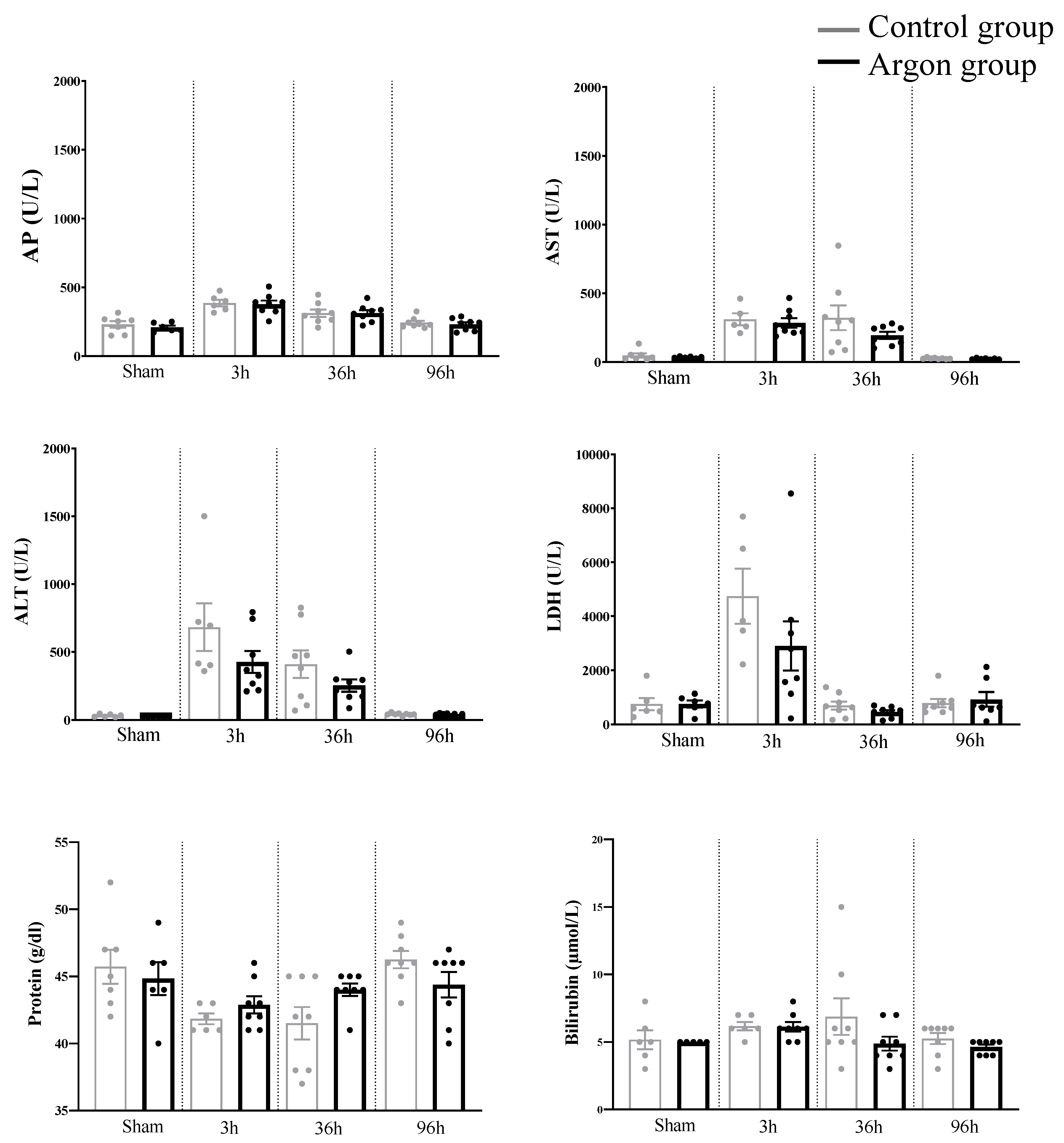
2.3. Plasma Biochemical Data and Parameters of Liver Function (Liver Specific Values)
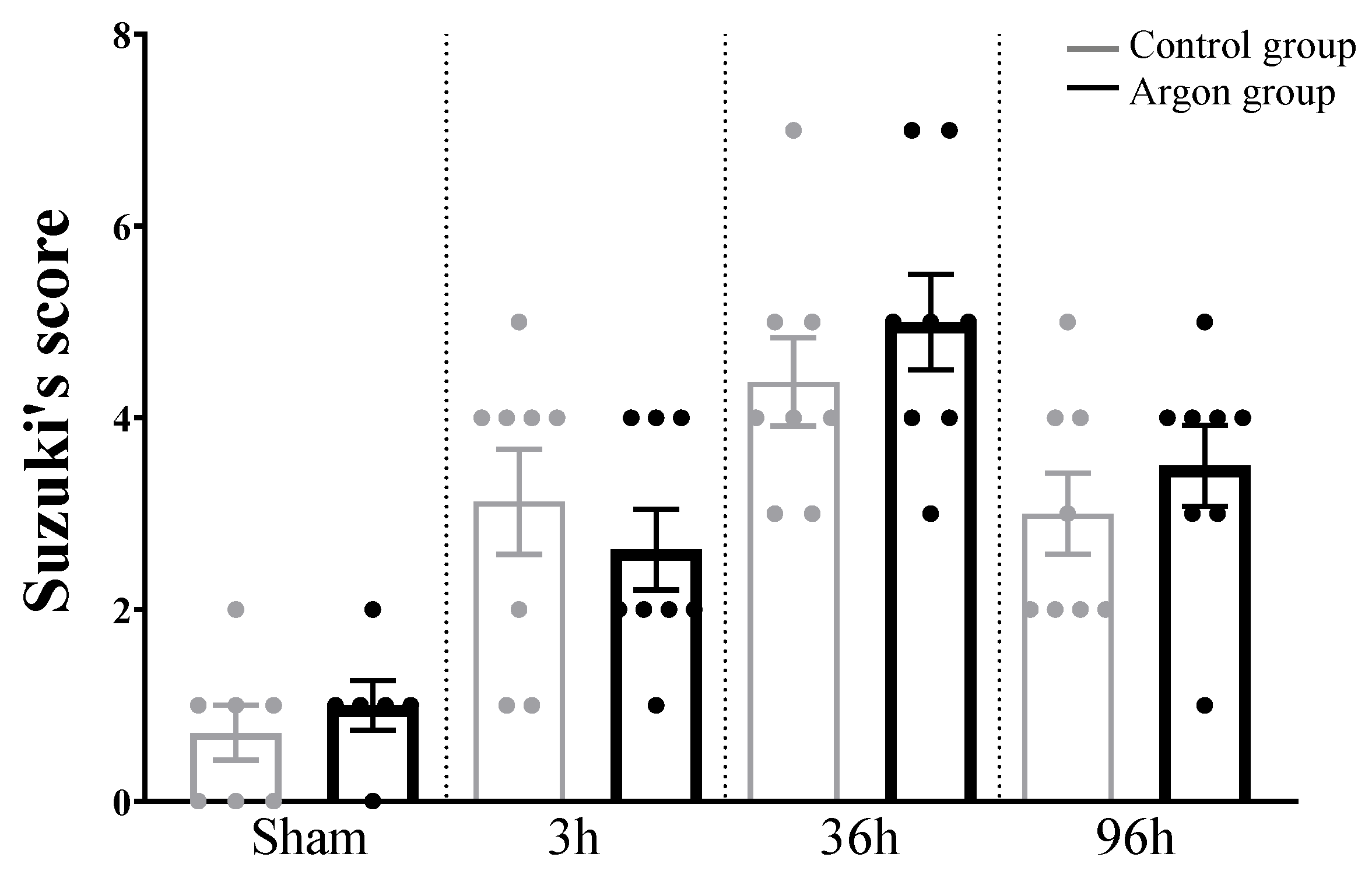
2.4. Liver Injury after IRI Measured by Suzuki’s Score
3. Methods
3.1. Animals and Study Design
3.2. Anesthesia
3.3. Surgical Procedure and Postoperative Care
3.4. Histology
3.5. Immunohistochemistry
3.5.1. Bromodeoxyuridine
3.5.2. KI-67
3.5.3. TUNEL
3.6. RNA Isolation and Quantitative RT-PCR
3.7. Blood Samples
3.8. Quantification and Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Eipel, C.; Glanemann, M.; Nuessler, A.K.; Menger, M.D.; Neuhaus, P.; Vollmar, B. Ischemic preconditioning impairs liver regeneration in extended reduced-size livers. Ann. Surg. 2005, 241, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Lentsch, A.B.; Kato, A.; Yoshidome, H.; McMasters, K.M.; Edwards, M.J. Inflammatory mechanisms and therapeutic strategies for warm hepatic ischemia/reperfusion injury. Hepatology 2000, 32, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Serracino-Inglott, F.; Habib, N.A.; Mathie, R.T. Hepatic ischemia-reperfusion injury. Am. J. Surg. 2001, 181, 160–166. [Google Scholar] [CrossRef]
- Saidi, R.F.; Kenari, S.K. Liver ischemia/reperfusion injury: An overview. J. Investig. Surg. 2014, 27, 366–379. [Google Scholar] [CrossRef]
- Maksan, S.M.; Kraus, T.; Hofmann, W.J.; Mehrabi, A.; Gebhard, M.M.; Herfarth, C.; Klar, E. Hepatocellular injury early after reperfusion is correlated with liver microcirculation and predicts outcome after transplantation. Transpl. Proc. 1998, 30, 3716–3717. [Google Scholar] [CrossRef]
- Casillas-Ramirez, A.; Mosbah, I.B.; Ramalho, F.; Rosello-Catafau, J.; Peralta, C. Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci. 2006, 79, 1881–1894. [Google Scholar] [CrossRef]
- Guimaraes Filho, M.A.; Cortez, E.; Garcia-Souza, E.P.; Soares Vde, M.; Moura, A.S.; Carvalho, L.; Maya, M.C.; Pitombo, M.B. Effect of remote ischemic preconditioning in the expression of IL-6 and IL-10 in a rat model of liver ischemia-reperfusion injury. Acta Cir. Bras. 2015, 30, 452–460. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z.; Feng, N.; Tang, J.; Zhao, X.; Liu, C.; Xu, H.; Zhang, M. Protective effect of propofol preconditioning on ischemia-reperfusion injury in human hepatocyte. J. Thorac. Dis. 2017, 9, 702–710. [Google Scholar] [CrossRef]
- Coburn, M.; Maze, M.; Franks, N.P. The neuroprotective effects of xenon and helium in an in vitro model of traumatic brain injury. Crit. Care Med. 2008, 36, 588–595. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, F.; Liu, G.; Shi, X.; Liu, Y.; Liu, W.; Luo, X.; Sun, X.; Kang, Z. Helium preconditioning attenuates hypoxia/ischemia-induced injury in the developing brain. Brain Res. 2011, 1376, 122–129. [Google Scholar] [CrossRef]
- Veldeman, M.; Coburn, M.; Rossaint, R.; Clusmann, H.; Nolte, K.; Kremer, B.; Hollig, A. Xenon Reduces Neuronal Hippocampal Damage and Alters the Pattern of Microglial Activation after Experimental Subarachnoid Hemorrhage: A Randomized Controlled Animal Trial. Front. Neurol. 2017, 8, 511. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.; Ma, D.; Maze, M.; Franks, N.P. Effects of xenon on in vitro and in vivo models of neuronal injury. Anesthesiology 2002, 96, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Breuer, T.; Emontzpohl, C.; Coburn, M.; Benstoem, C.; Rossaint, R.; Marx, G.; Schalte, G.; Bernhagen, J.; Bruells, C.S.; Goetzenich, A.; et al. Xenon triggers pro-inflammatory effects and suppresses the anti-inflammatory response compared to sevoflurane in patients undergoing cardiac surgery. Crit. Care 2015, 19, 365. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Plitzko, G.; Bicknell, L.; van Caster, P.; Schulz, J.; Barthuber, C.; Preckel, B.; Pannen, B.H.; Bauer, I. Pretreatment with helium does not attenuate liver injury after warm ischemia-reperfusion. Shock 2014, 41, 413–419. [Google Scholar] [CrossRef]
- Reinelt, H.; Marx, T.; Kotzerke, J.; Topalidis, P.; Luederwald, S.; Armbruster, S.; Schirmer, U.; Schmidt, M. Hepatic function during xenon anesthesia in pigs. Acta Anaesthesiol. Scand. 2002, 46, 713–716. [Google Scholar] [CrossRef]
- Wilke, H.J.; Moench, C.; Lotz, G.; Bechstein, W.; Zacharowski, K. Xenon anesthesia for liver transplant surgery: A report of four cases. Transplant. Proc. 2011, 43, 2683–2686. [Google Scholar] [CrossRef]
- Irani, Y.; Pype, J.L.; Martin, A.R.; Chong, C.F.; Daniel, L.; Gaudart, J.; Ibrahim, Z.; Magalon, G.; Lemaire, M.; Hardwigsen, J. Noble gas (argon and xenon)-saturated cold storage solutions reduce ischemia-reperfusion injury in a rat model of renal transplantation. Nephron. Extra 2011, 1, 272–282. [Google Scholar] [CrossRef]
- Alderliesten, T.; Favie, L.M.A.; Neijzen, R.W.; Auwärter, V.; Nijboer, C.H.A.; Marges, R.E.J.; Rademaker, C.M.A.; Kempf, J.; van Bel, F.; Groenendaal, F. Neuroprotection by argon ventilation after perinatal asphyxia: A safety study in newborn piglets. PLoS ONE 2014, 9, e113575. [Google Scholar] [CrossRef]
- Zhuang, L.; Yang, T.; Zhao, H.; Fidalgo, A.R.; Vizcaychipi, M.P.; Sanders, R.D.; Yu, B.; Takata, M.; Johnson, M.R.; Ma, D. The protective profile of argon, helium, and xenon in a model of neonatal asphyxia in rats. Crit. Care Med. 2012, 40, 1724–1730. [Google Scholar] [CrossRef]
- Fahlenkamp, A.V.; Rossaint, R.; Coburn, M. Neuroprotection by noble gases: New developments and insights. Anaesthesist 2015, 64, 855–858. [Google Scholar] [CrossRef]
- Pagel, P.S.; Krolikowski, J.G.; Shim, Y.H.; Venkatapuram, S.; Kersten, J.R.; Weihrauch, D.; Warltier, D.C.; Pratt, P.F. Noble gases without anesthetic properties protect myocardium against infarction by activating prosurvival signaling kinases and inhibiting mitochondrial permeability transition in vivo. Anesth. Analg. 2007, 105, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Jawad, N.; Rizvi, M.; Gu, J.; Adeyi, O.; Tao, G.; Maze, M.; Ma, D. Neuroprotection (and lack of neuroprotection) afforded by a series of noble gases in an in vitro model of neuronal injury. Neurosci. Lett. 2009, 460, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Z.W.; Wang, Z.; Li, J.Q.; Chen, G. The role of argon in stroke. Med. Gas Res. 2018, 8, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, F.; Schallner, N.; Coburn, M.; Loop, T.; Lagrèze, W.A.; Biermann, J.; Goebel, U. Argon inhalation attenuates retinal apoptosis after ischemia/reperfusion injury in a time- and dose-dependent manner in rats. PLoS ONE 2014, 9, e115984. [Google Scholar] [CrossRef]
- Ulmer, T.F.; Fragoulis, A.; Dohmeier, H.; Kroh, A.; Andert, A.; Stoppe, C.; Alizai, H.; Klink, C.; Coburn, M.; Neumann, U.P. Argon Delays Initiation of Liver Regeneration after Partial Hepatectomy in Rats. Eur. Surg. Res. 2017, 58, 204–215. [Google Scholar] [CrossRef]
- Ryang, Y.-M.M.; Fahlenkamp, A.V.; Rossaint, R.; Wesp, D.; Loetscher, P.D.; Beyer, C.; Coburn, M. Neuroprotective effects of argon in an in vivo model of transient middle cerebral artery occlusion in rats. Crit. Care Med. 2011, 39, 1448–1453. [Google Scholar] [CrossRef]
- Sun, P.; Lu, Y.X.; Cheng, D.; Zhang, K.; Zheng, J.; Liu, Y.; Wang, X.; Yuan, Y.F.; Tang, Y.D. Monocyte Chemoattractant Protein-Induced Protein 1 Targets Hypoxia-Inducible Factor 1alpha to Protect Against Hepatic Ischemia/Reperfusion Injury. Hepatology 2018, 68, 2359–2375. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Jaeschke, H. Molecular mechanisms of hepatic ischemia-reperfusion injury and preconditioning. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G15–G26. [Google Scholar] [CrossRef]
- Wanner, G.A.; Ertel, W.; Müller, P.; Höfer, Y.; Leiderer, R.; Menger, M.D.; Messmer, K. Liver ischemia and reperfusion induces a systemic inflammatory response through Kupffer cell activation. Shock 1996, 5, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Gracia-Sancho, J.; Peralta, C. Inflammasome-Mediated Inflammation in Liver Ischemia-Reperfusion Injury. Cells 2019, 8, 1131. [Google Scholar] [CrossRef] [PubMed]
- Shito, M.; Wakabayashi, G.; Ueda, M.; Shimazu, M.; Shirasugi, N.; Endo, M.; Mukai, M.; Kitajima, M. Interleukin 1 receptor blockade reduces tumor necrosis factor production, tissue injury, and mortality after hepatic ischemia-reperfusion in the rat. Transplantation 1997, 63, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Kamo, N.; Ke, B.; Ghaffari, A.A.; Shen, X.D.; Busuttil, R.W.; Cheng, G.; Kupiec-Weglinski, J.W. ASC/caspase-1/IL-1beta signaling triggers inflammatory responses by promoting HMGB1 induction in liver ischemia/reperfusion injury. Hepatology 2013, 58, 351–362. [Google Scholar] [CrossRef]
- Teoh, N.C.; Farrell, G.C. Hepatic ischemia reperfusion injury: Pathogenic mechanisms and basis for hepatoprotection. J. Gastroenterol. Hepatol. 2003, 18, 891–902. [Google Scholar] [CrossRef]
- Jaeschke, H.; Hasegawa, T. Role of neutrophils in acute inflammatory liver injury. Liver Int. 2006, 26, 912–919. [Google Scholar] [CrossRef]
- Kato, A.; Gabay, C.; Okaya, T.; Lentsch, A.B. Specific role of interleukin-1 in hepatic neutrophil recruitment after ischemia/reperfusion. Am. J. Pathol. 2002, 161, 1797–1803. [Google Scholar] [CrossRef]
- Shirasugi, N.; Wakabayashi, G.; Shimazu, M.; Shito, M.; Kawachi, S.; Kitajima, M. Interleukin-1 receptor blockade attenuates oxygen-derived free radical production and microcirculatory disturbances in ischemia/reperfusion injury in the liver. Transpl. Proc. 1997, 29, 371–373. [Google Scholar] [CrossRef]
- Sultan, M.; Ben-Ari, Z.; Masoud, R.; Pappo, O.; Harats, D.; Kamari, Y.; Safran, M. Interleukin-1alpha and Interleukin-1beta play a central role in the pathogenesis of fulminant hepatic failure in mice. PLoS ONE 2017, 12, e0184084. [Google Scholar] [CrossRef]
- Iracheta-Vellve, A.; Petrasek, J.; Gyogyosi, B.; Bala, S.; Csak, T.; Kodys, K.; Szabo, G. Interleukin-1 inhibition facilitates recovery from liver injury and promotes regeneration of hepatocytes in alcoholic hepatitis in mice. Liver Int. 2017, 37, 968–973. [Google Scholar] [CrossRef]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. Hepatology 2006, 43, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Camargo, C.A.; Madden, J.F.; Gao, W.; Selvan, R.S.; Clavien, P.A. Interleukin-6 protects liver against warm ischemia/reperfusion injury and promotes hepatocyte proliferation in the rodent. Hepatology 1997, 26, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Clavien, P.A. IL-6, a key cytokine in liver regeneration. Hepatology 1997, 25, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Teoh, N.; Field, J.; Farrell, G. Interleukin-6 is a key mediator of the hepatoprotective and pro-proliferative effects of ischaemic preconditioning in mice. J. Hepatol. 2006, 45, 20–27. [Google Scholar] [CrossRef]
- Klein, C.; Wustefeld, T.; Assmus, U.; Roskams, T.; Rose-John, S.; Muller, M.; Manns, M.P.; Ernst, M.; Trautwein, C. The IL-6-gp130-STAT3 pathway in hepatocytes triggers liver protection in T cell-mediated liver injury. J. Clin. Investig. 2005, 115, 860–869. [Google Scholar] [CrossRef]
- Blindenbacher, A.; Wang, X.; Langer, I.; Savino, R.; Terracciano, L.; Heim, M.H. Interleukin 6 is important for survival after partial hepatectomy in mice. Hepatology 2003, 38, 674–682. [Google Scholar] [CrossRef]
- Sun, R.; Jaruga, B.; Kulkarni, S.; Sun, H.; Gao, B. IL-6 modulates hepatocyte proliferation via induction of HGF/p21cip1: Regulation by SOCS3. Biochem. Biophys. Res. Commun. 2005, 338, 1943–1949. [Google Scholar] [CrossRef]
- Wüstefeld, T.; Rakemann, T.; Kubicka, S.; Manns, M.P.; Trautwein, C. Hyperstimulation with interleukin 6 inhibits cell cycle progression after hepatectomy in mice. Hepatology 2000, 32, 514–522. [Google Scholar] [CrossRef]
- Gujral, J.S.; Bucci, T.J.; Farhood, A.; Jaeschke, H. Mechanism of cell death during warm hepatic ischemia-reperfusion in rats: Apoptosis or necrosis? Hepatology 2001, 33, 397–405. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmitz, S.M.; Dohmeier, H.; Stoppe, C.; Alizai, P.H.; Schipper, S.; Neumann, U.P.; Coburn, M.; Ulmer, T.F. Inhaled Argon Impedes Hepatic Regeneration after Ischemia/Reperfusion Injury in Rats. Int. J. Mol. Sci. 2020, 21, 5457. https://doi.org/10.3390/ijms21155457
Schmitz SM, Dohmeier H, Stoppe C, Alizai PH, Schipper S, Neumann UP, Coburn M, Ulmer TF. Inhaled Argon Impedes Hepatic Regeneration after Ischemia/Reperfusion Injury in Rats. International Journal of Molecular Sciences. 2020; 21(15):5457. https://doi.org/10.3390/ijms21155457
Chicago/Turabian StyleSchmitz, Sophia M., Henriette Dohmeier, Christian Stoppe, Patrick H. Alizai, Sandra Schipper, Ulf P. Neumann, Mark Coburn, and Tom F. Ulmer. 2020. "Inhaled Argon Impedes Hepatic Regeneration after Ischemia/Reperfusion Injury in Rats" International Journal of Molecular Sciences 21, no. 15: 5457. https://doi.org/10.3390/ijms21155457
APA StyleSchmitz, S. M., Dohmeier, H., Stoppe, C., Alizai, P. H., Schipper, S., Neumann, U. P., Coburn, M., & Ulmer, T. F. (2020). Inhaled Argon Impedes Hepatic Regeneration after Ischemia/Reperfusion Injury in Rats. International Journal of Molecular Sciences, 21(15), 5457. https://doi.org/10.3390/ijms21155457