Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases?

 , , and
, , and 
Abstract
1. Introduction
1.1. Characteristics and Isoforms of Adiponectin in Liver Diseases
1.2. Source of Adiponectin in Liver Diseases
1.3. Adiponectin Receptors in Liver Diseases
2. Adiponectin Effects on Hepatic Damage and Regenerative Failure Associated with Hepatic I/R
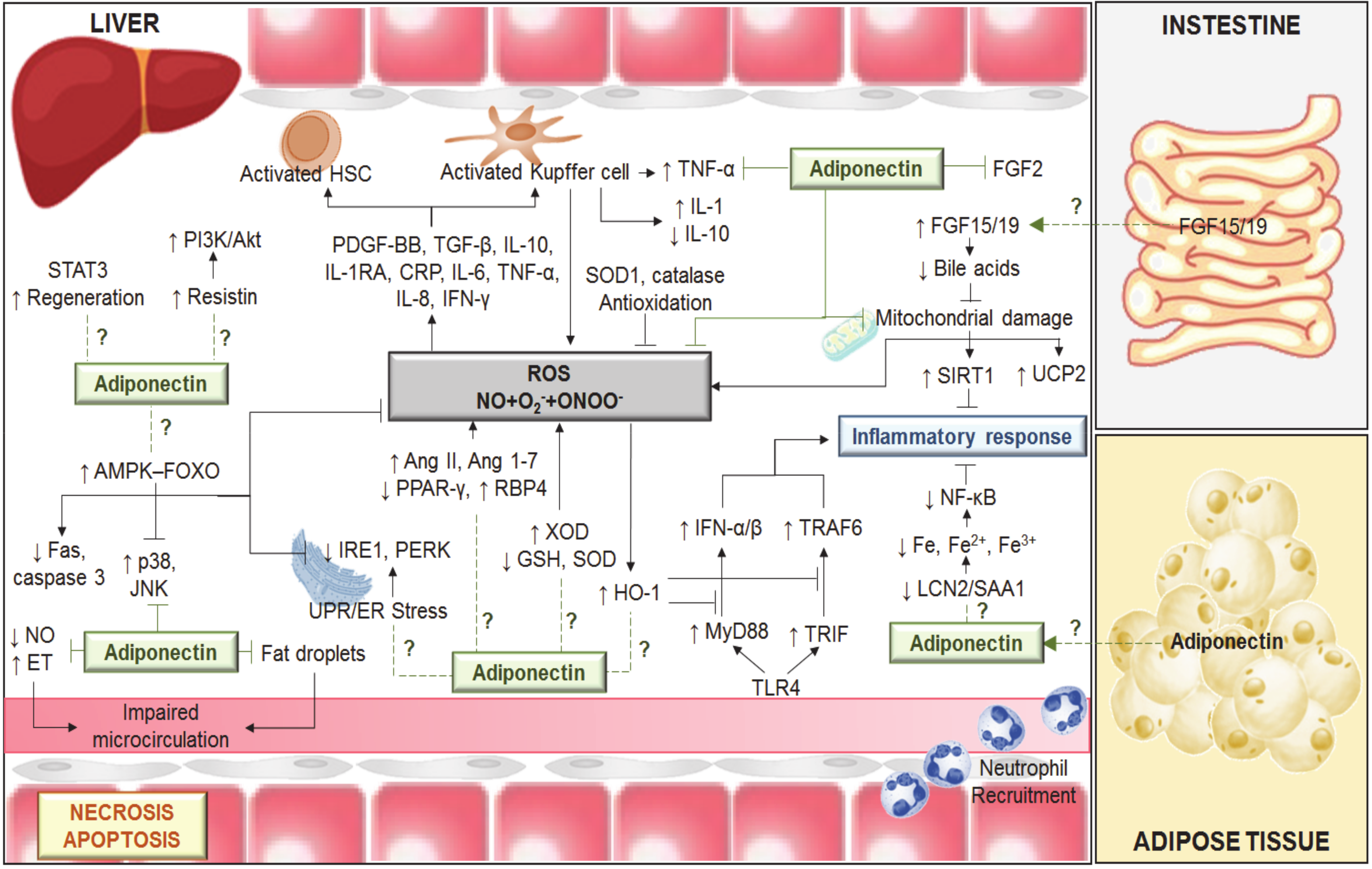
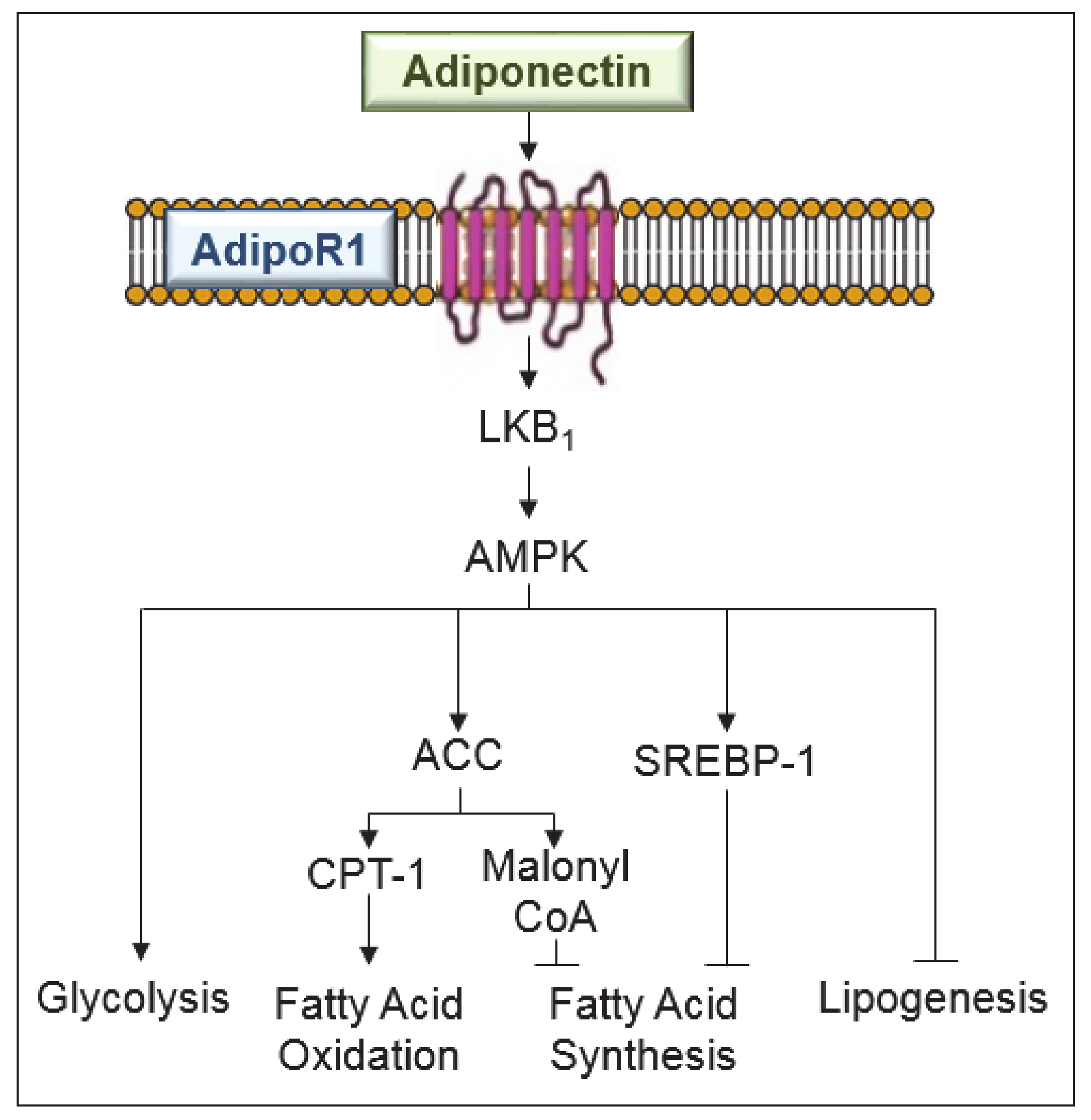
3. Anti-Steatotic Effects of Adiponectin in Liver Diseases
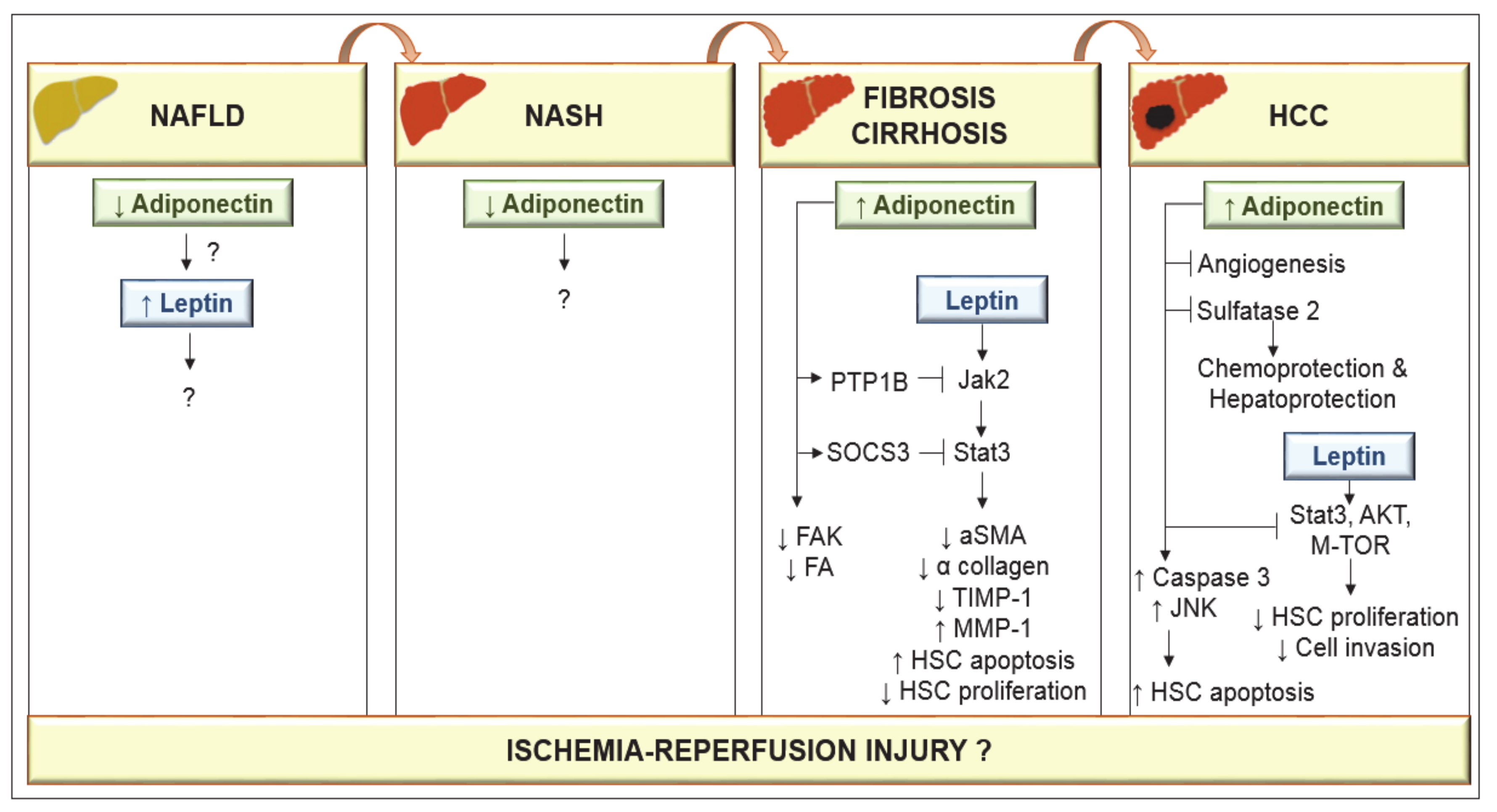
4. Relationship between Adiponectin and Leptin in Fibrogenesis
5. Pharmacological Strategies Regulating the Action of Adiponectin on Hepatic Damage and Regenerative Failure Associated with Hepatic I/R
6. Pharmacological Strategies to Regulate Adiponectin Action in Liver Diseases and the Absence of Hepatic I/R
7. Adiponectin as a Prognostic Factor in Liver Diseases
8. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA carboxylase |
| AdipoQ | Adiponectin, C1Q and collagen domain containing |
| AdipoR1/R2 | Adiponectin receptor type 1/2 |
| And-/- | Adiponectin knockout |
| AICAR | Aminoimidazole-4-carboxamide ribonucleoside |
| Akt | Protein kinase B |
| ALD | Alcoholic liver disease |
| ALT | Alanine aminotransferase |
| AMPK | Adenosine monophosphate-activated protein kinase |
| Ang | Angiotensin |
| APPL1 | Adaptor protein |
| ATF6 | Activating transcription factor 6 |
| ATP | Adenosine triphosphate |
| CCL2 | C-C motif chemokine ligand 2 |
| CHIP | C-terminus of Hsc70-interacting protein |
| CPT-1 | Carnitine palmitoyl- transferase 1 |
| CRP | C-reactive protein |
| CXCL-10 | C-X-C motif chemokine ligand 10 |
| Cyp7a1 | Cholesterol 7 alpha-hydroxylase |
| EC | Endothelial cell |
| eNOS | Endothelial nitric oxide synthase |
| ER | Endoplasmic reticulum |
| ET | Endothelin |
| FA | Focal adhesion |
| FAK | Focal adhesion kinase |
| Fas | Fas-associated death domain |
| Fe | Iron |
| FGF2 | Fibroblast growth factor 2 |
| FGF15/19 | Fibroblast growth factor 15/19 |
| FOXO | Forkhead box protein O |
| FXR | Farsenoid-X receptor |
| gAcrp30 | Globular adiponectin |
| GSH | Glutathione |
| h | Hours |
| HCC | Hepatocellular carcinoma |
| HFD | High fat diet |
| HMW | High molecular weight |
| HO-1 | Heme oxygenase-1 |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| HSC | Hepatic stellate cells |
| I/R | Ischemia/reperfusion |
| ICAM | Intercellular cell adhesion molecule |
| IFN | Interferon |
| IL | Interleukin |
| IRE1 | Inositol-requiring enzyme 1 |
| Jak2 | Janus kinase 2 |
| JNK | c-Jun N-terminal kinase |
| KC | Kupffer cells |
| Ki-67 | Antigen Ki-67, a marker of proliferation |
| KO | Knockout |
| LCN2/SAA1 | Lipocalin-2/serum amyloid A1 |
| LKB1 | Liver kinase B1 |
| LMW | Low molecular weight |
| LT | Liver transplantation |
| MCD | Methionine-choline deficiency |
| min | Minutes |
| mLipin-1 | Myeloid cell-specific lipin-1 |
| MMP-1 | Matrix metalloproteinase 1 |
| MMW | Middle molecular weight |
| mNT | MitoNEET |
| MPO | Myeloperoxidase |
| m-TOR | Mammalian target of rapamycin |
| MyD88 | Myeloid differentiation primary response gene 88 |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| O2 | Superoxide |
| ONOO- | Peroxynitrite |
| p38 | Mitogen-activated protein kinase p38 |
| PDGF-BB | Platelet-derived growth factor-BB |
| PERK | Protein kinase-like endoplasmic reticulum kinase |
| PI3K | Phosphoinositide 3-kinase |
| PPAR | Peroxisome proliferator-activated receptor |
| PTP1B | Protein tyrosine phosphatase 1B |
| RBP4 | Retinol binding protein 4 |
| ROS | Reactive oxygen species |
| siRNA | Silent small interfering RNA |
| SIRT1 | Sirtuin-1 |
| α-SMA | α-smooth muscle actin |
| SOCS3 | Suppressors of cytokine signaling 3 |
| SOD | Superoxide dismutase |
| SREBP-1c | Sterol regulatory element-binding protein 1c |
| STAT3 | Signal transducer and activator of transcription 3 |
| TGF-β | Tumor growth factor beta |
| TIMP-1 | Tissue inhibitor of metalloproteinase 1 |
| TLR4 | Toll-like receptor 4 |
| TNF-α | Tumor necrosis factor alpha |
| TRAF6 | TNF receptor-associated factor 6 |
| TRIF | TIR-domain-containing adapter-inducing interferon-β |
| UPC2 | Uncoupling protein 2 |
| UPR | Unfolded protein response |
| X/XOD | Xanthine/Xanthine oxidase |
| XOD | Xanthine oxidase |
References
- Nocito, A.; El-Badry, A.M.; Clavien, P.A. When is steatosis too much for transplantation? J. Hepatol. 2006, 45, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Mercado, A.I.; Gulfo, J.; Romero-Gómez, M.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. Use of steatotic grafts in liver transplantation: Current status. Liver Transpl. 2019, 25, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Peralta, C.; Jiménez-Castro, M.B.; Gracia-Sancho, J. Hepatic ischemia and reperfusion injury: Effects on the liver sinusoidal milieu. J. Hepatol. 2013, 59, 1094–1106. [Google Scholar] [CrossRef] [PubMed]
- Tsutamoto, T.; Tanaka, T.; Sakai, H.; Ishikawa, C.; Fujii, M.; Yamamoto, T.; Horie, M. Total and high molecular weight adiponectin, haemodynamics, and mortality in patients with chronic heart failure. Eur. Heart J. 2007, 28, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Cheng, K.K.; Vanhoutte, P.M.; Lam, K.S.; Xu, A. Vascular effects of adiponectin: Molecular mechanisms and potential therapeutic intervention. Clin. Sci. 2008, 114, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Bugianesi, E.; Frystyk, J.; Tarnow, L.; Flyvbjerg, A.; Marchesini, G. Adiponectin isoforms, insulin resistance and liver histology in nonalcoholic fatty liver disease. Dig. Liver Dis. 2011, 43, 73–77. [Google Scholar] [CrossRef]
- Adachi, M.; Brenner, D.A. High molecular weight adiponectin inhibits proliferation of hepatic stellate cells via activation of adenosine monophosphate-activated protein kinase. Hepatology 2008, 47, 677–685. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, G.; Wong, W.P.; Vliegenthart, J.F.; Gerwig, G.J.; Lam, K.S.; Cooper, G.J.; Xu, A. Proteomic and functional characterization of endogenous adiponectin purified from fetal bovine serum. Proteomics 2004, 4, 3933–3942. [Google Scholar] [CrossRef]
- Massip-Salcedo, M.; Zaouali, M.A.; Padrissa-Altés, S.; Casillas-Ramirez, A.; Rodés, J.; Roselló-Catafau, J.; Peralta, C. Activation of peroxisome proliferator-activated receptor-alpha inhibits the injurious effects of adiponectin in rat steatotic liver undergoing ischemia-reperfusion. Hepatology 2008, 47, 461–472. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 1996, 271, 10697–10703. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Bunt, J.C.; Salbe, A.D.; Funahashi, T.; Matsuzawa, Y.; Tataranni, P.A. Plasma adiponectin concentrations in children: Relationships with obesity and insulinemia. J. Clin. Endocrinol. Metab. 2002, 87, 4652–4656. [Google Scholar] [CrossRef]
- Ostrowska, L.; Fiedorczuk, J.; Adamska, E. Effect of diet and other factors on serum adiponectin concentrations in 49 patients with type 2 diabetes. Roczniki Państwowego Zakładu Higieny 2013, 64, 61–66. [Google Scholar] [PubMed]
- Yatagai, T.; Nagasaka, S.; Taniguchi, A.; Fukushima, M.; Nakamura, T.; Kuroe, A.; Nakai, Y.; Ishibashi, S. Hypoadiponectinemia is associated with visceral fat accumulation and insulin resistance in Japanese men with type 2 diabetes mellitus. Metabolism 2003, 52, 1274–1278. [Google Scholar] [CrossRef]
- Bidulescu, A.; Liu, J.; Hickson, D.A.; Hairston, K.G.; Fox, E.R.; Arnett, D.K.; Sumner, A.E.; Taylor, H.A.; Gibbons, G.H. Gender differences in the association of visceral and subcutaneous adiposity with adiponectin in African americans: The jackson heart study. BMC Cardiovasc. Disord. 2013, 13, 9–19. [Google Scholar] [CrossRef]
- Jiménez-Castro, M.B.; Casillas-Ramírez, A.; Mendes-Braz, M.; Massip-Salcedo, M.; Gracia-Sancho, J.; Elias-Miró, M.; Rodés, J.; Peralta, C. Adiponectin and resistin protect steatotic livers undergoing transplantation. J. Hepatol. 2013, 59, 1208–1214. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Lin, Y.S.; Yeh, T.S.; Cheong, C.F.; Chang, C.H.; Chen, T.C.; Chen, M.F. Disrupted hepatic adiponectin signaling impairs liver regeneration of steatotic rats. Chang Gung Med. J. 2011, 34, 248–259. [Google Scholar]
- Deepa, S.S.; Dong, L.Q. APPL1: Role in adiponectin signaling and beyond. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E22–E36. [Google Scholar] [CrossRef]
- Holmes, R.M.; Yi, Z.; De Filippis, E.; Berria, R.; Shahani, S.; Sathyanarayana, P.; Sherman, V.; Fujiwara, K.; Meyer, C.; Christ-Roberts, C.; et al. Increased abundance of the adaptor protein containing pleckstrin homology domain, phosphotyrosine binding domain and leucine zipper motif (APPL1) in patients with obesity and type 2 diabetes: Evidence for altered adiponectin signalling. Diabetologia 2011, 54, 2122–2131. [Google Scholar] [CrossRef]
- Tarantino, G.; Savastano, S.; Colao, A. Hepatic steatosis, low-grade chronic inflammation and hormone/growth factor/adipokine imbalance. World J. Gastroenterol. 2010, 16, 4773–4783. [Google Scholar] [CrossRef]
- Ma, H.; Gomez, V.; Lu, L.; Yang, X.; Wu, X.; Xiao, S.Y. Expression of adiponectin and its receptors in livers of morbidly obese patients with non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2009, 24, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, M.; Cecchetti, F.; Anselmino, M.; Mancini, E.; Marchetti, G.; Bonotti, A.; Baldi, S.; Solito, B.; Giannetti, M.; Pinchera, A.; et al. Pattern of expression of adiponectin receptors in human liver and its relation to nonalcoholic steatohepatitis. Obes. Surg. 2009, 19, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Tiniakos, D.G. Nonalcoholic fatty liver disease/nonalcoholic steatohepatitis: Histological diagnostic criteria and scoring systems. Eur. J. Gastroenterol. Hepatol. 2010, 22, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Vuppalanchi, R.; Marri, S.; Kolwankar, D.; Considine, R.V.; Chalasani, N. Is adiponectin involved in the pathogenesis of nonalcoholic steatohepatitis? A preliminary human study. J. Clin. Gastroenterol. 2005, 39, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Hirata, T.; Wada, A.; Kawamura, N.; Yamaguchi, Y.; Fujimura, K.; Ueda, T.; Yurugi, Y.; Soya, H.; Nishibe, S. Chronic administration of Eucommia leaf stimulates metabolic function of rats across several organs. Br. J. Nutr. 2010, 104, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Baranova, A.; Gowder, S.J.; Schlauch, K.; Elariny, H.; Collantes, R.; Afendy, A.; Ong, J.P.; Goodman, Z.; Chandhoke, V.; Younossi, Z.M. Gene expression of leptin, resistin, and adiponectin in the white adipose tissue of obese patients with non-alcoholic fatty liver disease and insulin resistance. Obes. Surg. 2006, 16, 1118–1125. [Google Scholar] [CrossRef]
- Jiang, L.L.; Li, L.; Hong, X.F.; Li, Y.M.; Zhang, B.L. Patients with nonalcoholic fatty liver disease display increased serum resistin levels and decreased adiponectin levels. Eur. J. Gastroenterol. Hepatol. 2009, 21, 662–666. [Google Scholar] [CrossRef]
- Jiménez-Castro, M.B.; Meroño, N.; Mendes-Braz, M.; Gracia-Sancho, J.; Martínez-Carreres, L.; Cornide-Petronio, M.E.; Casillas-Ramirez, A.; Rodés, J.; Peralta, C. The effect of brain death in rat steatotic and non-steatotic liver transplantation with previous ischemic preconditioning. J. Hepatol. 2015, 62, 83–91. [Google Scholar] [CrossRef]
- Ezaki, H.; Yoshida, Y.; Saji, Y.; Takemura, T.; Fukushima, J.; Matsumoto, H.; Kamada, Y.; Wada, A.; Igura, T.; Kihara, S.; et al. Delayed liver regeneration after partial hepatectomy in adiponectin knockout mice. Biochem. Biophys. Res. Commun. 2009, 378, 68–72. [Google Scholar] [CrossRef]
- Shu, R.Z.; Zhang, F.; Wang, F.; Feng, D.C.; Li, X.H.; Ren, W.H.; Wu, X.L.; Yang, X.; Liao, X.D.; Huang, L.; et al. Adiponectin deficiency impairs liver regeneration through attenuating STAT3 phosphorylation in mice. Lab. Invest. 2009, 89, 1043–1052. [Google Scholar] [CrossRef]
- Correnti, J.M.; Cook, D.; Aksamitiene, E.; Swarup, A.; Ogunnaike, B.; Vadigepalli, R.; Hoeck, J.B. Adiponectin fine-tuning of liver regeneration dynamics revealed through cellular network modelling. J. Physiol. 2015, 593, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Sturm, J.; Keese, M.; Zhang, H.; Bönninghoff, R.; Magdeburg, R.; Vajkoczy, P.; Dono, R.; Zeller, R.; Gretz, N. Liver regeneration in FGF-2-deficient mice: VEGF acts as potential functional substitute for FGF-2. Liver Int. 2004, 24, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Takehara, T.; Hayashi, N. Adipocytokines and liver disease. J. Gastroenterol. 2008, 43, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Awazawa, M.; Ueki, K.; Inabe, K.; Yamauchi, T.; Kaneko, K.; Okazaki, Y.; Bardeesy, N.; Ohnishi, S.; Nagai, R.; Kadowaki, T. Adiponectin suppresses hepatic SREBP1c expression in an AdipoR1/LKB1/AMPK dependent pathway. Biochem. Biophys. Res. Commun. 2009, 382, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Shklyaev, S.; Aslanidi, G.; Tennant, M.; Prima, V.; Kohlbrenner, E.; Kroutov, V.; Campbell-Thompson, M.; Crawford, J.; Shek, E.W.; Scarpace, P.J.; et al. Sustained peripheral expression of transgene adiponectin offsets the development of diet-induced obesity in rats. Proc. Nat. Acad. Sci. USA 2003, 100, 14217–14222. [Google Scholar] [CrossRef]
- Hardie, D.G.; Pan, D.A. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase. Biochem. Soc. Trans. 2002, 30, 1064–1070. [Google Scholar] [CrossRef]
- You, M.; Rogers, C.Q. Adiponectin: A key adipokine in alcoholic fatty liver. Exp. Biol. Med. 2009, 234, 850–859. [Google Scholar] [CrossRef]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Watt, K.D.; Fan, C.; Therneau, T.; Heimbach, J.K.; Seaberg, E.C.; Charlton, M.R. Serum adipokine and inflammatory markers before and after liver transplantation in recipients with major cardiovascular events. Liver Transpl. 2014, 20, 791–797. [Google Scholar] [CrossRef]
- Fernández, L.; Heredia, N.; Grande, L.; Gómez, G.; Rimola, A.; Marco, A.; Gelpí, E.; Roselló-Catafau, J.; Peralta, C. Preconditioning protects liver and lung damage in rat liver transplantation: Role of xanthine/xanthine oxidase. Hepatology 2002, 36, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Peralta, C.; Perales, J.C.; Bartrons, R.; Mitchell, C.; Gilgenkrantz, H.; Xaus, C.; Prats, N.; Fernández, L.; Gelpí, E.; Panés, J.; et al. The combination of ischemic preconditioning and liver Bcl-2 overexpression is a suitable strategy to prevent liver and lung damage after hepatic ischemia-reperfusion. Am. J. Pathol. 2002, 160, 2111–2122. [Google Scholar] [CrossRef][Green Version]
- Saxena, N.K.; Anania, F.A. Adipocytokines and hepatic fibrosis. Trends Endocrinol. Metab. 2015, 26, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Haberl, E.M.; Rein-Fischboeck, L.; Aslanidis, C. Adipokines in liver cirrhosis. Int. J. Mol. Sci. 2017, 18, 1392. [Google Scholar] [CrossRef]
- Kaser, S.; Moschen, A.; Kaser, A.; Ludwiczek, O.; Ebenbichler, C.F.; Vogel, W.; Jaschke, W.; Patsch, J.R.; Tilg, H. Circulating adiponectin reflects severity of liver disease but not insulin sensitivity in liver cirrhosis. J. Intern. Med. 2005, 258, 274–280. [Google Scholar] [CrossRef]
- Tacke, F.; Wüstefeld, T.; Horn, R.; Luedde, T.; Rao, A.S.; Manns, M.P.; Trautwein, C.; Brabant, G. High adiponectin in chronic liver disease and cholestasis suggests biliary route of adiponectin excretion in vivo. J. Hepatol. 2005, 42, 666–673. [Google Scholar] [CrossRef]
- Sohara, N.; Takagi, H.; Kakizaki, S.; Sato, K.; Mori, M. Elevated plasma adiponectin concentrations in patients with liver cirrhosis correlate with plasma insulin levels. Liver Int. 2005, 25, 28–32. [Google Scholar] [CrossRef]
- Adolph, T.E.; Grander, C.; Grabherr, F.; Tilg, H. Adipokines and non-alcoholic fatty liver disease: Multiple interactions. Int. J. Mol. Sci. 2017, 18, 1649. [Google Scholar] [CrossRef]
- Kamada, Y.; Matsumoto, H.; Tamura, S.; Fukushima, J.; Kiso, S.; Fukui, K.; Igura, T.; Maeda, N.; Kihara, S.; Funahashi, T.; et al. Hypoadiponectinemia accelerates hepatic tumor formation in a nonalcoholic steatohepatitis mouse model. J. Hepatol. 2007, 47, 556–564. [Google Scholar] [CrossRef]
- Saxena, N.K.; Fu, P.P.; Nagalingam, A.; Wang, J.; Handy, J.; Cohen, C.; Tighiouart, M.; Sharma, D.; Anania, F.A. Adiponectin modulates C-jun N-terminal kinase and mammalian target of rapamycin and inhibits hepatocellular carcinoma. Gastroenterology 2010, 139, 1762–1773.e1–5. [Google Scholar] [CrossRef]
- Hui, J.M.; Hodge, A.; Farrell, G.C.; Kench, J.G.; Kriketos, A.; George, J. Beyond insulin resistance in NASH: TNF-alpha or adiponectin? Hepatology 2004, 40, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Molnar, C.; Wolf, A.M.; Weiss, H.; Graziadei, I.; Kaser, S.; Ebenbichler, C.F.; Stadlmann, S.; Moser, P.L.; Tilg, H. Effects of weight loss induced by bariatric surgery on hepatic adipocytokine expression. J. Hepatol. 2009, 51, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Al-Gayyar, M.M.; Abbas, A.; Hamdan, A.M. Chemopreventive and hepatoprotective roles of adiponectin (SULF2 inhibitor) in hepatocelluar carcinoma. Biol. Chem. 2016, 397, 257–267. [Google Scholar] [CrossRef]
- Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; Hsueh, W.A. Obesity, inflammation, and cancer. Annu. Rev. Pathol. 2016, 11, 421–449. [Google Scholar] [CrossRef]
- Wang, S.N.; Yeh, Y.T.; Yang, S.F.; Chai, C.Y.; Lee, K.T. Potential role of leptin expression in hepatocellular carcinoma. J. Clin. Pathol. 2006, 59, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.; Yu, Y.D.; Kim, D.S.; Won, N.H. Adiponectin receptor expression predicts favorable prognosis in cases of hepatocellular carcinoma. Pathol. Oncol. Res. 2014, 20, 667–675. [Google Scholar] [CrossRef]
- Wang, S.N.; Yang, S.F.; Tsai, H.H.; Lee, K.T.; Yeh, Y.T. Increased adiponectin associated with poor survival in hepatocellular carcinoma. J. Gastroenterol. 2014, 49, 1342–1351. [Google Scholar] [CrossRef]
- Kasztelan-Szczerbinska, B.; Surdacka, A.; Slomka, M.; Rolinski, J.; Celinski, K.; Smolen, A.; Szczerbinski, M. Association of serum adiponectin, leptin, and resistin concentrations with the severity of liver dysfunction and the disease complications in alcoholic liver disease. Mediat. Inflamm. 2013, 2013, 148526. [Google Scholar] [CrossRef]
- Manieri, E.; Herrera-Melle, L.; Mora, A.; Tomás-Loba, A.; Leiva-Vega, L.; Fernández, D.I.; Rodríguez, E.; Morán, L.; Hernández-Cosido, L.; Torres, J.L.; et al. Adiponectin accounts for gender differences in hepatocellular carcinoma incidence. J. Exp. Med. 2019, 216, 1108–1119. [Google Scholar] [CrossRef]
- Zhang, C.; Liao, Y.; Li, Q.; Chen, M.; Zhao, Q.; Deng, R.; Wu, C.; Yang, A.; Guo, Z.; Wang, D.; et al. Recombinant adiponectin ameliorates liver ischemia reperfusion injury via activating the AMPK/eNOS pathway. PLoS ONE 2013, 8, e66382. [Google Scholar] [CrossRef]
- Ding, W.; Zhang, Q.; Dong, Y.; Ding, N.; Huang, H.; Zhu, X.; Hutchinson, S.; Gao, X.; Zhang, X. Adiponectin protects the rats liver against chronic intermittent hypoxia induced injury through AMP-activated protein kinase pathway. Sci. Rep. 2016, 6, 34151. [Google Scholar] [CrossRef]
- Xia, Y.; Gong, J.P. Impact of recombinant globular adiponectin on early warm ischemia-reperfusion injury in rat bile duct after liver transplantation. Sci. Rep. 2014, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Turmelle, Y.P.; Shikapwashya, O.; Tu, S.; Hruz, P.W.; Yan, Q.; Rudnick, D.A. Rosiglitazone inhibits mouse liver regeneration. FASEB J. 2006, 20, 2609–2611. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Braz, M.; Elias-Miró, M.; Kleuser, B.; Fayyaz, S.; Jiménez-Castro, M.B.; Massip-Salcedo, M.; Gracia-Sancho, J.; Ramalho, F.S.; Rodes, J.; Peralta, C. The effects of glucose and lipids in steatotic and non-steatotic livers in conditions of partial hepatectomy under ischaemia-reperfusion. Liver Int. 2014, 34, e271–e289. [Google Scholar] [CrossRef] [PubMed]
- Parkin, E.; O’Reilly, D.A.; Plumb, A.A.; Manoharan, P.; Rao, M.; Coe, P.; Frystyk, J.; Ammori, B.; de Liguori Carino, N.; Deshpande, R.; et al. Digital histology quantification of intra-hepatic fat in patients undergoing liver resection. Eur. J. Surg. Oncol. 2015, 41, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Roychowdhury, S.; Park, P.H.; Pratt, B.T.; Roger, T.; Nagy, L.E. Adiponectin and heme oxygenase-1 suppress TLR4/MyD88-independent signaling in rat Kupffer cells and in mice after chronic ethanol exposure. J. Immunol. 2010, 185, 4928–4937. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Nio, Y.; Maki, T.; Kobayashi, M.; Takazawa, T.; Iwabu, M.; Okada-Iwabu, M.; Kawamoto, S.; Kubota, N.; Kubota, T.; et al. Targeted disruption of AdipoR1 and AdipoR2 causes abrogation of adiponectin binding and metabolic actions. Nat. Med. 2007, 13, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Grenert, J.P.; Patterson, C.; Correia, M.A. CHIP (-/-)-mouse liver: Adiponectin-AMPK- FOXO-activation overrides CYP2E1-elicited JNK1-activation, delaying onset of NASH: Therapeutic implications. Sci. Rep. 2016, 6, 29423. [Google Scholar] [CrossRef]
- Wang, J.; Kim, C.; Jogasuria, A.; Han, Y.; Hu, X.; Wu, J.; Shen, H.; Chrast, R.; Finck, B.N.; You, M. Myeloid cell-specific lipin-1 deficiency stimulates endocrine adiponectin-FGF15 axis and ameliorates ethanol-induced liver injury in mice. Sci. Rep. 2016, 6, 34117. [Google Scholar] [CrossRef]
- Hu, X.; Jogasuria, A.; Wang, J.; Kim, C.; Han, Y.; Shen, H.; Wu, J.; You, M. MitoNEET deficiency alleviates experimental alcoholic steatohepatitis in mice by stimulating endocrine adiponectin-Fgf15 Axis. J. Biol. Chem. 2016, 291, 22482–22495. [Google Scholar] [CrossRef]
- Bechmann, L.P.; Kocabayoglu, P.; Sowa, J.P.; Sydor, S.; Best, J.; Schlattjan, M.; Beilfuss, A.; Schmitt, J.; Hannivoort, R.A.; Kilicarslan, A.; et al. Free fatty acids repress small heterodimer partner (SHP) activation and adiponectin counteracts bile acid-induced liver injury in super obese patients with nonalcoholic steatohepatitis. Hepatology 2013, 57, 1394–1406. [Google Scholar] [CrossRef]
- You, M.; Zhou, Z.; Daniels, M.; Jogasuria, A. Endocrine adiponectin-FGF15/19 axis in ethanol-induced inflammation and alcoholic liver injury. Gene Expr. 2018, 18, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Gulfo, J.; Rotondo, F.; Ávalos de León, C.G.; Cornide-Petronio, M.E.; Fuster, C.; Gracia-Sancho, J.; Jiménez-Castro, M.B.; Peralta, C. FGF15 improves outcomes after brain dead donor liver transplantation with steatotic and non-steatotic grafts in rats. J. Hepatol. 2020, 30299–30303. [Google Scholar] [CrossRef]
- Finelli, C.; Tarantino, G. What is the role of adiponectin in obesity related non-alcoholic fatty liver disease? World J. Gastroenterol. 2013, 19, 802–812. [Google Scholar] [CrossRef]
- Gastaldelli, A.; Harrison, S.; Belfort-Aguiar, R.; Hardies, J.; Balas, B.; Schenker, S.; Cusi, K. Pioglitazone in the treatment of NASH: The role of adiponectin. Aliment. Pharmacol. Ther. 2010, 32, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Lutchman, G.; Promrat, K.; Kleiner, D.E.; Heller, T.; Ghany, M.G.; Yanovski, J.A.; Liang, T.J.; Hoofnagle, J.H. Changes in serum adipokine levels during pioglitazone treatment for nonalcoholic steatohepatitis: Relationship to histological improvement. Clin. Gastroenterol. Hepatol. 2006, 4, 1048–1052. [Google Scholar] [CrossRef]
- Landrier, J.F.; Gouranton, E.; El Yazidi, C.; Malezet, C.; Balaguer, P.; Borel, P.; Amiot, M.J. Adiponectin expression is induced by vitamin E via a peroxisome proliferator-activated receptor gamma-dependent mechanism. Endocrinology 2009, 150, 5318–5325. [Google Scholar] [CrossRef]
- Shen, Z.; Liang, X.; Rogers, C.Q.; Rideout, D.; You, M. Involvement of adiponectin-SIRT1-AMPK signaling in the protective action of rosiglitazone against alcoholic fatty liver in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G364–G374. [Google Scholar] [CrossRef]
- Hume, M.P.; Nicolucci, A.C.; Reimer, R.A. Prebiotic supplementation improves appetite control in children with overweight and obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 790–799. [Google Scholar] [CrossRef]
- Kadooka, Y.; Sato, M.; Imaizumi, K.; Ogawa, A.; Ikuyama, K.; Akai, Y.; Okano, M.; Kagoshima, M.; Tsuchida, T. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur. J. Clin. Nutr. 2010, 64, 636–643. [Google Scholar] [CrossRef]
- Shetty, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin in health and disease: Evaluation of adiponectin-targeted drug development strategies. Trends Pharmacol. Sci. 2009, 30, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Gonciarz, M.; Bielański, W.; Partyka, R.; Brzozowski, T.; Konturek, P.C.; Eszyk, J.; Celiński, K.; Reiter, R.J.; Konturek, S.J. Plasma insulin, leptin, adiponectin, resistin, ghrelin, and melatonin in nonalcoholic steatohepatitis patients treated with melatonin. J. Pineal Res. 2013, 54, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Zavos, C.; Tsiaousi, E. The role of adiponectin in the pathogenesis and treatment of non-alcoholic fatty liver disease. Diabetes Obes. Metab. 2010, 12, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Ali Khan, R.; Kapur, P.; Jain, A.; Farah, F.; Bhandari, U. Effect of orlistat on periostin, adiponectin, inflammatory markers and ultrasound grades of fatty liver in obese NAFLD patients. Ther. Clin. Risk Manag. 2017, 13, 139–149. [Google Scholar] [CrossRef]
- Galeno, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Konturek, S.J.; Konturek, P.C.; Brzozowski, T.; Bubenik, G.A. Role of melatonin in upper gastrointenstinal tract. J. Physiol. Pharmacol. 2007, 58, 23–52. [Google Scholar]
- Polyzos, S.A.; Kountouras, J.; Zavos, C. Adiponectin as a potential therapeutic agent for nonalcoholic steatohepatitis. Hepatol. Res. 2010, 40, 446–447. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Zavos, C. Adiponectin in non-alcoholic fatty liver disease treatment: Therapeutic perspectives and unresolved dilemmas. Int J. Clin. Pract. 2011, 65, 373–374. [Google Scholar] [CrossRef]
- Narasimhan, M.L.; Coca, M.A.; Jin, J.; Yamauchi, T.; Ito, Y.; Kadowaki, T.; Kim, K.K.; Pardo, J.M.; Damsz, B.; Hasegawa, P.M.; et al. Osmotin is a homolog of mammalian adiponectin and controls apoptosis in yeast through a homolog of mammalian adiponectin receptor. Mol. Cell 2005, 17, 171–180. [Google Scholar] [CrossRef]
- Razavizade, M.; Jamali, R.; Arj, A.; Matini, S.M.; Moraveji, A.; Taherkhani, E. The effect of pioglitazone and metformin on liver function tests, insulin resistance, and liver fat content in nonalcoholic Fatty liver disease: A randomized double blinded clinical trial. Hepat. Mon. 2013, 13, e9270. [Google Scholar] [CrossRef]
- Promrat, K.; Lutchman, G.; Uwaifo, G.I.; Freedman, R.J.; Soza, A.; Heller, T.; Doo, E.; Ghany, M.; Premkumar, A.; Park, Y.; et al. A pilot study of pioglitazone treatment for nonalcoholic steatohepatitis. Hepatology 2004, 39, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Mofrad, P.S.; Contos, M.J.; Sargeant, C.; Luketic, V.A.; Sterling, R.K.; Stravitz, R.T.; Shiffman, M.L.; Clore, J.; Mills, A.S. A pilot study of vitamin E versus vitamin E and pioglitazone for the treatment of nonalcoholic steatohepatitis. Clin. Gastroenterol. Hepatol. 2004, 2, 1107–1115. [Google Scholar] [CrossRef]
- Lutchman, G.; Modi, A.; Kleiner, D.E.; Promrat, K.; Heller, T.; Ghany, M.; Borg, B.; Loomba, R.; Liang, T.J.; Premkumar, A.; et al. The effects of discontinuing pioglitazone in patients with nonalcoholic steatohepatitis. Hepatology 2007, 46, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Belfort, R.; Harrison, S.A.; Brown, K.; Darland, C.; Finch, J.; Hardies, J.; Balas, B.; Gastaldelli, A.; Tio, F.; Pulcini, J.; et al. A placebo-controlled trial of pioglitazone in subjects with nonalcoholic steatohepatitis. N. Engl. J. Med. 2006, 355, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Liss, K.H.; Finck, B.N. PPARs and nonalcoholic fatty liver disease. Biochimie 2017, 136, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Matsusue, K.; Haluzik, M.; Lambert, G.; Yim, S.H.; Gavrilova, O.; Ward, J.M.; Brewer, B.; Reitman, M.L.; Gonzalez, F.J. Liver-specific disruption of PPARγ in leptin-deficient mice improves fatty liver but aggravates diabetic phenotypes. J. Clin. Invest. 2003, 111, 737–747. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, I.; Rodríguez-Juan, C.; Díaz-Sanjuán, T.; Martínez, M.Á.; Muñoz-Yagüe, T.; Solís-Herruzo, J.A. Effects of rosiglitazone on the liver histology and mitochondrial function in ob/ob mice. Hepatology 2007, 46, 414–423. [Google Scholar] [CrossRef]
- Hadi, H.E.; Vettor, R.; Rossato, M. Vitamin E as a treatment for nonalcoholic fatty liver disease: Reality or myth? Antioxidants 2018, 7, 12. [Google Scholar] [CrossRef]
- Schürks, M.; Glynn, R.J.; Rist, P.M.; Tzourio, C.; Kurth, T. Effects of vitamin E on stroke subtypes: Meta-analysis of randomised controlled trials. BMJ 2010, 341, c5702. [Google Scholar] [CrossRef]
- Klein, E.A.; Thompson, I.M.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the risk of prostate cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef]
- Hemmeryckx, B.; Gaekens, M.; Gallacher, D.J.; Lu, H.R.; Lijnen, H.R. Effect of rosiglitazone on liver structure and function in genetically diabetic Akita mice. Basic Clin. Pharmacol. Toxicol. 2013, 113, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Blandon, J.; Rude, J.; Elfar, A.; Mukherjee, D. PPAR-γ agonist in treatment of diabetes: Cardiovascular safety considerations. Cardiovasc. Hematol. Agents Med. Chem. 2012, 10, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Douglas, I.J.; Langham, J.; Bhaskaran, K.; Brauer, R.; Smeeth, L. Orlistat and the risk of acute liver injury: Self controlled case series study in UK clinical practice research datalink. BMJ 2013, 346, f1936. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, E.; Gemici, G.; Bayrak, F.; Kalkan, A.K.; Degertekin, M. Acute myocardial infarction in a 24 year-old man possibly associated with sibutramine use. Int. J. Cardiol. 2009, 137, e43–e45. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Tapia, N.C.; Tellez-Avila, F.I.; Bedogni, G.; Crocè, L.S.; Masutti, F.; Tiribelli, C. Systematic review and meta-analysis on the adverse events of rimonabant treatment: Considerations for its potential use in hepatology. BMC Gastroenterol. 2009, 9, 75. [Google Scholar] [CrossRef]
- Halberg, N.; Schraw, T.D.; Wang, Z.V.; Kim, J.Y.; Yi, J.; Hamilton, M.P.; Luby-Phelps, K.; Scherer, P.E. Systemic fate of the adipocyte-derived factor adiponectin. Diabetes 2009, 58, 1961–1970. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K. Adiponectin and adiponectin receptors in obesity-linked insulin resistance. Novartis Found. Symp. 2007, 286, 164–176. [Google Scholar] [CrossRef]
- Neumeier, M.; Hellerbrand, C.; Gäbele, E.; Buettner, R.; Bollheimer, C.; Weigert, J.; Schäffler, A.; Weiss, T.S.; Lichtenauer, M.; Schölmerich, J.; et al. Adiponectin and its receptors in rodent models of fatty liver disease and liver cirrhosis. World J. Gastroenterol. 2006, 12, 5490–5494. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid Res. 2009, 48, 1–26. [Google Scholar] [CrossRef]
- Gatselis, N.K.; Ntaios, G.; Makaritsis, K.; Dalekos, G.N. Adiponectin: A key playmaker adipocytokine in non-alcoholic fatty liver disease. Clin. Exp. Med. 2014, 14, 121–131. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Toulis, K.A.; Goulis, D.G.; Zavos, C.; Kountouras, J. Serum total adiponectin in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Metabolism 2011, 60, 313–326. [Google Scholar] [CrossRef]
- Lian, K.; Feng, Y.N.; Li, R.; Liu, H.L.; Han, P.; Zhou, L.; Li, C.X.; Wang, Q. Middle- and high-molecular weight adiponectin levels in relation to nonalcoholic fatty liver disease. J. Clin. Lab. Anal. 2020, 34, e23148. [Google Scholar] [CrossRef] [PubMed]
- Ozcelik, F.; Yuksel, C.; Arslan, E.; Genc, S.; Omer, B.; Serdar, M.A. Relationship between visceral adipose tissue and adiponectin, inflammatory markers and thyroid hormones in obese males with hepatosteatosis and insulin resistance. Arch. Med. Res. 2013, 44, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Balmer, M.L.; Joneli, J.; Schoepfer, A.; Stickel, F.; Thormann, W.; Dufour, J.F. Significance of serum adiponectin levels in patients with chronic liver disease. Clin. Sci. 2010, 119, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Jamali, R.; Arj, A.; Razavizade, M.; Aarabi, M.H. Prediction of nonalcoholic fatty liver disease via a novel panel of serum adipokines. Medicine 2016, 95, e2630. [Google Scholar] [CrossRef]
- Targher, G.; Bertolini, L.; Scala, L.; Poli, F.; Zenari, L.; Falezza, G. Decreased plasma adiponectin concentrations are closely associated with nonalcoholic hepatic steatosis in obese individuals. Clin. Endocrinol. 2004, 61, 700–703. [Google Scholar] [CrossRef]
- Wong, V.W.; Wong, G.L.; Choi, P.C.; Chan, A.W.; Li, M.K.; Chan, H.Y.; Chim, A.M.; Yu, J.; Sung, J.J.; Chan, H.L. Disease progression of non-alcoholic fatty liver disease: A prospective study with paired liver biopsies at 3 years. Gut 2010, 59, 969–974. [Google Scholar] [CrossRef]
- Zelber-Sagi, S.; Lotan, R.; Shlomai, A.; Webb, M.; Harrari, G.; Buch, A.; Nitzan Kaluski, D.; Halpern, Z.; Oren, R. Predictors for incidence and remission of NAFLD in the general population during a seven-year prospective follow-up. J. Hepatol. 2012, 56, 1145–1151. [Google Scholar] [CrossRef]
- Verhaegh, P.; Bavalia, R.; Winkens, B.; Masclee, A.; Jonkers, D.; Koek, G. Noninvasive tests do not accurately differentiate nonalcoholic steatohepatitis from simple steatosis: A systematic review and meta-analysis. Clin. Gastroenterol. Hepatol. 2018, 16, 837–861. [Google Scholar] [CrossRef]
- Aksoy, G.K.; Artan, R.; Aksoy, C.; Özdem, S.; Atalay, A.; Yılmaz, A. Role of soluble adiponectin receptor 2 in non-alcoholic fatty liver disease in children. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 470–478. [Google Scholar] [CrossRef]
- Boyraz, M.; Cekmez, F.; Karaoglu, A.; Cinaz, P.; Durak, M.; Bideci, A. Serum adiponectin, leptin, resistin and RBP4 levels in obese and metabolic syndrome children with nonalcoholic fatty liver disease. Biomark. Med. 2013, 7, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Kawahara, H.; Ozaki, K.; Fukura, M.; Yano, H.; Tsuchishima, M.; Tsutsumi, M.; Takase, S. Usefulness of a combined evaluation of the serum adiponectin level, HOMA-IR, and serum type IV collagen 7S level to predict the early stage of nonalcoholic steatohepatitis. Am. J. Gastroenterol. 2007, 102, 1931–1938. [Google Scholar] [CrossRef]
- Da Silva, T.E.; Costa-Silva, M.; Correa, C.G.; Denardin, G.; Ayres Alencar, M.L.; Pacheco Honorico Coelho, M.S.; Muraro-Wildner, L.; Luiza-Bazzo, M.; González-Chica, D.A.; Dantas-Correa, E.B.; et al. Clinical significance of serum adiponectin and resistin levels in liver cirrhosis. Ann. Hepatol. 2018, 17, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Salman, T.A.; Allam, N.; Azab, G.I.; Shaarawy, A.A.; Hassouna, M.M.; El-Haddad, O.M. Study of adiponectin in chronic liver disease and cholestasis. Hepatol. Int. 2010, 4, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Siegel, A.B.; Goyal, A.; Salomao, M.; Wang, S.; Lee, V.; Hsu, C.; Rodriguez, R.; Hershman, D.L.; Brown, R.S.; Neugut, A.I.; et al. Serum adiponectin is associated with worsened overall survival in a prospective cohort of hepatocellular carcinoma patients. Oncology 2015, 88, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Kalafateli, M.; Triantos, C.; Tsochatzis, E.; Michalaki, M.; Koutroumpakis, E.; Thomopoulos, K.; Kyriazopoulou, V.; Jelastopulu, E.; Burroughs, A.; Lambropoulou-Karatza, C.; et al. Adipokines levels are associated with the severity of liver disease in patients with alcoholic cirrhosis. World J. Gastroenterol. 2015, 21, 3020–3029. [Google Scholar] [CrossRef]
- Nkontchou, G.; Bastard, J.P.; Ziol, M.; Aout, M.; Cosson, E.; Ganne-Carrie, N.; Grando-Lemaire, V.; Roulot, D.; Capeau, J.; Trinchet, J.C.; et al. Insulin resistance, serum leptin, and adiponectin levels and outcomes of viral hepatitis C cirrhosis. J. Hepatol. 2010, 53, 827–833. [Google Scholar] [CrossRef]
- Hamdy, K.; Al Swaff, R.; Hussein, H.A.; Gamal, M. Assessment of serum adiponectin in Egyptian patients with HCV-related cirrhosis and hepatocellular carcinoma. J. Endocrinol. Invest. 2015, 38, 1225–1231. [Google Scholar] [CrossRef]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [Google Scholar] [CrossRef]
- Wu, Z.J.; Cheng, Y.J.; Gu, W.J.; Aung, L.H. Adiponectin is associated with increased mortality in patients with already established cardiovascular disease: A systematic review and meta-analysis. Metabolism 2014, 63, 1157–1166. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Isoform and Receptor | Type of Liver and Specie | Surgical Condition | Effect and Signaling Pathways |
|---|---|---|---|---|
| Adiponectin recombinant (Rat Acrp30) [16] | Not reported | Steatotic and non-steatotic livers from Zucker rats | LT Ischemia: 6 h Reperfusion: 4 h | ↓ Hepatic injury and mortality ↑ Ki-67 Adiponectin-↑ Resistin-↑ PI3K/Akt |
| Dietary model [17] | Not reported; AdipoR1 and AdipoR2 | Steatotic livers from Sprague-Dawley MCD or HFD rat | Partial (70%) hepatectomy | ↓ AdipoR1 and AdipoR2 ↑ Hepatic adiponectin, TNF-α, AMPK AdipoR1-↑ AMPK in MCD and HFD AdipoR2-↓ PPARα in MCD |
| Adiponectin (-/-) [31] | Not reported | Livers from B6.129-Adipoqtm1Cha knockout mice | Partial hepatectomy | ↓ Regeneration by controlling cell cycle progression, cytokine signaling and growth factor bioavailability Adiponectin-↑ STAT3 |
| Adiponectin recombinant [60] | Not reported | Livers from Wistar rats | Partial warm ischemia Ischemia: 60 min Reperfusion: 6, 24 h | ↓ Hepatic injury, inflammatory cell infiltration, IL-1β, IL-6, TNF-α, CCL2, CXCL10, ICAM1, apoptosis Adiponectin-↑ AMPK/eNOS |
| Adiponectin recombinant and supplementation (Rat gAcrp30) [61] | Not reported | Livers from Wistar rats | Chronic intermittent hypoxia events for 8 h per day for 4 months | ↓ Hepatic injury, ROS production, fasting blood glucose, triglycerides Adiponectin-↑ AMPK |
| Adiponectin recombinant (Rat gAcrp30) [62] | Not reported | Livers from Sprague-Dawley rats | LT Ischemia: 30 min Reperfusion: 3, 6, 12, 24 h | ↓ Bile duct injury and apoptosis, Fas, caspase 3, TNF-α, NF-κB activation, MPO, IL-6 Adiponectin-↓ NF-κB |
| Treatment | Isoform and Receptor | Type of Liver and Specie | Surgical Condition | Effect and Signaling Pathways |
|---|---|---|---|---|
| Adiponectin recombinant (Human gAcrp30) [66] | HMW and Adipo R1 and AdipoR2 | KC and RAW 264.7 macrophages from Wistar rats with chronic ethanol-feeding | Cell culture | ↓ TLR4 (/MyD88), IFN-β, CXCL10 Adiponectin–↓ TLR4 |
| CHIP (-/-) [68] | Not reported; AdipoR1 and AdipoR2 | Livers from CHIP knockout mice | Cell culture | ↓ Oxidative stress and JNK ↑ Adiponectin, AdipoR1, AdipoR2, AMPK and FOXO Adiponectin-↑ AMPK-FOXO |
| mLipin-1 (-/-) (Human gAcrp30) [69] | HMW; AdipoR1 and AdipoR2 | Livers from mLipin-1 knockout mice with chronic ethanol-feeding | NA | ↓ Hepatic injury, inflammation, NF-κB, ↑ Adiponectin, AdipoR1, AdipoR2 and FGF15 Adiponectin-↑ FGF15 signaling |
| mNT (-/-) [70] | Not reported | mNT knockout mice with chronic ethanol-feeding | NA | ↓ Hepatic injury, NF-κB, oxidative stress ↑ Adiponectin, FGF15, Sirt1 Adiponectin-↑ FGF15 signaling |
| Pioglitazone [75] | Not reported | NASH patients | NA | ↓ Hepatic steatosis and necroinflammation ↑ Adiponectin Adiponectin-↓ NF-κB/JNK |
| Rosiglitazone [78] | HMW; AdipoR1 and AdipoR2 | Livers from C57BL/6J mice with ethanol-feeding | NA | ↓ Hepatic injury, steatosis, lipogenesis ↑ Adiponectin and hepatic AdipoR1/R2 Adiponectin–↑ SIRT1-AMPK |
| Prebiotic fiber supplementation [79] | Not reported | Children patients with overweight and obese | NA | ↑ Adiponectin and ghrelin Not reported adiponectin signaling pathway |
| Probiotic Lactobacillus gasseri (SBT2055) [80] | HMW, MMW, LMW and not reported receptor | Obese patients | NA | ↓ Abdominal visceral fat ↑ HMW in obese and control group ↑ MMW only in control group Not reported adiponectin signaling pathway |
| Melatonin [82] | Not reported | NASH patients | NA | ↓ HOMA-IR ↑ Adiponectin, leptin and ghrelin Not reported adiponectin signaling pathway |
| Orlistat [84] | Not reported | NAFLD patients | NA | ↓ Fatty infiltration, periostin, TNF-α ↑ Adiponectin Not reported adiponectin signaling pathway |
| Disease | Subjects (Etiology) | Adiponectin Levels | Effect |
|---|---|---|---|
| Alcoholic liver disease (ALD) [58] | 147 patients | 18.69 ALD 6.38 control | ↑ Adiponectin (Acrp30) associated with advanced liver dysfunction and ALD complications |
| NAFLD [112] | 63 patients | 4.26 ± 2.71 µg/mL NAFLD 5.85 ± 3.74 µg/mL controls | ↓ Three isoforms of adiponectin. HMW and MMW adiponectin involved in the pathogenesis and progression of NAFLD |
| NAFLD [113] | 315 patients (129 mild, 145 moderate, 41 severe) | 13.6 ± 3.3 µg/mL mild 12.4 ± 3.7 µg/mL moderate 11.6 ± 3.5 µg/mL severe | ↑ adiponectin correlated with a decreased risk of developing type 2 diabetes |
| NAFLD [114] | 232 patients (45 cirrhosis, 71 viral hepatitis, 64 NAFLD, 52 others) | 18.6 ± 14.5 µg/mL cirrhosis 8.4 ± 6.1 µg/mL without cirrhosis 4.8 ± 3.5 µg/mL NAFLD 9.1 µg/mL controls | Adiponectin correlate positively with markers of hepatic fibrosis. ↓ Adiponectin in NAFLD and ↑ in cirrhosis |
| NAFLD [115] | 70 patients | 8.14 ± 2.91 mg/L NAFLD 13.63 ± 2.88 mg/L controls | ↓ Adiponectin and ↑ visfatin, IL-6, TNF-α associated with increased NAFLD |
| NAFLD [117] | 52 patients | 3.9 (2.5 – 6.2) µg/mL | Adiponectin, TNF-α, IL-6, leptin were not associated with disease progression |
| NAFLD [118] | 147 patients | 9.6 ± 4.1 µg/mL NAFLD 14.0 ± 10.1 µg/mL without NAFLD | ↓ Adiponectin associated with NAFLD |
| NAFLD [120] | 71 children patients (37 with NAFLD, 14 with NASH, 20 without NAFLD) | 13.15 ± 5.33 ng/mL NAFLD 12.64 ± 5.54 ng/mL NASH | Adiponectin levels were similar in patients with and without NAFLD. ↑ AdipoR2 in NAFLD and is a noninvasive marker for diagnosis |
| NAFLD [121] | 148 children patients (63 steatosis, 12 steatosis and ↑ ALT, 85 without steatosis) | 2.7 ± 0.7 µg/mL steatosis 2.5 ± 0.4 µg/mL NAFLD 4.7 ± 1.1 µg/mL without steatosis | ↓ Adiponectin were negatively correlated with ALT activity |
| Cirrhosis [123] | 122 patients | 21.59 µg/mL cirrhosis 12.52 µg/mL controls | ↑ Adiponectin associated with ↑ liver dysfunction and worse prognosis |
| Cirrhosis [124] | 70 patients (40 cirrhosis, 30 cirrhosis and cholestasis) | 15.1 ± 12.1 µg/mL cirrhosis 21.28 ± 10.2 µg/mL cirrhosis with cholestasis 4.7 ± 4.48 µg/mL controls | ↑ Adiponectin shows correlation with degree of hepatocellular injury and cholestasis; but not with parameters of body composition or metabolism |
| Cirrhosis [125] | 140 patients | 13.050 ng/mL | Adiponectin was an independent predictor of overall survival in HCC patients |
| Cirrhosis [126] | 40 patients with non-diabetic alcoholic cirrhosis | 10.23 µg/mL | ↑ Adiponectin associated with shorter survival in the univariate analysis but not in the multivariate analysis |
| Cirrhosis [127] | 248 patients with compensated viral hepatitis C cirrhosis | 16.5 ± 15.3 µg/mL cirrhosis | Adiponectin was not related to HCC, liver-related death or LT during follow-up |
| Cirrhosis [128] | 90 patients with hepatitis C-related liver cirrhosis (61 with, 29 without) | 5.213 ± 3.840 µg/mL cirrhosis with HCC 9.000 ± 2.234 µg/mL cirrhosis without HCC | ↓ Adiponectin levels associated with HCC; a biomarker of HCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heydari, M.; Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Peralta, C. Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases? Int. J. Mol. Sci. 2020, 21, 5242. https://doi.org/10.3390/ijms21155242
Heydari M, Cornide-Petronio ME, Jiménez-Castro MB, Peralta C. Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases? International Journal of Molecular Sciences. 2020; 21(15):5242. https://doi.org/10.3390/ijms21155242
Chicago/Turabian StyleHeydari, Misaq, María Eugenia Cornide-Petronio, Mónica B. Jiménez-Castro, and Carmen Peralta. 2020. "Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases?" International Journal of Molecular Sciences 21, no. 15: 5242. https://doi.org/10.3390/ijms21155242
APA StyleHeydari, M., Cornide-Petronio, M. E., Jiménez-Castro, M. B., & Peralta, C. (2020). Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases? International Journal of Molecular Sciences, 21(15), 5242. https://doi.org/10.3390/ijms21155242