Proteome Analysis Reveals Syndecan 1 Regulates Porcine Sapelovirus Replication


Abstract
1. Introduction
2. Results
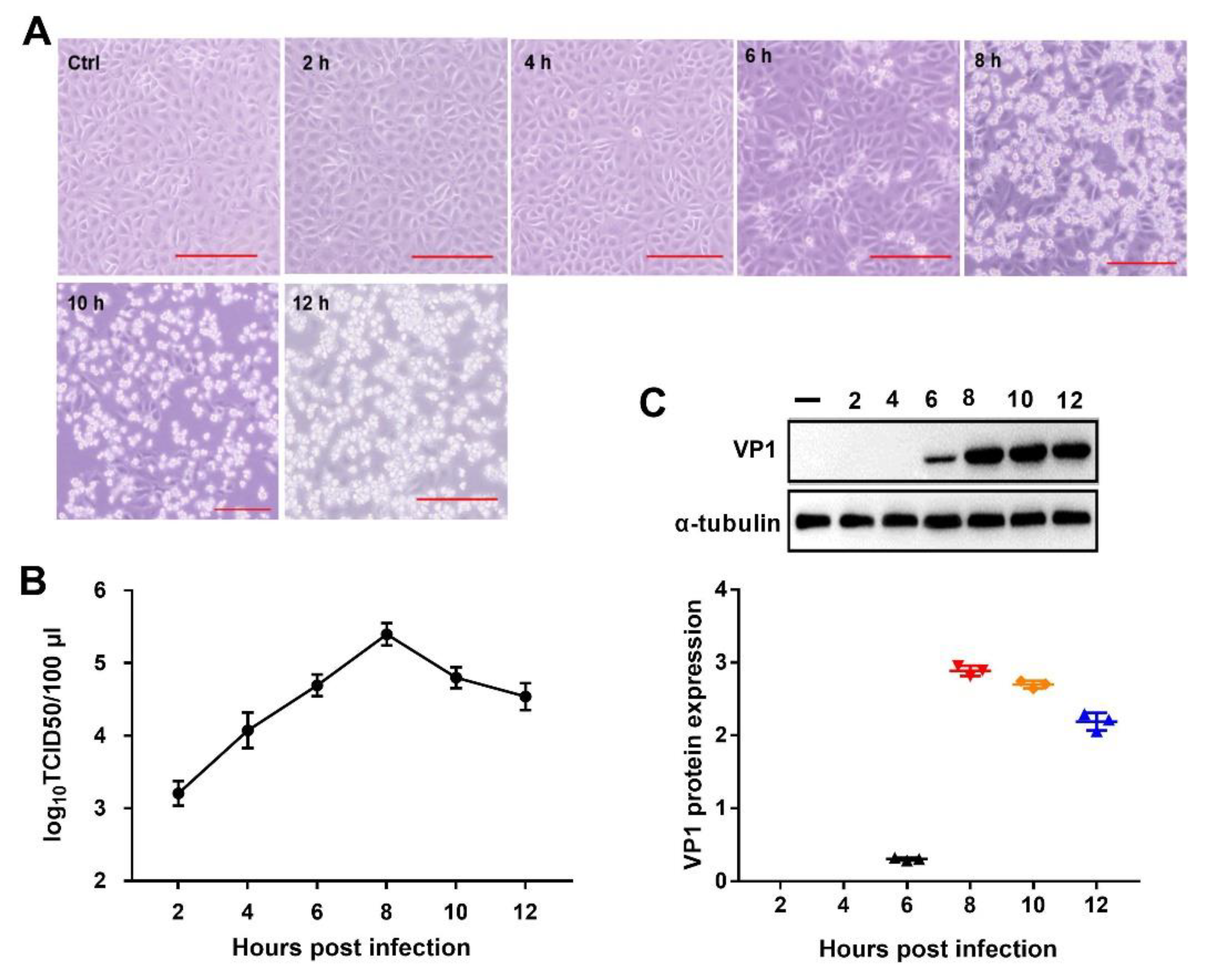
2.1. PSV Infection in PK-15 Cells
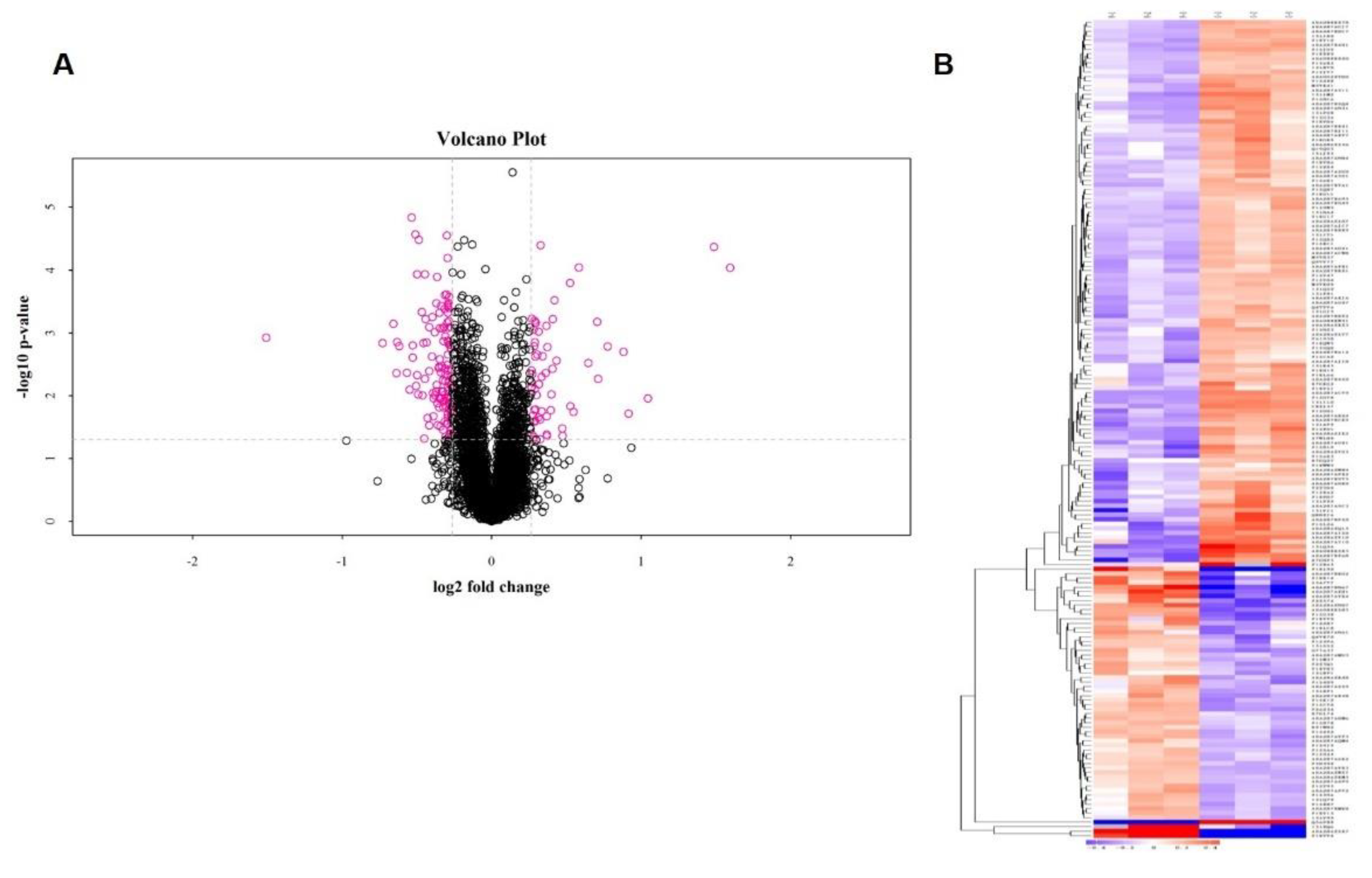
2.2. Identification of Differentially Expressed Proteins (DEPs)
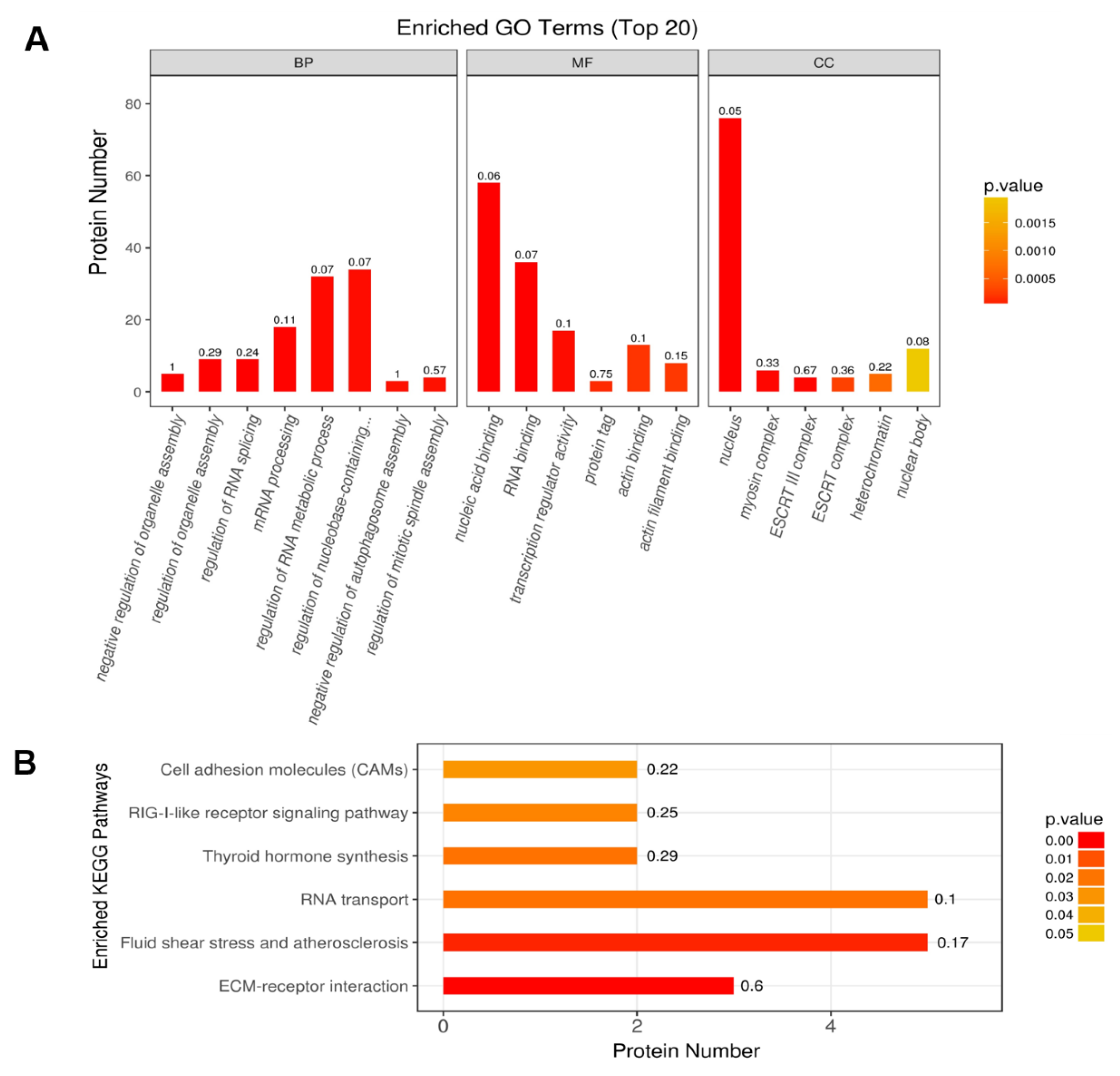
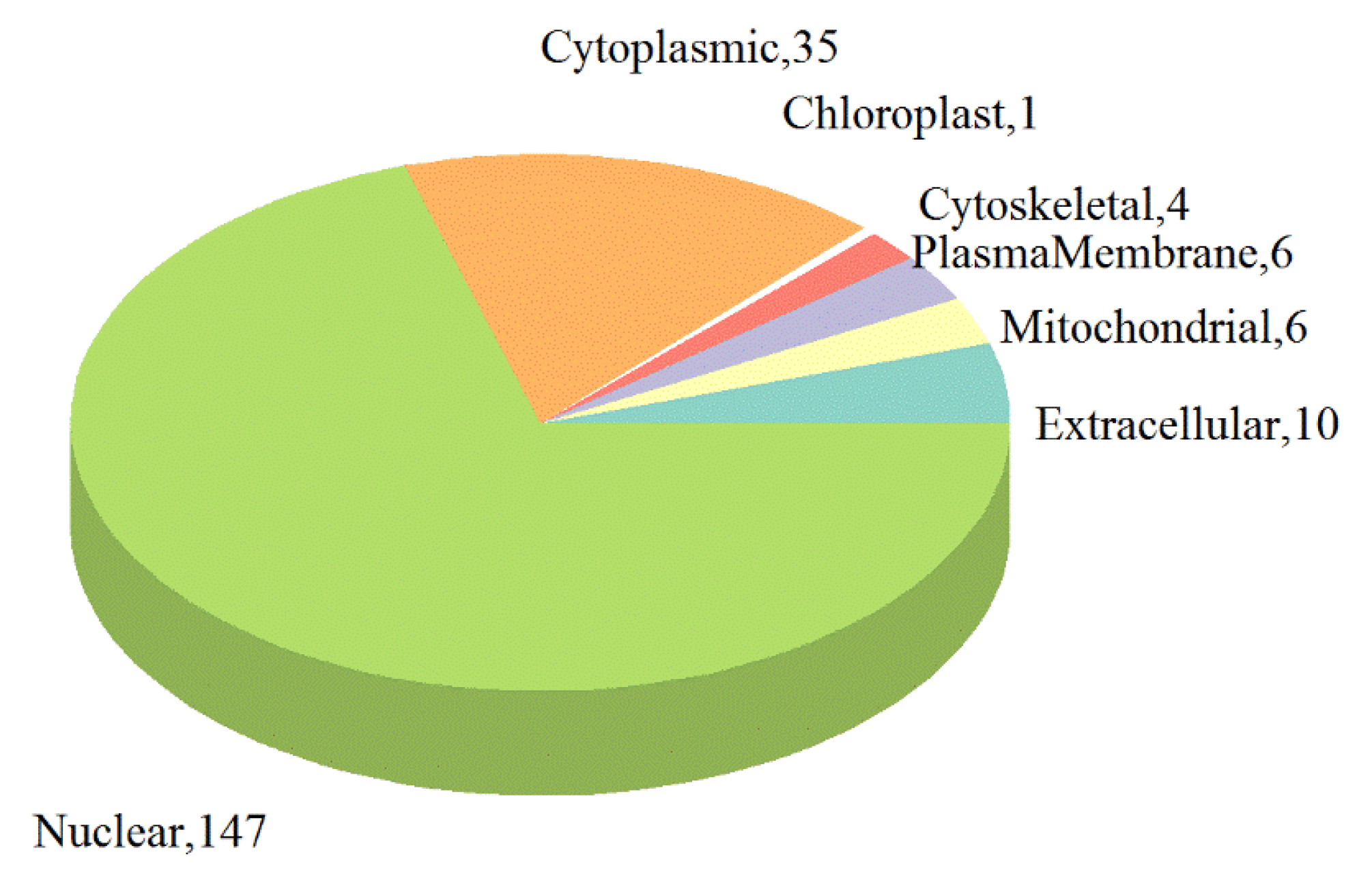
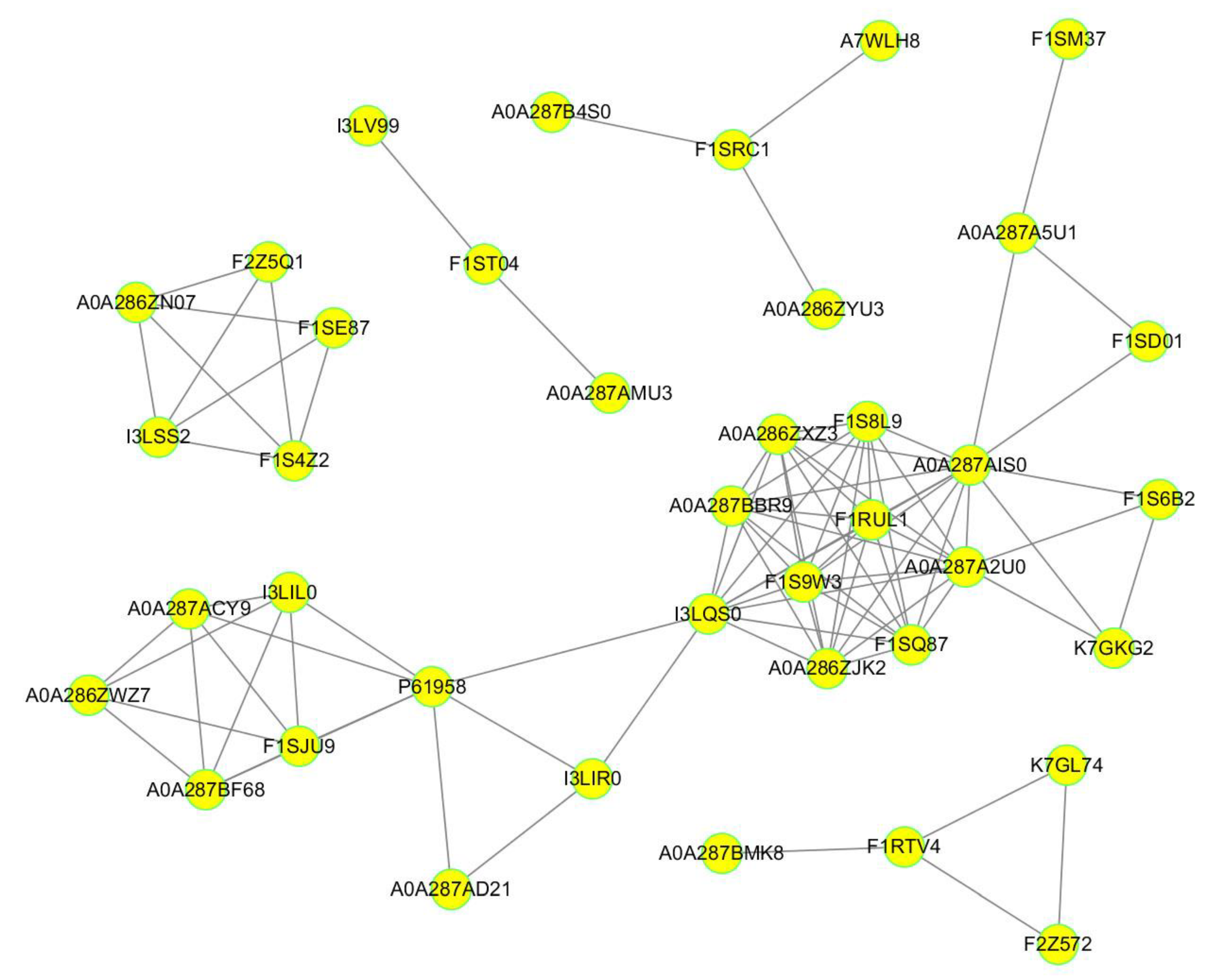
2.3. Functional Classification and Enrichment Analysis of the Identified DEPs
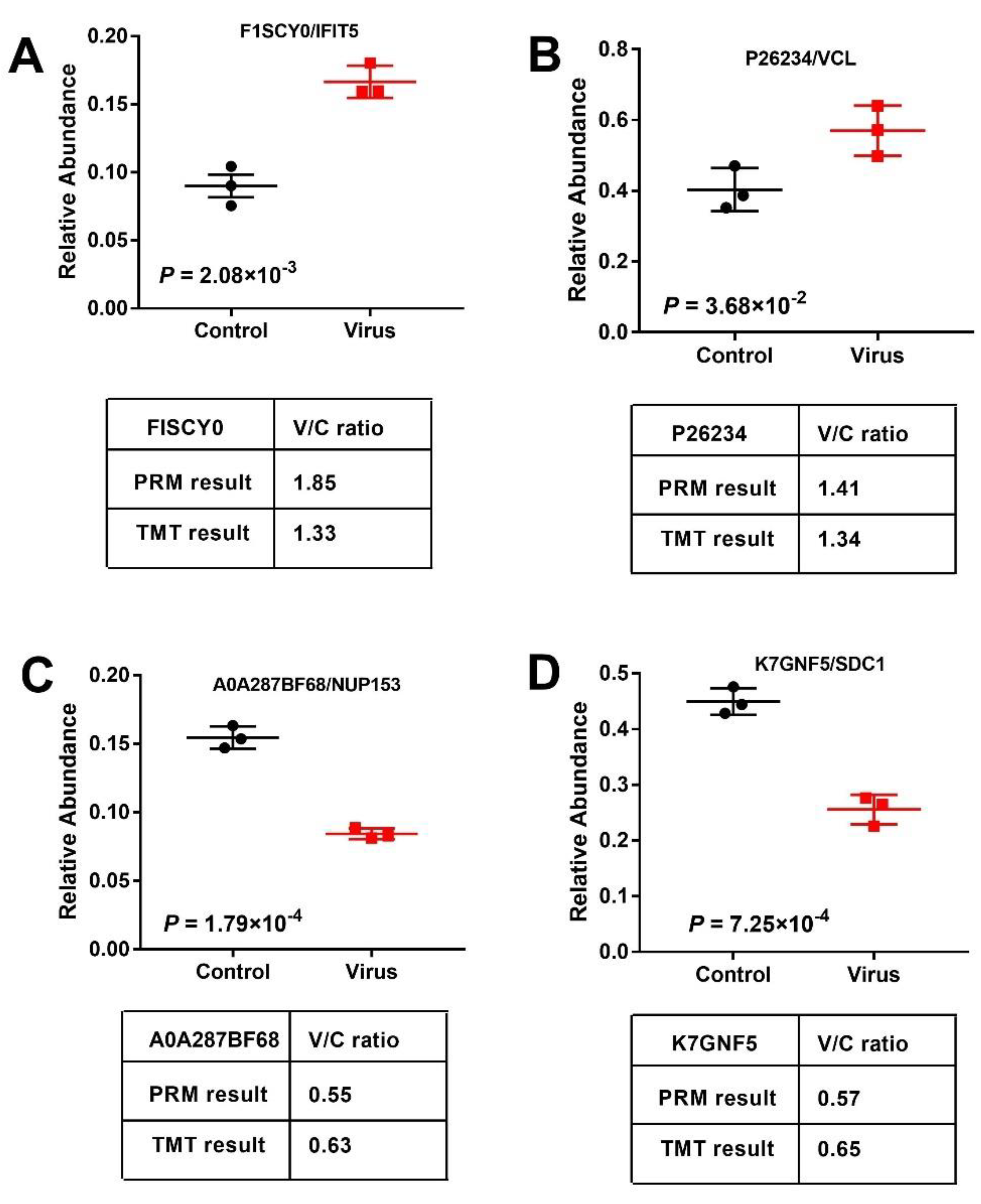
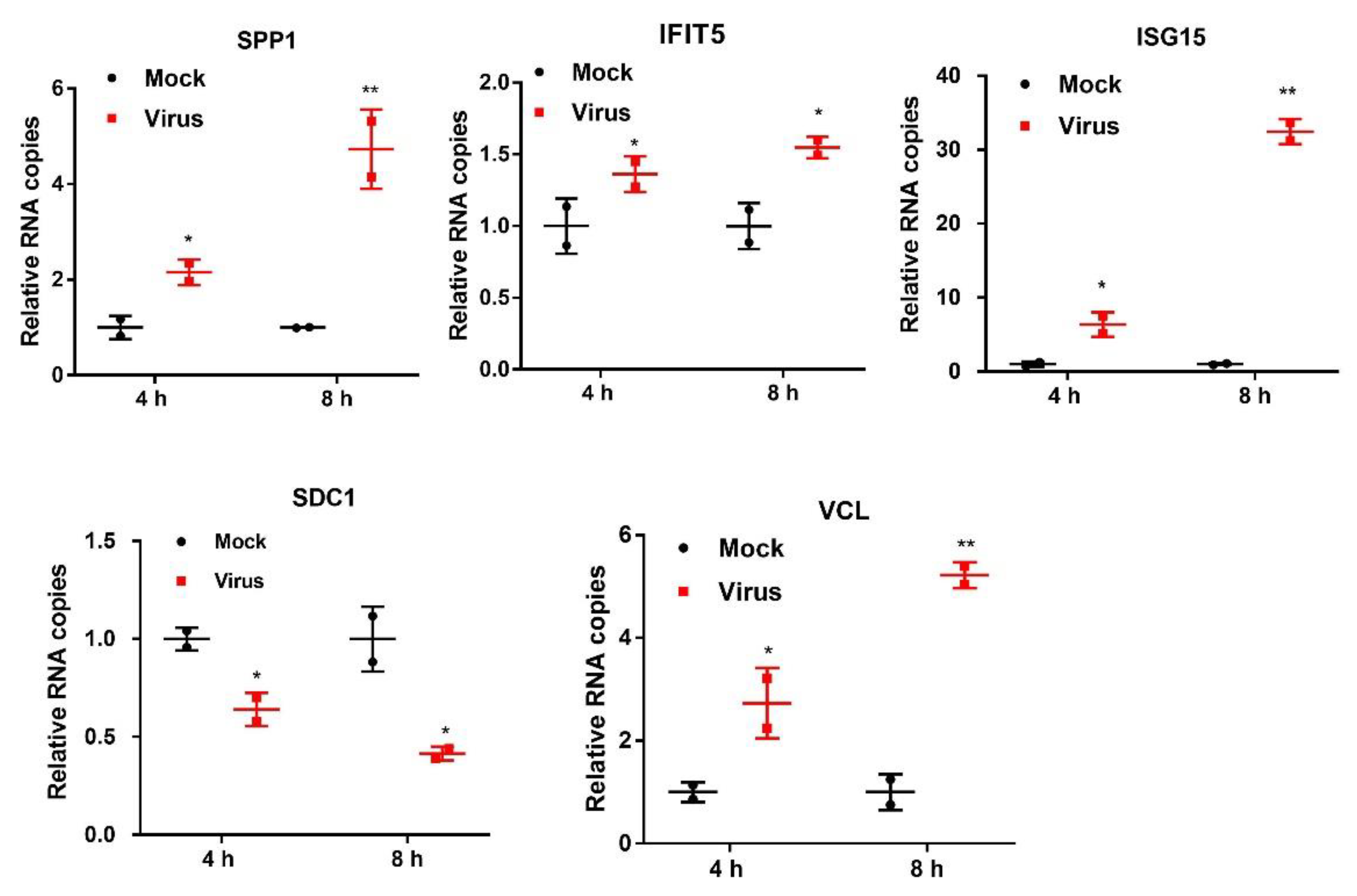
2.4. Validation of DEPs by PRM and qRT-PCR
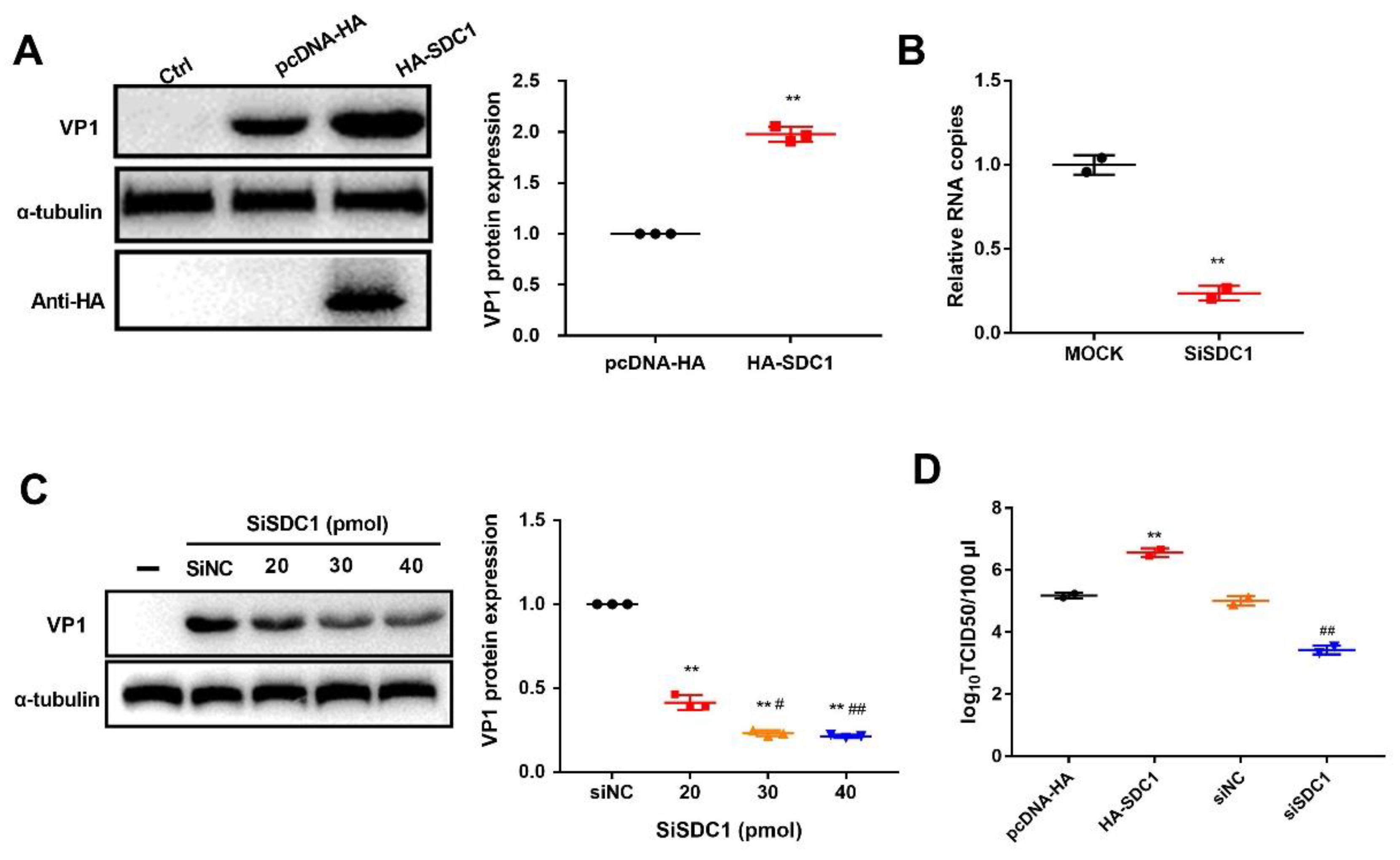
2.5. SDC1 Promotes PSV Replication
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Virus
4.2. Virus Titration
4.3. Protein Extraction
4.4. Tandem Mass Tag (TMT) Labeling
4.5. High pH Reversed-Phase Fractionation
4.6. Liquid Chromatography–Mass Spectrometry (LC–MS/MS) Analysis
4.7. Protein Identification and Quantitation
4.8. Bioinformatics Analyses
4.9. PRM Analysis
4.10. The qRT-PCR
4.11. Plasmid and RNA Interference Transfection
4.12. Western Blotting
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PSV | Porcine sapelovirus A |
| MS | Mass spectrometry |
| TMT | Tandem mass tag |
| PRM | Parallel reaction monitoring |
| SDC1 | Syndecan-1 |
| DMEM | Dulbecco’s minimal essential medium |
| MOI | Multiplicity of infection |
| hpi | Hour post-infection |
| GO | Gene ontology |
| BP | Biological process |
| MF | Molecular function |
| CC | Cellular component |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| siRNAs | Small interfering RNAs |
| NC | Negative control |
| CPEs | Cytopathogenic effects |
| DEPs | Differentially expressed proteins |
| SPP1 | Osteopontin |
| IFIT5 | Induced protein with tetratricopeptide repeats 5 |
| ISG15 | ISG15 ubiquitin-like modifier |
| VCL | Vinculin |
References
- Stanway, G. Structure, function and evolution of picornaviruses. J. Gen. Virol. 1990, 71, 2483–2501. [Google Scholar] [CrossRef] [PubMed]
- Arruda, P.; Arruda, B.; Schwartz, K.; Vannucci, F.; Resende, T.; Rovira, A.; Sundberg, P.; Nietfeld, J.; Hause, B. Detection of a novel sapelovirus in central nervous tissue of pigs with polioencephalomyelitis in the USA. Transbound. Emerg. Dis. 2017, 64, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, F.; Zhou, Q.; Li, W.; Song, Y.; Pan, Y.; Zhang, X.; Xue, C.; Bi, Y.; Cao, Y. Complete genome sequence of a novel porcine Sapelovirus strain YC2011 isolated from piglets with diarrhea. J. Virol. 2012, 86, 10898. [Google Scholar] [CrossRef][Green Version]
- Lan, D.; Ji, W.; Yang, S.; Cui, L.; Yang, Z.; Yuan, C.; Hua, X. Isolation and characterization of the first Chinese porcine sapelovirus strain. Arch. Virol. 2011, 156, 1567–1574. [Google Scholar] [CrossRef]
- Kim, D.-S.; Son, K.-Y.; Koo, K.-M.; Kim, J.-Y.; Alfajaro, M.M.; Park, J.-G.; Hosmillo, M.; Soliman, M.; Baek, Y.-B.; Cho, E.-H. Porcine sapelovirus uses α2, 3-linked sialic acid on GD1a ganglioside as a receptor. J. Virol. 2016, 90, 4067–4077. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Liu, J.; Fang, L.; Kataoka, M.; Takeda, N.; Wakita, T.; Li, T.C. Characterization of porcine sapelovirus isolated from Japanese swine with PLC/PRF/5 cells. Transbound. Emerg. Dis. 2018, 65, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Cui, L.; Yu, X.; Zhang, Z.; Shen, X.; Hua, X. Entry of sapelovirus into IPEC-J2 cells is dependent on caveolae-mediated endocytosis. Virol. J. 2019, 16, 019–1144. [Google Scholar] [CrossRef] [PubMed]
- An, K.; Fang, L.; Luo, R.; Wang, D.; Xie, L.; Yang, J.; Chen, H.; Xiao, S. Quantitative proteomic analysis reveals that transmissible gastroenteritis virus activates the JAK-STAT1 signaling pathway. J. Proteome Res. 2014, 13, 5376–5390. [Google Scholar] [CrossRef]
- Xin, Q.-L.; Deng, C.-L.; Chen, X.; Wang, J.; Wang, S.-B.; Wang, W.; Deng, F.; Zhang, B.; Xiao, G.; Zhang, L.-K. Quantitative proteomic analysis of mosquito C6/36 cells reveals host proteins involved in Zika virus infection. J. Virol. 2017, 91, e00554-17. [Google Scholar] [CrossRef]
- Lan, J.; Zhang, R.; Yu, H.; Wang, J.; Xue, W.; Chen, J.; Lin, S.; Wang, Y.; Xie, Z.; Jiang, S. Quantitative Proteomic Analysis Uncovers the Mediation of Endoplasmic Reticulum Stress-Induced Autophagy in DHAV-1-Infected DEF Cells. Int. J. Mol. Sci. 2019, 20, 6160. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, J.; Zhu, L.; Toufeeq, S.; Xu, X.; You, L.; Li, B.; Hu, P.; Xu, J. Quantitative label-free proteomic analysis reveals differentially expressed proteins in the digestive juice of resistant versus susceptible silkworm strains and their predicted impacts on BmNPV infection. J. Proteom. 2020, 210, 103527. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Li, J.; Mao, L.; Liu, M.; Zhu, X.; Li, W.; Sun, M.; Ji, X.; Xiao, F.; Yang, L.; et al. Proteomics analysis reveals heat shock proteins involved in caprine parainfluenza virus type 3 infection. BMC Vet. Res. 2019, 15, 019–1897. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, B.; Downard, K.M. A proteomics approach to survey the antigenicity of the influenza virus by mass spectrometry. Proteomics 2010, 6, 2034–2041. [Google Scholar] [CrossRef]
- Miao, M.; Yu, F.; Wang, D.; Tong, Y.; Yang, L.; Xu, J.; Qiu, Y.; Zhou, X.; Zhao, X. Proteomics Profiling of Host Cell Response via Protein Expression and Phosphorylation upon Dengue Virus Infection. Virol. Sin. 2019, 34, 549–562. [Google Scholar] [CrossRef]
- Hao, J.; Guo, H.; Shi, X.; Wang, Y.; Wan, Q.; Song, Y.-B.; Zhang, L.; Dong, M.; Shen, C. Comparative proteomic analyses of two Taxus species (Taxus× media and Taxus mairei) reveals variations in the metabolisms associated with paclitaxel and other metabolites. Plant. Cell Physiol. 2017, 58, 1878–1890. [Google Scholar] [CrossRef]
- Teng, Y.H.; Aquino, R.S.; Park, P.W. Molecular functions of syndecan-1 in disease. Matrix Biol. 2012, 31, 3–16. [Google Scholar] [CrossRef]
- Hu, S.; Li, Z.; Lan, Y.; Guan, J.; Zhao, K.; Chu, D.; Fan, G.; Guo, Y.; Gao, F.; He, W. MiR-10a-5p-Mediated Syndecan 1 Suppression Restricts Porcine Hemagglutinating Encephalomyelitis Virus Replication. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Zhao, T.; Cui, L.; Yu, X.; Zhang, Z.; Shen, X.; Hua, X. Porcine sapelovirus enters PK-15 cells via caveolae-dependent endocytosis and requires Rab7 and Rab11. Virology 2019, 529, 160–168. [Google Scholar] [CrossRef]
- Lum, K.K.; Cristea, I.M. Proteomic approaches to uncovering virus–host protein interactions during the progression of viral infection. Expert Rev. Proteom. 2016, 13, 325–340. [Google Scholar] [CrossRef]
- Yang, F.; Li, W.; Derbyshire, M.; Larsen, M.R.; Rudd, J.J.; Palmisano, G. Unraveling incompatibility between wheat and the fungal pathogen Zymoseptoria tritici through apoplastic proteomics. BMC Genom. 2015, 16, 362. [Google Scholar] [CrossRef]
- Scaturro, P.; Stukalov, A.; Haas, D.A.; Cortese, M.; Draganova, K.; Plaszczyca, A.; Bartenschlager, R.; Gotz, M.; Pichlmair, A. An orthogonal proteomic survey uncovers novel Zika virus host factors. Nature 2018, 561, 253–257. [Google Scholar] [CrossRef]
- Liu, R.; Hu, X.; Lu, A.; Song, Y.; Lian, Z.; Sun, J.; Sung, Y.Y. Proteomic Profiling of Zebrafish Challenged by Spring Viremia of Carp Virus Provides Insight into Skin Antiviral Response. Zebrafish 2020, 16. [Google Scholar] [CrossRef] [PubMed]
- Fensterl, V.; Sen, G.C. The ISG56/IFIT1 gene family. J. Interferon Cytokine Res. 2011, 31, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Jin, H.; Wang, J. WITHDRAWN: Silencing of Interferon-Induced Transmembrane Protein 1 (IFITM1) Inhibits Proliferation, Migration, and Invasion in Lung Cancer Cells. Oncol Res. 2017, 20. [Google Scholar] [CrossRef]
- Terenzi, F.; Saikia, P.; Sen, G.C. Interferon-inducible protein, P56, inhibits HPV DNA replication by binding to the viral protein E1. Embo J. 2008, 27, 3311–3321. [Google Scholar] [CrossRef]
- Santhakumar, D.; Rohaim, M.; Hussein, H.A.; Hawes, P.; Ferreira, H.L.; Behboudi, S.; Iqbal, M.; Nair, V.; Arns, C.W.; Munir, M. Chicken Interferon-induced Protein with Tetratricopeptide Repeats 5 Antagonizes Replication of RNA Viruses. Sci. Rep. 2018, 8, 018–24905. [Google Scholar] [CrossRef]
- Baldanta, S.; Fernandez-Escobar, M.; Acin-Perez, R.; Albert, M.; Camafeita, E.; Jorge, I.; Vazquez, J.; Enriquez, J.A.; Guerra, S. ISG15 governs mitochondrial function in macrophages following vaccinia virus infection. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef]
- Zhao, C.; Hsiang, T.Y.; Kuo, R.L.; Krug, R.M. ISG15 conjugation system targets the viral NS1 protein in influenza A virus-infected cells. Proc. Natl Acad Sci USA 2010, 107, 2253–2258. [Google Scholar] [CrossRef]
- Gonzalez-Sanz, R.; Mata, M.; Bermejo-Martin, J.; Alvarez, A.; Cortijo, J.; Melero, J.A.; Martinez, I. ISG15 Is Upregulated in Respiratory Syncytial Virus Infection and Reduces Virus Growth through Protein ISGylation. J. Virol. 2016, 90, 3428–3438. [Google Scholar] [CrossRef]
- Liu, H.; Li, S.; Yang, X.; Wang, X.; Li, Y.; Wang, C.; Chen, L.; Chang, H. Porcine ISG15 modulates the antiviral response during pseudorabies virus replication. Gene 2018, 679, 212–218. [Google Scholar] [CrossRef]
- Chen, L.; Sun, J.; Meng, L.; Heathcote, J.; Edwards, A.M.; McGilvray, I.D. ISG15, a ubiquitin-like interferon-stimulated gene, promotes hepatitis C virus production in vitro: Implications for chronic infection and response to treatment. J. Gen. Virol. 2010, 91, 382–388. [Google Scholar] [CrossRef]
- Moal, V.; Textoris, J.; Ben Amara, A.; Mehraj, V.; Berland, Y.; Colson, P.; Mege, J.L. Chronic hepatitis E virus infection is specifically associated with an interferon-related transcriptional program. J. Infect. Dis 2013, 207, 125–132. [Google Scholar] [CrossRef]
- Syn, W.K.; Choi, S.S.; Liaskou, E.; Karaca, G.F.; Agboola, K.M.; Oo, Y.H.; Mi, Z.; Pereira, T.A.; Zdanowicz, M.; Malladi, P.; et al. Osteopontin is induced by hedgehog pathway activation and promotes fibrosis progression in nonalcoholic steatohepatitis. Hepatology 2011, 53, 106–115. [Google Scholar] [CrossRef]
- Brown, A.; Islam, T.; Adams, R.; Nerle, S.; Kamara, M.; Eger, C.; Marder, K.; Cohen, B.; Schifitto, G.; McArthur, J.C.; et al. Osteopontin enhances HIV replication and is increased in the brain and cerebrospinal fluid of HIV-infected individuals. J. Neurovirol. 2011, 17, 382–392. [Google Scholar] [CrossRef]
- Choi, S.S.; Claridge, L.C.; Jhaveri, R.; Swiderska-Syn, M.; Clark, P.; Suzuki, A.; Pereira, T.A.; Mi, Z.; Kuo, P.C.; Guy, C.D.; et al. Osteopontin is up-regulated in chronic hepatitis C and is associated with cellular permissiveness for hepatitis C virus replication. Clin. Sci. 2014, 126, 845–855. [Google Scholar] [CrossRef]
- Paul, A.M.; Acharya, D.; Duty, L.; Thompson, E.A.; Le, L.; Stokic, D.S.; Leis, A.A.; Bai, F. Osteopontin facilitates West Nile virus neuroinvasion via neutrophil “Trojan horse” transport. Sci. Rep. 2017, 7, 4722. [Google Scholar] [CrossRef]
- Ziegler, W.H.; Liddington, R.C.; Critchley, D.R. The structure and regulation of vinculin. Trends Cell Biol. 2006, 16, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jia, X.; Zhang, X.; Sun, J.; Lu, H. Proteomic analysis of PBMCs: Characterization of potential HIV-associated proteins. Proteome Sci. 2010, 8, 12. [Google Scholar] [CrossRef]
- Brown, C.; Morham, S.; Walsh, D.; Naghavi, M. Focal adhesion proteins talin-1 and vinculin negatively affect paxillin phosphorylation and limit retroviral infection. J. Mol. Biol. 2011, 410, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Reijmers, R.M.; Spaargaren, M.; Pals, S.T. Heparan sulfate proteoglycans in the control of B cell development and the pathogenesis of multiple myeloma. Febs J. 2013, 280, 2180–2193. [Google Scholar] [CrossRef]
- Hoffmann, C.; Tiemann, M.; Schrader, C.; Janssen, D.; Wolf, E.; Vierbuchen, M.; Parwaresch, R.; Ernestus, K.; Plettenberg, A.; Stoehr, A.; et al. AIDS-related B-cell lymphoma (ARL): Correlation of prognosis with differentiation profiles assessed by immunophenotyping. Blood 2005, 106, 1762–1769. [Google Scholar] [CrossRef]
- Dhodapkar, M.V.; Sanderson, R.D. Syndecan-1 (CD 138) in myeloma and lymphoid malignancies: A multifunctional regulator of cell behavior within the tumor microenvironment. Leuk Lymphoma 1999, 34, 35–43. [Google Scholar] [CrossRef]
- Hayashida, K.; Stahl, P.D.; Park, P.W. Syndecan-1 ectodomain shedding is regulated by the small GTPase Rab5. J. Biol. Chem. 2008, 283, 35435–35444. [Google Scholar] [CrossRef]
- Bacsa, S.; Karasneh, G.; Dosa, S.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Syndecan-1 and syndecan-2 play key roles in herpes simplex virus type-1 infection. J. Gen. Virol. 2011, 92, 733–743. [Google Scholar] [CrossRef]
- Freissler, E.; Meyer auf der Heyde, A.; David, G.; Meyer, T.F.; Dehio, C. Syndecan-1 and syndecan-4 can mediate the invasion of OpaHSPG-expressing Neisseria gonorrhoeae into epithelial cells. Cell Microbiol. 2000, 2, 69–82. [Google Scholar] [CrossRef]
- Wang, S.H.; Zhang, C.; Liao, C.P.; Lasbury, M.E.; Durant, P.J.; Tschang, D.; Lee, C.H. Syndecan-1 expression in the lung during Pneumocystis infection. J. Eukaryot Microbiol. 2006, 53, 1550–7408. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Vaeth, S.; Cuomo, L.; Boccellato, F.; Vincenti, S.; Cirone, M.; Presutti, C.; Junker, S.; Winberg, G.s.; Frati, L. Epstein–Barr virus infection leads to partial phenotypic reversion of terminally differentiated malignant B cells. Cancer lett. 2009, 284, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Couchman, J.R. Syndecans: Proteoglycan regulators of cell-surface microdomains? Nat. Rev. Mol. Cell Biol. 2003, 4, 926–937. [Google Scholar] [CrossRef]
- Letoha, T.; Kolozsi, C.; Ekes, C.; Keller-pintér, A.; Kusz, E.; Szakonyi, G.; Duda, E.; Szilák, L. Contribution of syndecans to lipoplex-mediated gene delivery. Eur. J. Pharm. Sci. 2013, 49, 550–555. [Google Scholar] [CrossRef]
- Hulst, M.M.; van Gennip, H.G.; Moormann, R.J. Passage of classical swine fever virus in cultured swine kidney cells selects virus variants that bind to heparan sulfate due to a single amino acid change in envelope protein E(rns). J. Virol. 2000, 74, 9553–9561. [Google Scholar] [CrossRef]
- Hulst, M.M.; van Gennip, H.G.; Vlot, A.C.; Schooten, E.; de Smit, A.J.; Moormann, R.J. Interaction of classical swine fever virus with membrane-associated heparan sulfate: Role for virus replication in vivo and virulence. J. Virol. 2001, 75, 9585–9595. [Google Scholar] [CrossRef]
- Grigorov, B.; Reungoat, E.; Gentil Dit Maurin, A.; Varbanov, M.; Blaising, J.; Michelet, M.; Manuel, R.; Parent, R.; Bartosch, B.; Zoulim, F.; et al. Hepatitis C virus infection propagates through interactions between Syndecan-1 and CD81 and impacts the hepatocyte glycocalyx. Cell Microbiol. 2017, 19, 23. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359. [Google Scholar] [CrossRef]
- Wei, Y.; Fang, C.-L.; Liu, S.-J.; Yang, W.-Q.; Wei, L.-S.; Lei, X.-J.; Hu, F.; Huang, H.-Y.; Li, W.; Chen, W. Long-term moderate exercise enhances specific proteins that constitute neurotrophin signaling pathway: A TMT-based quantitative proteomic analysis of rat plasma. J. Proteom. 2018, 185, 39–50. [Google Scholar] [CrossRef]
- Gluck, F.; Hoogland, C.; Antinori, P.; Robin, X.; Nikitin, F.; Zufferey, A.; Pasquarello, C.; Fétaud, V.; Dayon, L.; Müller, M.; et al. EasyProt--an easy-to-use graphical platform for proteomics data analysis. J. Proteomics 2013, 79, 146–160. [Google Scholar] [CrossRef]
- Chiang, D.Y.; Brown, P.O.; Eisen, M.B. Visualizing associations between genome sequences and gene expression data using genome-mean expression profiles. Bioinformatics 2001, 17, S49–S55. [Google Scholar] [CrossRef]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, 3. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, 17. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Coverage | Unique Peptides | Fold-Change Ratios | p-Value |
|---|---|---|---|---|---|
| Up-regulated | |||||
| A0A286ZSR7 | Olfactory receptor | 2.56 | 1 | 3.02 | 9.2167 × 10−5 |
| F1RTV4 | Amidophosphoribosyltransferase | 1.9 | 1 | 2.80 | 4.2788 × 10−5 |
| I3LEQ6 | Ral GTPase activating protein catalytic alpha subunit 2 | 2.13 | 1 | 2.06 | 0.0111 |
| F1RL90 | PPARG coactivator 1 beta | 1.56 | 1 | 1.88 | 0.0194 |
| A0A287BN67 | Nucleosome assembly protein 1 like 1 | 21.93 | 1 | 1.84 | 0.0020 |
| A0A287AZH1 | Nucleosome assembly protein 1 like 4 | 27.73 | 9 | 1.71 | 0.0017 |
| A0A287ATX4 | Wiskott-Aldrich syndrome like | 1.58 | 1 | 1.64 | 0.0054 |
| A0A286ZN07 | Charged multivesicular body protein 4A | 10.36 | 2 | 1.63 | 0.0007 |
| F1RRI4 | Sjogren syndrome/scleroderma autoantigen 1 | 7.54 | 2 | 1.56 | 0.0030 |
| F1SU38 | Urokinase-type plasminogen activator | 16.29 | 6 | 1.50 | 9.07 × 10−5 |
| S5A7T7 | SP110 nuclear body protein variant 2 | 8.67 | 1 | 1.46 | 0.0181 |
| F2Z572 | Phosphoribosyl pyrophosphate synthetase 1 | 18.55 | 2 | 1.44 | 0.0146 |
| A0A0B8RSH5 | Nuclear mitotic apparatus protein 1 | 45.07 | 2 | 1.44 | 0.0002 |
| F1RYV0 | Cyclin dependent kinase like 2 | 1.23 | 1 | 1.39 | 0.0438 |
| A0A287BRG2 | Required for excision 1-B domain containing | 4.15 | 1 | 1.39 | 0.0333 |
| O77637 | Transcription factor NFAT (Fragment) | 15.83 | 2 | 1.35 | 0.0028 |
| P26234 | Vinculin | 55.15 | 59 | 1.34 | 0.0028 |
| F1SCY0 | Interferon induced protein with tetratricopeptide repeats 5 | 6.02 | 3 | 1.33 | 0.0006 |
| Q4VK70 | Glycogen [starch] synthase (Fragment) | 3.91 | 2 | 1.33 | 0.0037 |
| A0A287AB58 | Ubiquitin-like modifier | 19.16 | 3 | 1.32 | 0.0095 |
| Down-regulated | |||||
| Q56PB8 | RPGR (Fragment) | 1.38 | 1 | 0.35 | 0.0012 |
| I3LQ36 | Uncharacterized protein | 7.38 | 1 | 0.60 | 0.0015 |
| A0A287BF68 | Nucleoporin 153 | 14.79 | 18 | 0.63 | 0.0007 |
| A0A0B8RSR5 | Interleukin enhancer binding factor 3 | 36.61 | 1 | 0.64 | 0.0044 |
| K7GNF5 | Syndecan | 4.85 | 1 | 0.65 | 0.0014 |
| F1SB63 | T-complex protein 1 subunit alpha | 54.92 | 1 | 0.65 | 0.0016 |
| A0A286ZQL5 | NOVA alternative splicing regulator 2 | 1.63 | 1 | 0.68 | 0.0043 |
| A0A287BPS0 | SET domain containing 2 | 1.4 | 3 | 0.68 | 0.0080 |
| A0A287ACY9 | Uncharacterized protein | 4.93 | 4 | 0.69 | 1.4560 × 10−5 |
| F1SFU1 | PHD finger protein 23 | 3.31 | 1 | 0.69 | 0.0016 |
| A0A286ZVL0 | Heterogeneous nuclear ribonucleoprotein U | 45.21 | 3 | 0.69 | 0.0025 |
| I3LIL0 | Nucleoporin like 2 | 4.31 | 1 | 0.70 | 2.6992 × 10−5 |
| Q8HZJ6 | Syndecan-4 | 35.64 | 5 | 0.70 | 0.0070 |
| C8ZL57 | Activating transcription factor 6 (Fragment) | 10.75 | 1 | 0.70 | 0.0001 |
| F1S8L9 | Heterogeneous nuclear ribonucleoprotein U | 44.65 | 2 | 0.70 | 0.0047 |
| F1SL26 | Tumor necrosis factor receptor superfamily member 1A | 2.63 | 1 | 0.71 | 0.0096 |
| F1SDY8 | Zinc finger CCCH-type containing 14 | 24.5 | 14 | 0.71 | 3.3193 × 10−5 |
| A0A286ZJK2 | Heterogeneous nuclear ribonucleoprotein M | 55.86 | 39 | 0.72 | 0.0005 |
| I3LLM2 | Olfactory receptor | 2.31 | 1 | 0.73 | 0.0010 |
| F1SD01 | Cleavage stimulation factor subunit 2 tau variant | 11.87 | 3 | 0.73 | 0.0014 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, T.; Cui, L.; Yu, X.; Zhang, Z.; Chen, Q.; Hua, X. Proteome Analysis Reveals Syndecan 1 Regulates Porcine Sapelovirus Replication. Int. J. Mol. Sci. 2020, 21, 4386. https://doi.org/10.3390/ijms21124386
Zhao T, Cui L, Yu X, Zhang Z, Chen Q, Hua X. Proteome Analysis Reveals Syndecan 1 Regulates Porcine Sapelovirus Replication. International Journal of Molecular Sciences. 2020; 21(12):4386. https://doi.org/10.3390/ijms21124386
Chicago/Turabian StyleZhao, Tingting, Li Cui, Xiangqian Yu, Zhonghai Zhang, Qi Chen, and Xiuguo Hua. 2020. "Proteome Analysis Reveals Syndecan 1 Regulates Porcine Sapelovirus Replication" International Journal of Molecular Sciences 21, no. 12: 4386. https://doi.org/10.3390/ijms21124386
APA StyleZhao, T., Cui, L., Yu, X., Zhang, Z., Chen, Q., & Hua, X. (2020). Proteome Analysis Reveals Syndecan 1 Regulates Porcine Sapelovirus Replication. International Journal of Molecular Sciences, 21(12), 4386. https://doi.org/10.3390/ijms21124386