Secretome Analysis of Mesenchymal Stem Cell Factors Fostering Oligodendroglial Differentiation of Neural Stem Cells In Vivo

 , , , , ,
, , , , ,  ,
, 
Abstract
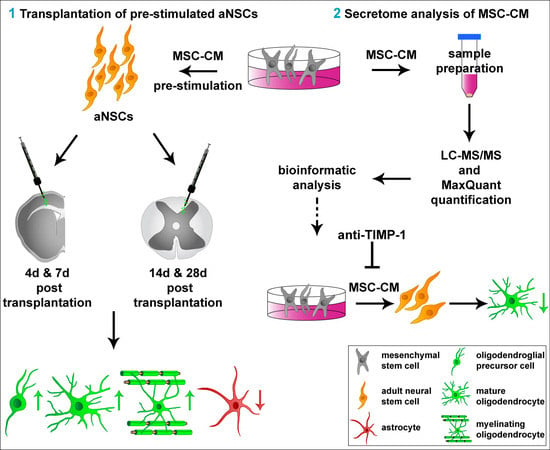
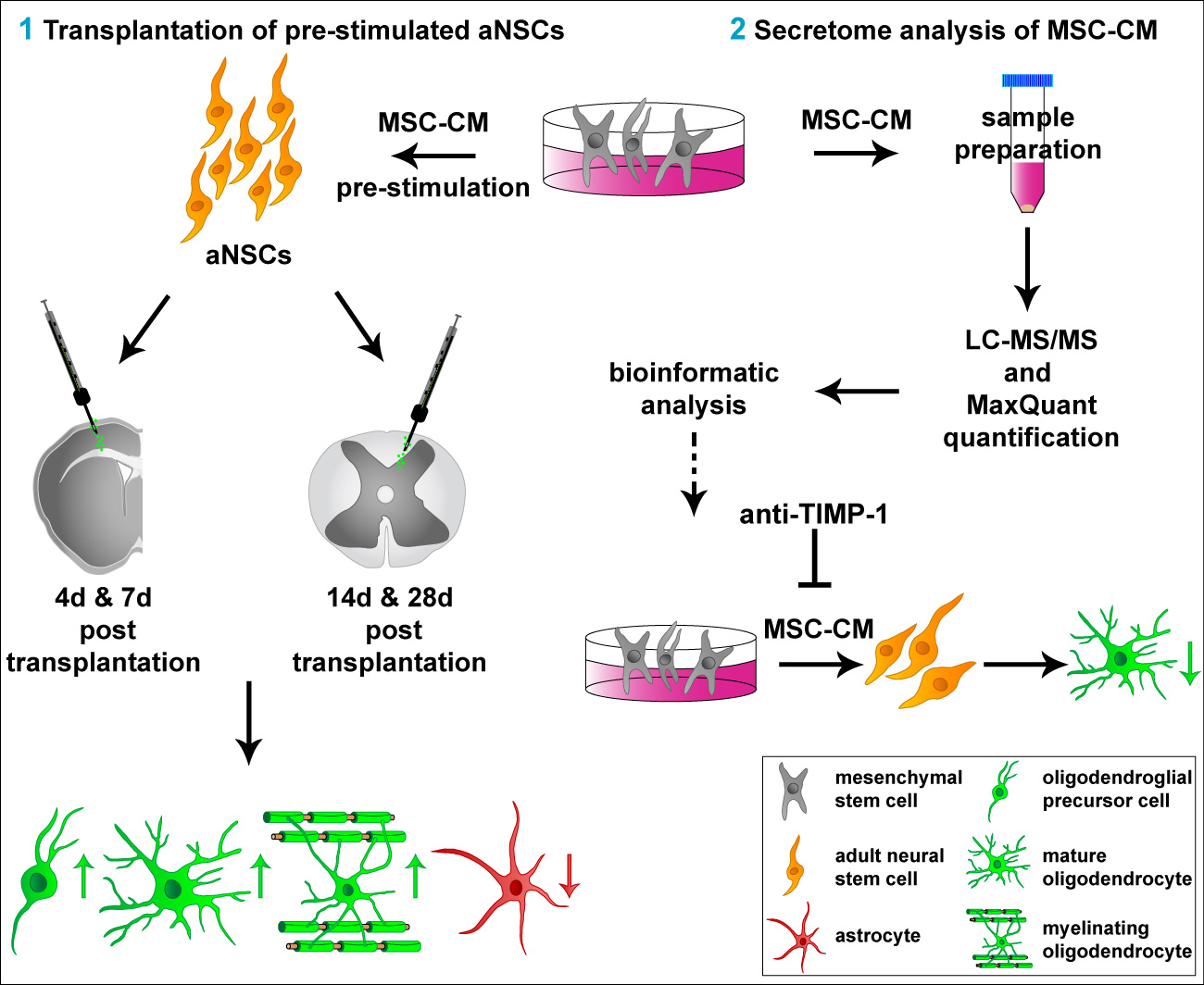
1. Introduction
2. Results
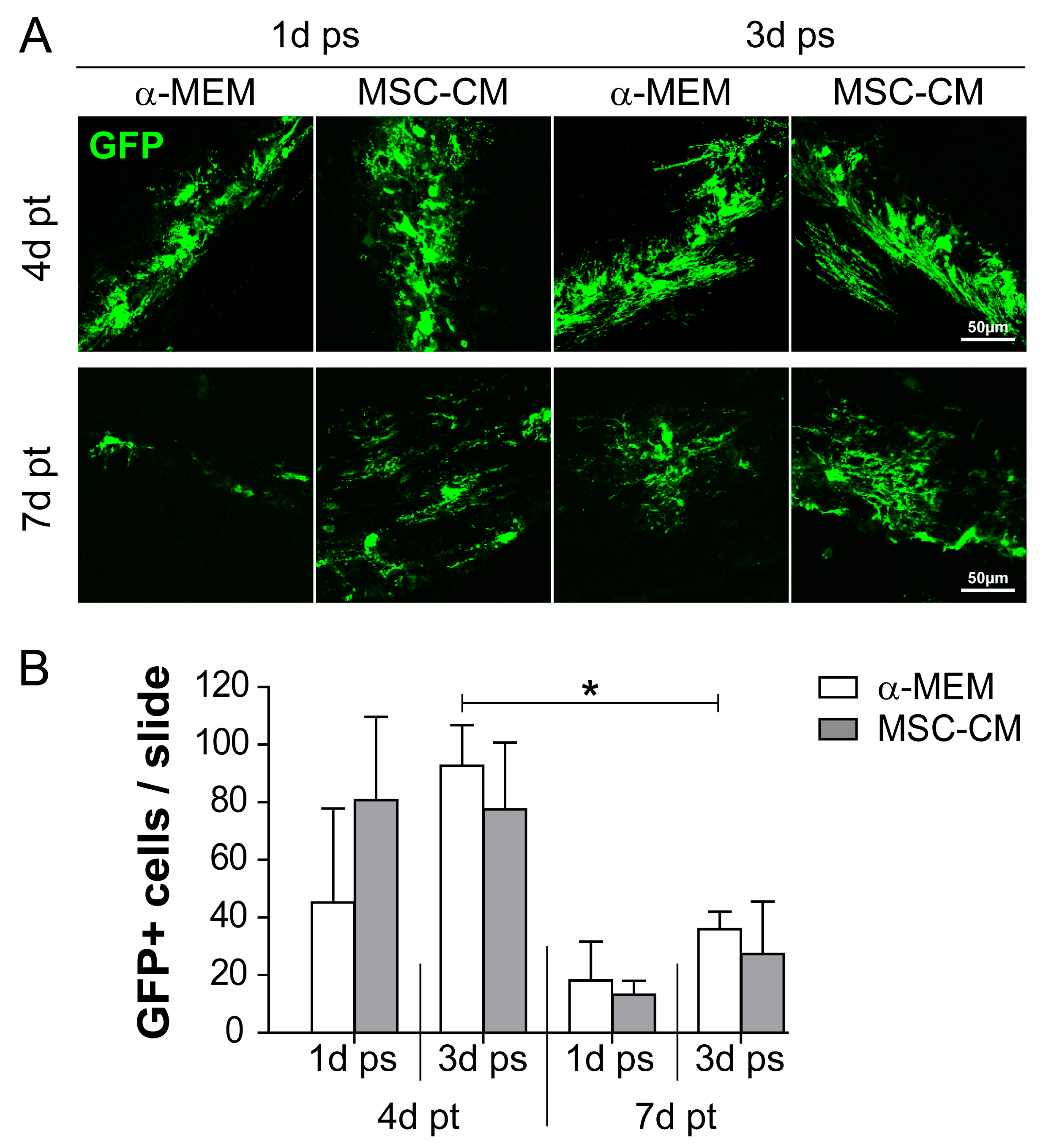
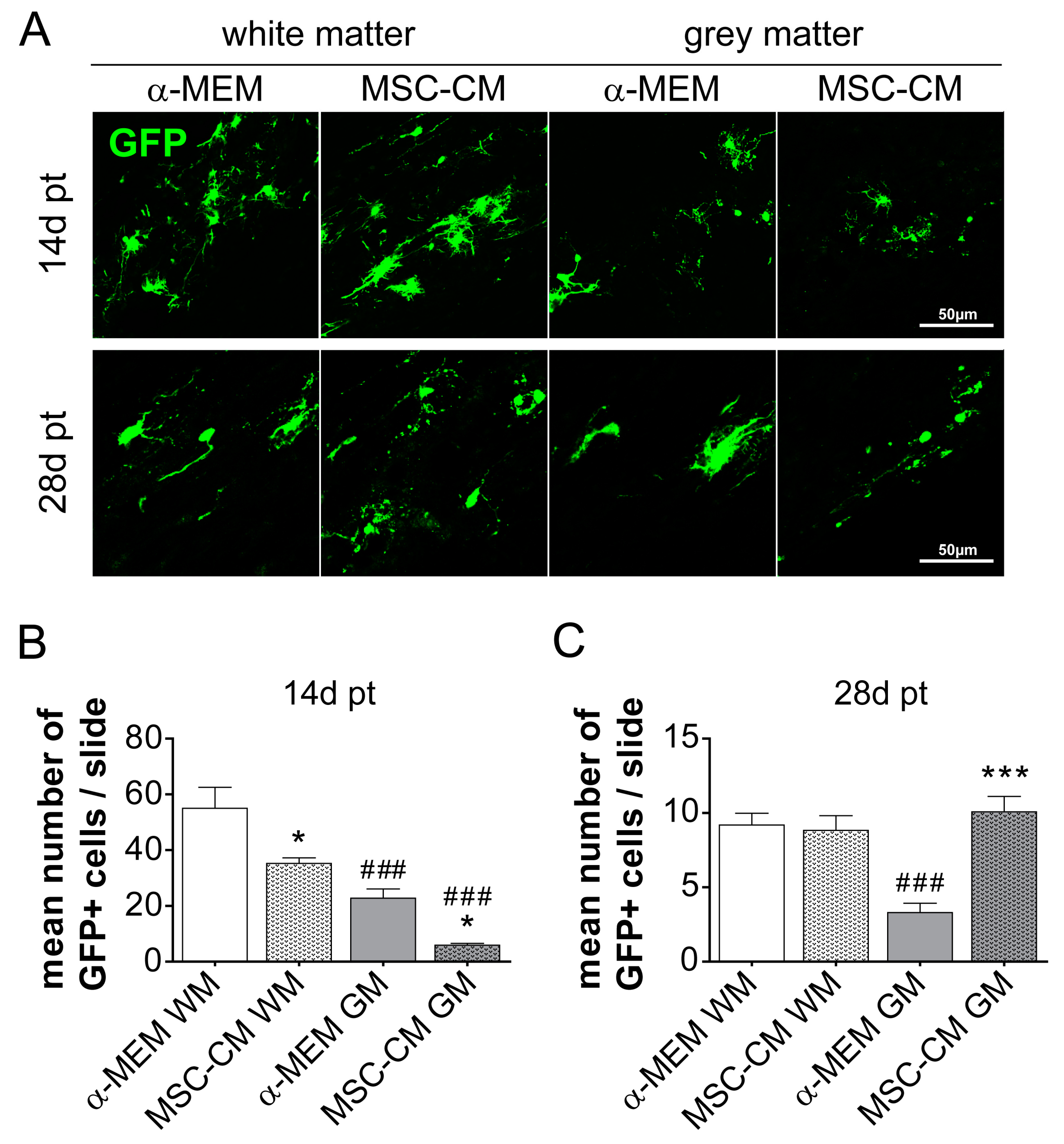
2.1. Survival of MSC-CM-Stimulated aNSC In Vivo
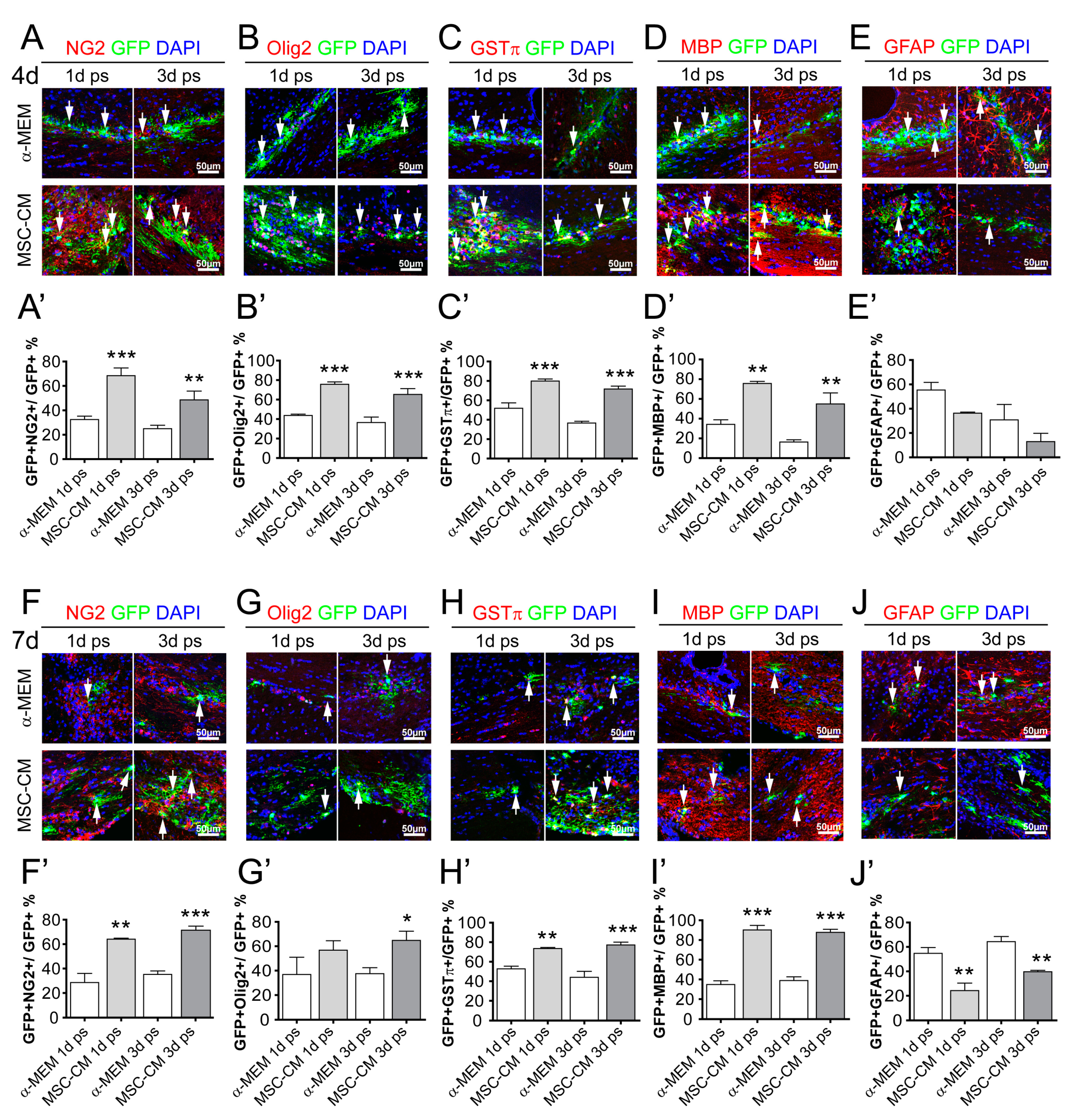
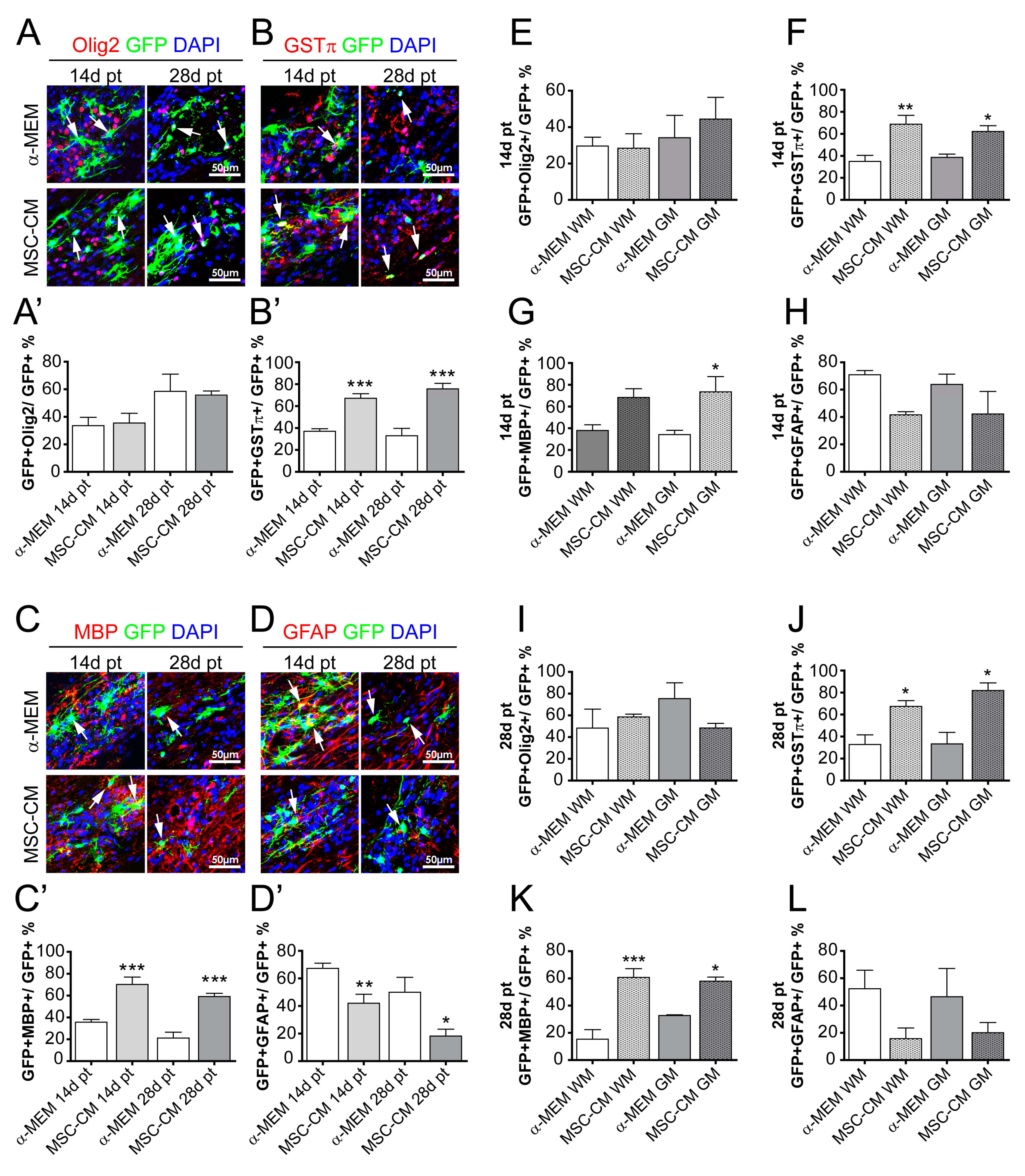
2.2. MSC-Derived Factors Promoted the Oligodendroglial Differentiation Process In Vivo
2.3. Extended Survival Time of MSC-CM Pre-Stimulated aNSCs Transplanted into the Adult Rat Spinal Cord
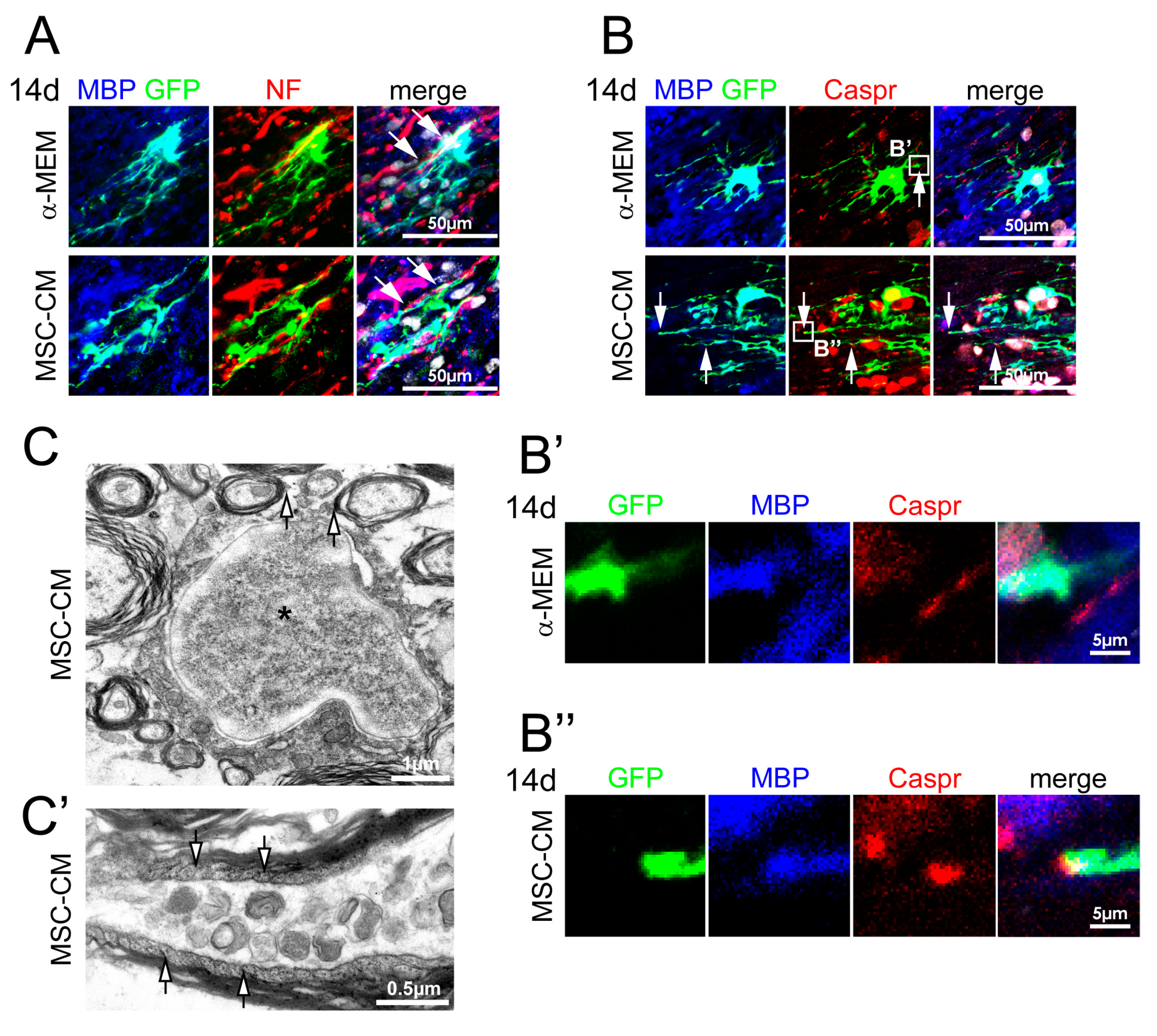
2.4. MSC-CM Pre-Stimulation Enabled aNSCs to Myelinate Axons In Vivo
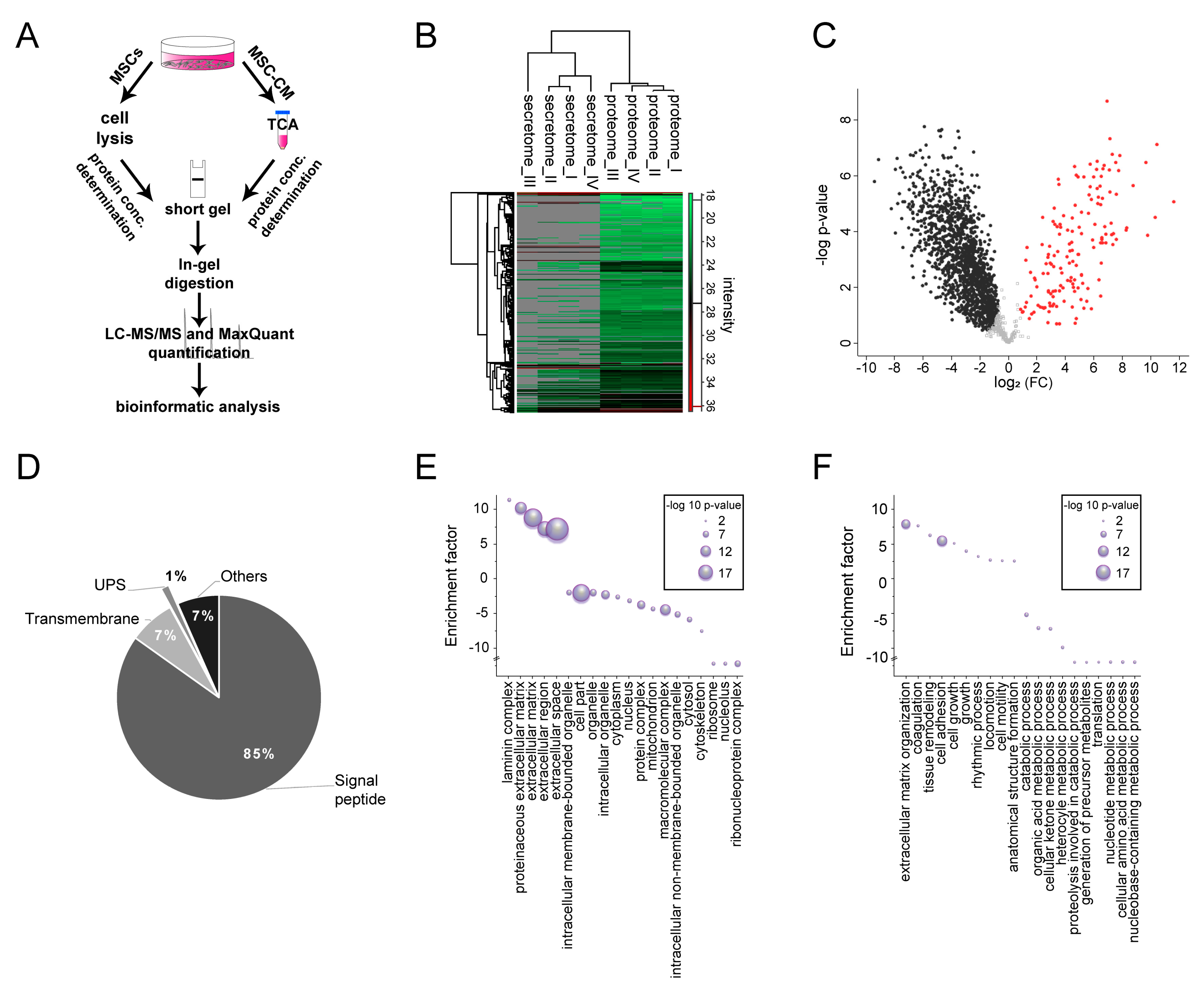
2.5. Characterization of Adult Rat Mesenchymal Stem Cell Secretome
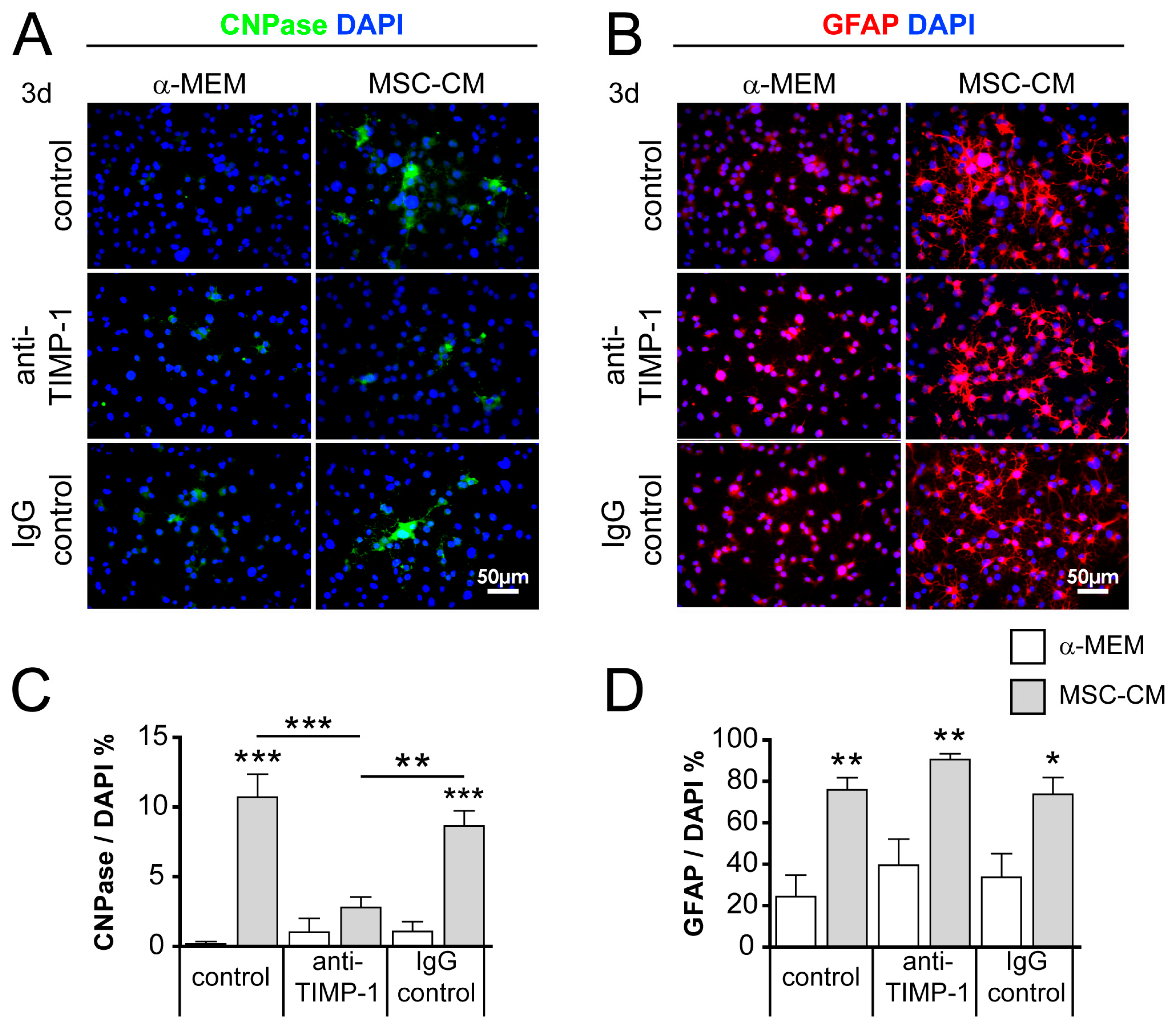
2.6. Neutralization of TIMP-1 Diminished the Pro-Oligodendroglial Effect of MSC-CM
3. Discussion
3.1. Enhanced Oligodendroglial Differentiation of MSC-CM-Stimulated aNSCs In Vivo
3.2. MSCs Secreted a Number of Oligodendroglial Regulatory Proteins
4. Materials and Methods
4.1. Animals
4.2. Mesenchymal Stem Cell Culture
4.3. Adult Rat Neural Stem Cell Culture and Transfection
4.4. Antibody Blocking and Immunocytochemistry
4.5. Stereotactic Cell Transplantations
4.6. Immunohistochemistry
4.7. Immunoelectron Microscopy
4.8. Statistical Analysis of IHC and ICC
4.9. Preparation of Mesenchymal Stem Cell Secretome and Proteome
4.10. Liquid Chromatography and Mass Spectrometric Analysis
4.11. Analysis of Mass Spectrometric Data
4.12. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-MEM | Alpha-minimum essential medium |
| Acan | Aggrecan |
| ANOVA | Analysis of variance |
| aNSCs | Adult neural stem cells |
| BMP | Bone morphogenetic protein |
| BSA | Bovine serum albumin |
| Caspr | Contactin-associated protein |
| Chrd | Chordin |
| CLSM | Confocal laser scanning microscopy |
| CNPase | 2′,3′-Cyclic-nucleotide 3′-phosphodiesterase |
| CNS | Central nervous system |
| CTGF | Connective tissue growth factor |
| Dag1 | Dystroglycan |
| DAPI | 4´,6-diamidin-2-phenylindol |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DTT | Dithiothreitol |
| ECM | Extracellular matrix |
| EF | Enrichment factor |
| EGF | Epidermal growth factor |
| EM | Electron microscopy |
| FCS | Fetal calf serum |
| FDR | False discovery rate |
| FGF | Fibroblast growth factor |
| Fn | Fibronectin |
| Gas6 | Growth arrest-specific protein 6 |
| GFAP | Glial fibrillary acidic protein |
| GFP | Green fluorescent protein |
| GM | Grey matter |
| GSTπ | Glutathione-S-transferase-π |
| GO | Gene Ontology |
| GOBP | Gene Ontology biological process |
| GOCC | Gene Ontology cellular component |
| IGFBP | Insulin-like growth factor-binding protein |
| IgG | Immunoglobulin G |
| Lamb2 | Laminin subunit beta-2 |
| LC–MS/MS | Liquid chromatography–mass spectrometry/mass spectrometry |
| MBP | Myelin basic protein |
| MMP | Matrix metalloproteinase |
| MSCs | Mesenchymal stem cells |
| MSC-CM | Mesenchymal stem cell conditioned medium |
| NB | Neurobasal |
| NF | Neurofilament |
| NG2 | Neural/glial antigen 2 |
| NGS | Normal goat serum |
| Olig2 | Oligodendrocyte transcription factor 2 |
| OPCs | Oligodendrocyte progenitor cells |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| ps | Pre-stimulated |
| Psap | Prospaposin |
| pt | Post transplantation |
| Sdcbp | Syntenin-1 |
| SP | Signal peptide |
| Spp1 | Osteopontin |
| SVZ | Subventricular zone |
| TCA | Trichloroacetic acid |
| Thbs1 | Thrombospondin 1 |
| TIMP-1 | Tissue inhibitor of metalloproteinase type 1 |
| TM | Transmembrane |
| Tnc | Tenascin c |
| UPS | Unconventional protein secretion |
| WM | White matter |
References
- Franklin, R.J. Why does remyelination fail in multiple sclerosis? Nat. Rev. Neurosci. 2002, 3, 705–714. [Google Scholar] [CrossRef]
- Jessberger, S.; Toni, N.; Clemenson, G.D., Jr.; Ray, J.; Gage, F.H. Directed differentiation of hippocampal stem/progenitor cells in the adult brain. Nat. Neurosci. 2008, 11, 888–893. [Google Scholar] [CrossRef]
- Akkermann, R.; Jadasz, J.J.; Azim, K.; Küry, P. Taking Advantage of Nature’s Gift: Can Endogenous Neural Stem Cells Improve Myelin Regeneration? Int. J. Mol. Sci. 2016, 17, 1895. [Google Scholar] [CrossRef]
- Nait-Oumesmar, B.; Picard-Riera, N.; Kerninon, C.; Decker, L.; Seilhean, D.; Hoglinger, G.U.; Hirsch, E.C.; Reynolds, R.; Baron-Van Evercooren, A. Activation of the subventricular zone in multiple sclerosis: Evidence for early glial progenitors. Proc. Natl. Acad. Sci. USA 2007, 104, 4694–4699. [Google Scholar] [CrossRef]
- Nait-Oumesmar, B.; Picard-Riera, N.; Kerninon, C.; Baron-Van Evercooren, A. The role of SVZ-derived neural precursors in demyelinating diseases: From animal models to multiple sclerosis. J. Neurol. Sci. 2008, 265, 26–31. [Google Scholar] [CrossRef]
- Picard-Riera, N.; Decker, L.; Delarasse, C.; Goude, K.; Nait-Oumesmar, B.; Liblau, R.; Pham-Dinh, D.; Baron-Van Evercooren, A. Experimental autoimmune encephalomyelitis mobilizes neural progenitors from the subventricular zone to undergo oligodendrogenesis in adult mice. Proc. Natl. Acad. Sci. USA 2002, 99, 13211–13216. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.L.; Roth, P.T.; Stratton, J.A.; Chuang, B.H.; Danne, J.; Ellis, S.L.; Ng, S.W.; Kilpatrick, T.J.; Merson, T.D. Adult neural precursor cells from the subventricular zone contribute significantly to oligodendrocyte regeneration and remyelination. J. Neurosci. 2014, 34, 14128–14146. [Google Scholar] [CrossRef] [PubMed]
- Charles, P.; Reynolds, R.; Seilhean, D.; Rougon, G.; Aigrot, M.S.; Niezgoda, A.; Zalc, B.; Lubetzki, C. Re-expression of PSA-NCAM by demyelinated axons: An inhibitor of remyelination in multiple sclerosis? Brain 2002, 125, 1972–1979. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Miller, R.H.; Lee, X.; Scott, M.L.; Shulag-Morskaya, S.; Shao, Z.; Chang, J.; Thill, G.; Levesque, M.; Zhang, M.; et al. LINGO-1 negatively regulates myelination by oligodendrocytes. Nat. Neurosci. 2005, 8, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Pepinsky, R.B.; Cadavid, D. Blocking LINGO-1 as a therapy to promote CNS repair: From concept to the clinic. CNS Drugs 2013, 27, 493–503. [Google Scholar] [CrossRef]
- De Castro, F.; Bribian, A.; Ortega, M.C. Regulation of oligodendrocyte precursor migration during development, in adulthood and in pathology. Cell Mol. Life Sci. 2013, 70, 4355–4368. [Google Scholar] [CrossRef] [PubMed]
- Kremer, D.; Aktas, O.; Hartung, H.P.; Küry, P. The complex world of oligodendroglial differentiation inhibitors. Ann. Neurol. 2011, 69, 602–618. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, T.; Miron, V.; Cui, Q.; Wegner, C.; Antel, J.; Bruck, W. Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 2008, 131, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Gruchot, J.; Weyers, V.; Göttle, P.; Förster, M.; Hartung, H.P.; Küry, P.; Kremer, D. The Molecular Basis for Remyelination Failure in Multiple Sclerosis. Cells 2019, 8, 825. [Google Scholar] [CrossRef] [PubMed]
- Kotter, M.R.; Stadelmann, C.; Hartung, H.P. Enhancing remyelination in disease--can we wrap it up? Brain 2011, 134, 1882–1900. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Rivera, F.J.; Couillard-Despres, S.; Pedre, X.; Ploetz, S.; Caioni, M.; Lois, C.; Bogdahn, U.; Aigner, L. Mesenchymal stem cells instruct oligodendrogenic fate decision on adult neural stem cells. Stem Cells 2006, 24, 2209–2219. [Google Scholar] [CrossRef]
- Kassis, I.; Grigoriadis, N.; Gowda-Kurkalli, B.; Mizrachi-Kol, R.; Ben-Hur, T.; Slavin, S.; Abramsky, O.; Karussis, D. Neuroprotection and immunomodulation with mesenchymal stem cells in chronic experimental autoimmune encephalomyelitis. Arch. Neurol. 2008, 65, 753–761. [Google Scholar] [CrossRef]
- Jadasz, J.J.; Kremer, D.; Göttle, P.; Tzekova, N.; Domke, J.; Rivera, F.J.; Adjaye, J.; Hartung, H.P.; Aigner, L.; Küry, P. Mesenchymal stem cell conditioning promotes rat oligodendroglial cell maturation. PLoS ONE 2013, 8, e71814. [Google Scholar] [CrossRef]
- Jadasz, J.J.; Tepe, L.; Beyer, F.; Samper Agrelo, I.; Akkermann, R.; Spitzhorn, L.S.; Silva, M.E.; Oreffo, R.O.C.; Hartung, H.P.; Prigione, A.; et al. Human mesenchymal factors induce rat hippocampal- and human neural stem cell dependent oligodendrogenesis. Glia 2018, 66, 145–160. [Google Scholar] [CrossRef]
- Rivera, F.J.; Kandasamy, M.; Couillard-Despres, S.; Caioni, M.; Sanchez, R.; Huber, C.; Weidner, N.; Bogdahn, U.; Aigner, L. Oligodendrogenesis of adult neural progenitors: Differential effects of ciliary neurotrophic factor and mesenchymal stem cell derived factors. J. Neurochem. 2008, 107, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Schira-Heinen, J.; Grube, L.; Waldera-Lupa, D.M.; Baberg, F.; Langini, M.; Etemad-Parishanzadeh, O.; Poschmann, G.; Stühler, K. Pitfalls and opportunities in the characterization of unconventionally secreted proteins by secretome analysis. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140237. [Google Scholar] [CrossRef] [PubMed]
- Maffioli, E.; Nonnis, S.; Angioni, R.; Santagata, F.; Cali, B.; Zanotti, L.; Negri, A.; Viola, A.; Tedeschi, G. Proteomic analysis of the secretome of human bone marrow-derived mesenchymal stem cells primed by pro-inflammatory cytokines. J. Proteom. 2017, 166, 115–126. [Google Scholar] [CrossRef]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Baberg, F.; Geyh, S.; Waldera-Lupa, D.; Stefanski, A.; Zilkens, C.; Haas, R.; Schroeder, T.; Stühler, K. Secretome analysis of human bone marrow derived mesenchymal stromal cells. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 434–441. [Google Scholar] [CrossRef]
- Beyer, F.; Jadasz, J.; Samper Agrelo, I.; Schira-Heinen, J.; Groh, J.; Manousi, A.; Butermann, C.; Estrada, V.; Reiche, L.; Cantone, M.; et al. Heterogeneous fate choice of genetically modulated adult neural stem cells in gray and white matter of the central nervous system. Glia 2020, 68, 393–406. [Google Scholar] [CrossRef]
- Samanta, J.; Grund, E.M.; Silva, H.M.; Lafaille, J.J.; Fishell, G.; Salzer, J.L. Inhibition of Gli1 mobilizes endogenous neural stem cells for remyelination. Nature 2015, 526, 448–452. [Google Scholar] [CrossRef]
- Koutsoudaki, P.N.; Papastefanaki, F.; Stamatakis, A.; Kouroupi, G.; Xingi, E.; Stylianopoulou, F.; Matsas, R. Neural stem/progenitor cells differentiate into oligodendrocytes, reduce inflammation, and ameliorate learning deficits after transplantation in a mouse model of traumatic brain injury. Glia 2016, 64, 763–779. [Google Scholar] [CrossRef]
- Grube, L.; Dellen, R.; Kruse, F.; Schwender, H.; Stühler, K.; Poschmann, G. Mining the Secretome of C2C12 Muscle Cells: Data Dependent Experimental Approach To Analyze Protein Secretion Using Label-Free Quantification and Peptide Based Analysis. J. Proteome Res. 2018, 17, 879–890. [Google Scholar] [CrossRef]
- Keough, M.B.; Rogers, J.A.; Zhang, P.; Jensen, S.K.; Stephenson, E.L.; Chen, T.; Hurlbert, M.G.; Lau, L.W.; Rawji, K.S.; Plemel, J.R.; et al. An inhibitor of chondroitin sulfate proteoglycan synthesis promotes central nervous system remyelination. Nat. Commun. 2016, 7, 11312. [Google Scholar] [CrossRef]
- Jablonska, B.; Aguirre, A.; Raymond, M.; Szabo, G.; Kitabatake, Y.; Sailor, K.A.; Ming, G.L.; Song, H.; Gallo, V. Chordin-induced lineage plasticity of adult SVZ neuroblasts after demyelination. Nat. Neurosci. 2010, 13, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ercan, E.; Han, J.M.; Di Nardo, A.; Winden, K.; Han, M.J.; Hoyo, L.; Saffari, A.; Leask, A.; Geschwind, D.H.; Sahin, M. Neuronal CTGF/CCN2 negatively regulates myelination in a mouse model of tuberous sclerosis complex. J. Exp. Med. 2017, 214, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Leiton, C.V.; Aranmolate, A.; Eyermann, C.; Menezes, M.J.; Escobar-Hoyos, L.F.; Husain, S.; Winder, S.J.; Colognato, H. Laminin promotes metalloproteinase-mediated dystroglycan processing to regulate oligodendrocyte progenitor cell proliferation. J. Neurochem. 2015, 135, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Colognato, H.; Galvin, J.; Wang, Z.; Relucio, J.; Nguyen, T.; Harrison, D.; Yurchenco, P.D.; Ffrench-Constant, C. Identification of dystroglycan as a second laminin receptor in oligodendrocytes, with a role in myelination. Development 2007, 134, 1723–1736. [Google Scholar] [CrossRef]
- Lourenco, T.; Paes de Faria, J.; Bippes, C.A.; Maia, J.; Lopes-da-Silva, J.A.; Relvas, J.B.; Graos, M. Modulation of oligodendrocyte differentiation and maturation by combined biochemical and mechanical cues. Sci. Rep. 2016, 6, 21563. [Google Scholar] [CrossRef]
- Shankar, S.L.; O’Guin, K.; Kim, M.; Varnum, B.; Lemke, G.; Brosnan, C.F.; Shafit-Zagardo, B. Gas6/Axl signaling activates the phosphatidylinositol 3-kinase/Akt1 survival pathway to protect oligodendrocytes from tumor necrosis factor alpha-induced apoptosis. J. Neurosci. 2006, 26, 5638–5648. [Google Scholar] [CrossRef]
- Kuhl, N.M.; De Keyser, J.; De Vries, H.; Hoekstra, D. Insulin-like growth factor binding proteins-1 and -2 differentially inhibit rat oligodendrocyte precursor cell survival and differentiation in vitro. J. Neurosci. Res. 2002, 69, 207–216. [Google Scholar] [CrossRef]
- Li, N.; Han, J.; Tang, J.; Ying, Y. IGFBP-7 inhibits the differentiation of oligodendrocyte precursor cells via regulation of Wnt/beta-Catenin signaling. J. Cell Biochem. 2018, 119, 4742–4750. [Google Scholar] [CrossRef]
- Kim, W.K.; Kim, D.; Cui, J.; Jang, H.H.; Kim, K.S.; Lee, H.J.; Kim, S.U.; Ahn, S.M. Secretome analysis of human oligodendrocytes derived from neural stem cells. PLoS ONE 2014, 9, e84292. [Google Scholar] [CrossRef]
- Relucio, J.; Menezes, M.J.; Miyagoe-Suzuki, Y.; Takeda, S.; Colognato, H. Laminin regulates postnatal oligodendrocyte production by promoting oligodendrocyte progenitor survival in the subventricular zone. Glia 2012, 60, 1451–1467. [Google Scholar] [CrossRef]
- See, J.; Zhang, X.; Eraydin, N.; Mun, S.B.; Mamontov, P.; Golden, J.A.; Grinspan, J.B. Oligodendrocyte maturation is inhibited by bone morphogenetic protein. Mol. Cell Neurosci. 2004, 26, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.S.; Milner, R.; Nishiyama, A.; Frausto, R.F.; Serwanski, D.R.; Pagarigan, R.R.; Whitton, J.L.; Miller, R.H.; Crocker, S.J. Astrocytic tissue inhibitor of metalloproteinase-1 (TIMP-1) promotes oligodendrocyte differentiation and enhances CNS myelination. J. Neurosci. 2011, 31, 6247–6254. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, A.M.; Johnson, K.M.; Willis, C.M.; Guzzo, R.M.; Crocker, S.J. TIMP-1 Promotes Oligodendrocyte Differentiation Through Receptor-Mediated Signaling. Mol. Neurobiol. 2019, 56, 3380–3392. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, R.; Bernasconi, L.; Losberger, C.; Graber, P.; Kadi, L.; Avellana-Adalid, V.; Picard-Riera, N.; Baron Van Evercooren, A.; Cirillo, R.; Kosco-Vilbois, M.; et al. Osteopontin is upregulated during in vivo demyelination and remyelination and enhances myelin formation in vitro. Mol. Cell Neurosci. 2004, 25, 707–721. [Google Scholar] [CrossRef]
- Meyer, R.C.; Giddens, M.M.; Coleman, B.M.; Hall, R.A. The protective role of prosaposin and its receptors in the nervous system. Brain Res. 2014, 1585, 1–12. [Google Scholar] [CrossRef]
- Hiraiwa, M.; Campana, W.M.; Mizisin, A.P.; Mohiuddin, L.; O’Brien, J.S. Prosaposin: A myelinotrophic protein that promotes expression of myelin constituents and is secreted after nerve injury. Glia 1999, 26, 353–360. [Google Scholar] [CrossRef]
- Chatterjee, N.; Stegmüller, J.; Schätzle, P.; Karram, K.; Koroll, M.; Werner, H.B.; Nave, K.A.; Trotter, J. Interaction of syntenin-1 and the NG2 proteoglycan in migratory oligodendrocyte precursor cells. J. Biol. Chem. 2008, 283, 8310–8317. [Google Scholar] [CrossRef]
- Lu, Z.; Kipnis, J. Thrombospondin 1--a key astrocyte-derived neurogenic factor. FASEB J. 2010, 24, 1925–1934. [Google Scholar] [CrossRef]
- Czopka, T.; von Holst, A.; ffrench-Constant, C.; Faissner, A. Regulatory mechanisms that mediate tenascin C-dependent inhibition of oligodendrocyte precursor differentiation. J. Neurosci. 2010, 30, 12310–12322. [Google Scholar] [CrossRef]
- Jiang, P.; Chen, C.; Liu, X.B.; Pleasure, D.E.; Liu, Y.; Deng, W. Human iPSC-Derived Immature Astroglia Promote Oligodendrogenesis by Increasing TIMP-1 Secretion. Cell Rep. 2016, 15, 1303–1315. [Google Scholar] [CrossRef]
- Steffenhagen, C.; Dechant, F.X.; Oberbauer, E.; Furtner, T.; Weidner, N.; Küry, P.; Aigner, L.; Rivera, F.J. Mesenchymal stem cells prime proliferating adult neural progenitors toward an oligodendrocyte fate. Stem Cells Dev. 2012, 21, 1838–1851. [Google Scholar] [CrossRef]
- Fricker, R.A.; Carpenter, M.K.; Winkler, C.; Greco, C.; Gates, M.A.; Bjorklund, A. Site-specific migration and neuronal differentiation of human neural progenitor cells after transplantation in the adult rat brain. J. Neurosci. 1999, 19, 5990–6005. [Google Scholar] [CrossRef] [PubMed]
- McGinley, L.M.; Kashlan, O.N.; Chen, K.S.; Bruno, E.S.; Hayes, J.M.; Backus, C.; Feldman, S.; Kashlan, B.N.; Johe, K.; Feldman, E.L. Human neural stem cell transplantation into the corpus callosum of Alzheimer’s mice. Ann. Clin. Transl. Neurol. 2017, 4, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C.M.; Fehlings, M.G. Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cord injury. J. Neurosci. 2006, 26, 3377–3389. [Google Scholar] [CrossRef]
- Vigano, F.; Möbius, W.; Götz, M.; Dimou, L. Transplantation reveals regional differences in oligodendrocyte differentiation in the adult brain. Nat. Neurosci. 2013, 16, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, M.; Shinomiya, K.; Okabe, S. Migration and differentiation of neural progenitor cells from two different regions of embryonic central nervous system after transplantation into the intact spinal cord. Eur. J. Neurosci. 2003, 17, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.A.; Novitch, B.G.; Goldman, J.E. Olig2 directs astrocyte and oligodendrocyte formation in postnatal subventricular zone cells. J. Neurosci. 2005, 25, 7289–7298. [Google Scholar] [CrossRef]
- Jungbluth, P.; Spitzhorn, L.S.; Grassmann, J.; Tanner, S.; Latz, D.; Rahman, M.S.; Bohndorf, M.; Wruck, W.; Sager, M.; Grotheer, V.; et al. Human iPSC-derived iMSCs improve bone regeneration in mini-pigs. Bone Res. 2019, 7, 32. [Google Scholar] [CrossRef]
- Spitzhorn, L.S.; Rahman, M.S.; Schwindt, L.; Ho, H.T.; Wruck, W.; Bohndorf, M.; Wehrmeyer, S.; Ncube, A.; Beyer, I.; Hagenbeck, C.; et al. Isolation and Molecular Characterization of Amniotic Fluid-Derived Mesenchymal Stem Cells Obtained from Caesarean Sections. Stem. Cells Int. 2017, 2017, 5932706. [Google Scholar] [CrossRef]
- Timaner, M.; Tsai, K.K.; Shaked, Y. The multifaceted role of mesenchymal stem cells in cancer. Semin. Cancer Biol. 2020, 60, 225–237. [Google Scholar] [CrossRef]
- Chun, S.J.; Rasband, M.N.; Sidman, R.L.; Habib, A.A.; Vartanian, T. Integrin-linked kinase is required for laminin-2-induced oligodendrocyte cell spreading and CNS myelination. J. Cell Biol. 2003, 163, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.E.; Lange, S.; Hinrichsen, B.; Philp, A.R.; Reyes, C.R.; Halabi, D.; Mansilla, J.B.; Rotheneichner, P.; Guzman de la Fuente, A.; Couillard-Despres, S.; et al. Pericytes Favor Oligodendrocyte Fate Choice in Adult Neural Stem Cells. Front. Cell Neurosci. 2019, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- Jadasz, J.J.; Rivera, F.J.; Taubert, A.; Kandasamy, M.; Sandner, B.; Weidner, N.; Aktas, O.; Hartung, H.P.; Aigner, L.; Küry, P. p57kip2 regulates glial fate decision in adult neural stem cells. Development 2012, 139, 3306–3315. [Google Scholar] [CrossRef]
- Binder, M.D.; Cate, H.S.; Prieto, A.L.; Kemper, D.; Butzkueven, H.; Gresle, M.M.; Cipriani, T.; Jokubaitis, V.G.; Carmeliet, P.; Kilpatrick, T.J. Gas6 deficiency increases oligodendrocyte loss and microglial activation in response to cuprizone-induced demyelination. J. Neurosci. 2008, 28, 5195–5206. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.K.; Heywood, D.; Ralph, G.S.; Bienemann, A.; Baker, A.H.; Uney, J.B. Tissue inhibitor of metalloproteinase 1 inhibits excitotoxic cell death in neurons. Mol. Cell Neurosci. 2003, 22, 98–106. [Google Scholar] [CrossRef]
- Figueres-Onate, M.; Garcia-Marques, J.; Lopez-Mascaraque, L. UbC-StarTrack, a clonal method to target the entire progeny of individual progenitors. Sci. Rep. 2016, 6, 33896. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Schira, J.; Gasis, M.; Estrada, V.; Hendricks, M.; Schmitz, C.; Trapp, T.; Kruse, F.; Kögler, G.; Wernet, P.; Hartung, H.P.; et al. Significant clinical, neuropathological and behavioural recovery from acute spinal cord trauma by transplantation of a well-defined somatic stem cell from human umbilical cord blood. Brain 2012, 135, 431–446. [Google Scholar] [CrossRef]
- Schira, J.; Falkenberg, H.; Hendricks, M.; Waldera-Lupa, D.M.; Kögler, G.; Meyer, H.E.; Müller, H.W.; Stühler, K. Characterization of Regenerative Phenotype of Unrestricted Somatic Stem Cells (USSC) from Human Umbilical Cord Blood (hUCB) by Functional Secretome Analysis. Mol. Cell Proteom. 2015, 14, 2630–2643. [Google Scholar] [CrossRef]
- Schira, J.; Heinen, A.; Poschmann, G.; Ziegler, B.; Hartung, H.P.; Stühler, K.; Küry, P. Secretome analysis of nerve repair mediating Schwann cells reveals Smad-dependent trophism. FASEB J. 2019, 33, 4703–4715. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Michalski, A.; Mann, M. Software lock mass by two-dimensional minimization of peptide mass errors. J. Am. Soc. Mass Spectrom. 2011, 22, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Poschmann, G.; Waldera-Lupa, D.; Rafiee, N.; Kollmann, M.; Stuhler, K. OutCyte: A novel tool for predicting unconventional protein secretion. Sci. Rep. 2019, 9, 19448. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Oligodendroglial Lineage-Promoting Factor | Gene Name | Description | Secretion Via/Localization | Reference |
|---|---|---|---|---|---|
| P07897 | Aggrecan core protein | Acn | Inhibits oligodendrocyte myelination | SP 1 | [30] |
| A0A140TA94 | Chordin | Chrd | Promotes oligodendrogenesis From subventricular zone derived stem cells in vitro and in vivo | SP | [31] |
| Q9R1E9 | Connective tissue growth factor | Ctgf | Negatively regulates myelination | SP | [32] |
| F1M8K0 | Dystroglycan 1 | Dag1 | Intracellular portion of cleaved dag promotes oligodendroglial precursor cell proliferation/regulates myelin membrane production, growth, or stability | SP | [33,34] |
| A0A096P6L8 | Fibronectin | Fn | Maintenance and proliferation of OPCs | SP | [35] |
| Q63772 | Growth arrest-specific protein 6 | Gas6 | Protects oligodendrocytes from tumor necrosis factor-α-induced apoptosis | SP | [36] |
| P12843 | Insulin-like growth factor-binding protein 2 | Igfbp2 | Negative effector of oligodendrocyte survival and differentiation | SP | [37] |
| F1M9B2 | Insulin-like growth factor-binding protein 7 | Igfbp7 | Inhibits the differentiation of oligodendrocyte precursor cells via regulation of wingless-related integration site (Wnt) / β-catenin signaling | SP | [38] |
| P12843 | Laminin subunit beta-2 | Lamb2 | Extracellular matrix proteins known to be important for myelination | SP | [39,40] |
| F1M798 | Metalloendopeptidase | BMP1 | Inhibits oligodendrogenesis | SP | [41] |
| P30120 | Tissue inhibitor of metalloproteinase type 1 | Timp-1 | Promotes oligodendrocyte differentiation and central nervous system myelination | SP | [42,43] |
| P08721 | Osteopontin | Spp1 | Stimulates MBP synthesis and myelin sheath formation in vitro | SP | [44] |
| F7EPE0 | Sulfated glycoprotein 1/prosaposin | Psap | Known as a myelinotrophic factor protecting myelinating glial cells | SP | [45,46] |
| Q9JI92 | Syntenin-1 | Sdcbp | Plays a role in OPC migration | UPS 2 | [47] |
| M0R979 | Thrombospondin 1 | Thbs1 | Reduces oligodendrogenesis | SP | [48] |
| A0A0G2K1L0 | Tenascin c | Tnc | Inhibits OPC differentiation and myelination | SP | [49] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samper Agrelo, I.; Schira-Heinen, J.; Beyer, F.; Groh, J.; Bütermann, C.; Estrada, V.; Poschmann, G.; Bribian, A.; Jadasz, J.J.; Lopez-Mascaraque, L.; et al. Secretome Analysis of Mesenchymal Stem Cell Factors Fostering Oligodendroglial Differentiation of Neural Stem Cells In Vivo. Int. J. Mol. Sci. 2020, 21, 4350. https://doi.org/10.3390/ijms21124350
Samper Agrelo I, Schira-Heinen J, Beyer F, Groh J, Bütermann C, Estrada V, Poschmann G, Bribian A, Jadasz JJ, Lopez-Mascaraque L, et al. Secretome Analysis of Mesenchymal Stem Cell Factors Fostering Oligodendroglial Differentiation of Neural Stem Cells In Vivo. International Journal of Molecular Sciences. 2020; 21(12):4350. https://doi.org/10.3390/ijms21124350
Chicago/Turabian StyleSamper Agrelo, Iria, Jessica Schira-Heinen, Felix Beyer, Janos Groh, Christine Bütermann, Veronica Estrada, Gereon Poschmann, Ana Bribian, Janusz J. Jadasz, Laura Lopez-Mascaraque, and et al. 2020. "Secretome Analysis of Mesenchymal Stem Cell Factors Fostering Oligodendroglial Differentiation of Neural Stem Cells In Vivo" International Journal of Molecular Sciences 21, no. 12: 4350. https://doi.org/10.3390/ijms21124350
APA StyleSamper Agrelo, I., Schira-Heinen, J., Beyer, F., Groh, J., Bütermann, C., Estrada, V., Poschmann, G., Bribian, A., Jadasz, J. J., Lopez-Mascaraque, L., Kremer, D., Martini, R., Müller, H. W., Hartung, H. P., Adjaye, J., Stühler, K., & Küry, P. (2020). Secretome Analysis of Mesenchymal Stem Cell Factors Fostering Oligodendroglial Differentiation of Neural Stem Cells In Vivo. International Journal of Molecular Sciences, 21(12), 4350. https://doi.org/10.3390/ijms21124350