A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research

 , ,
, ,
Abstract
1. Introduction
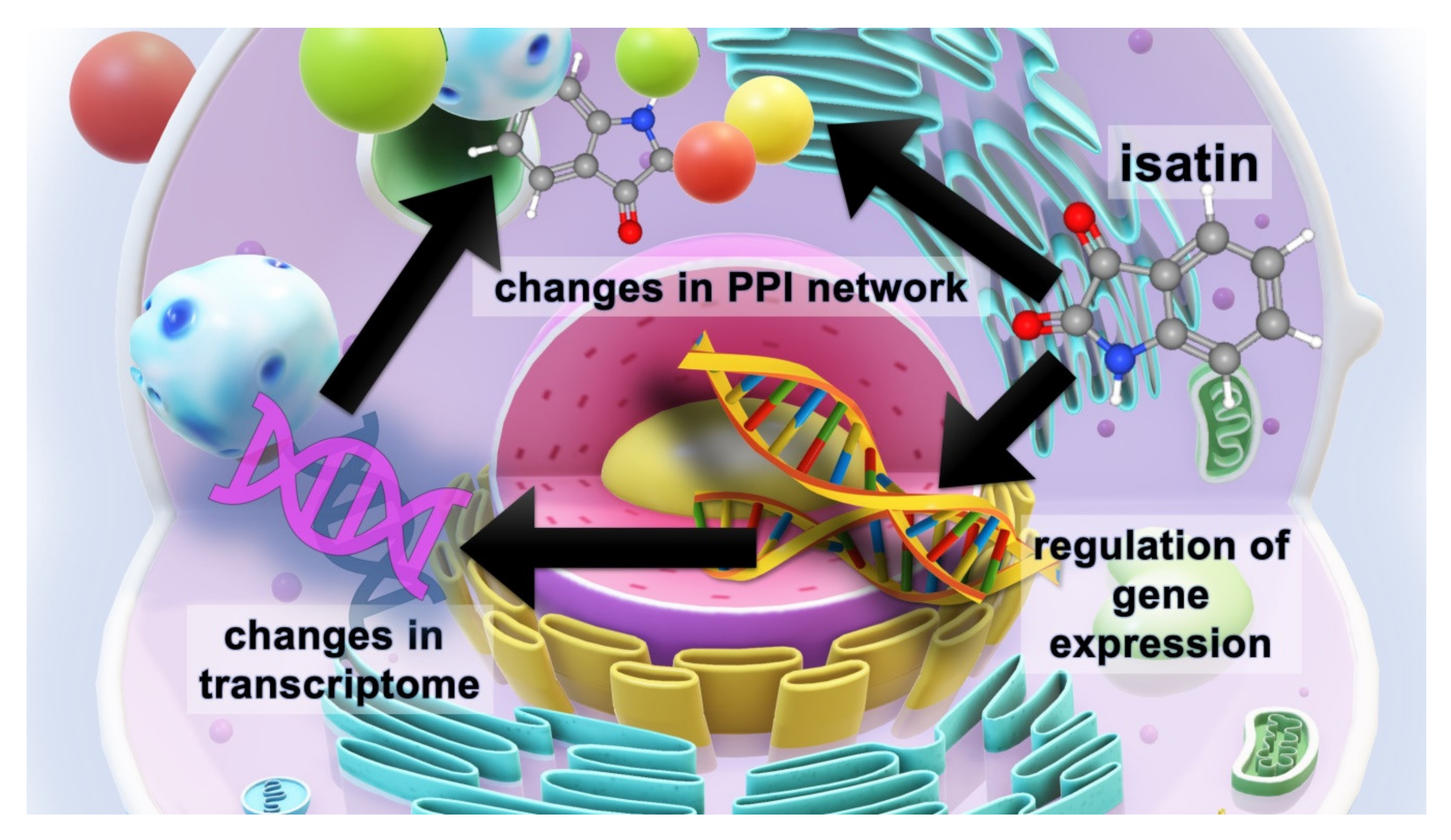
2. Results
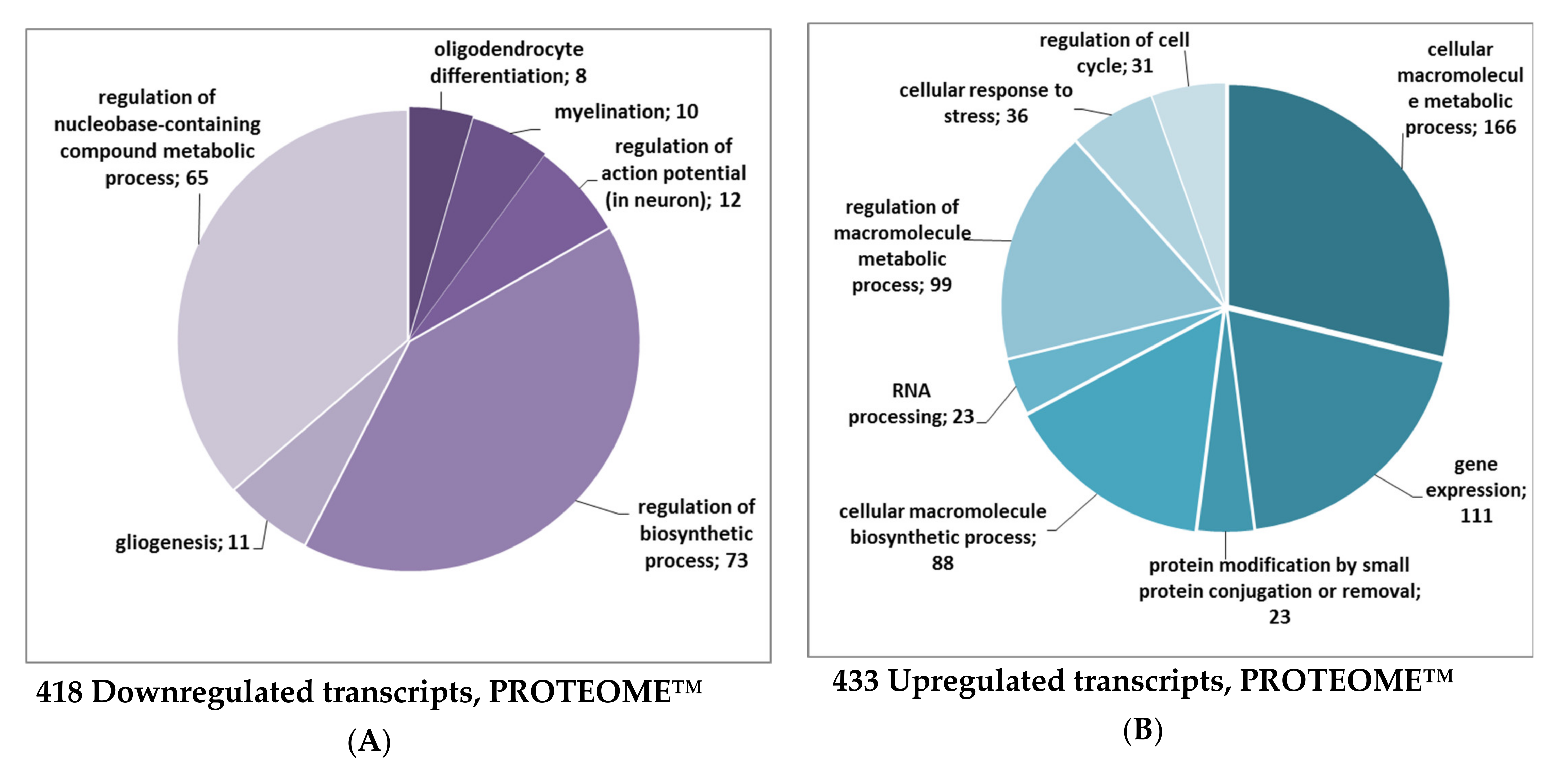
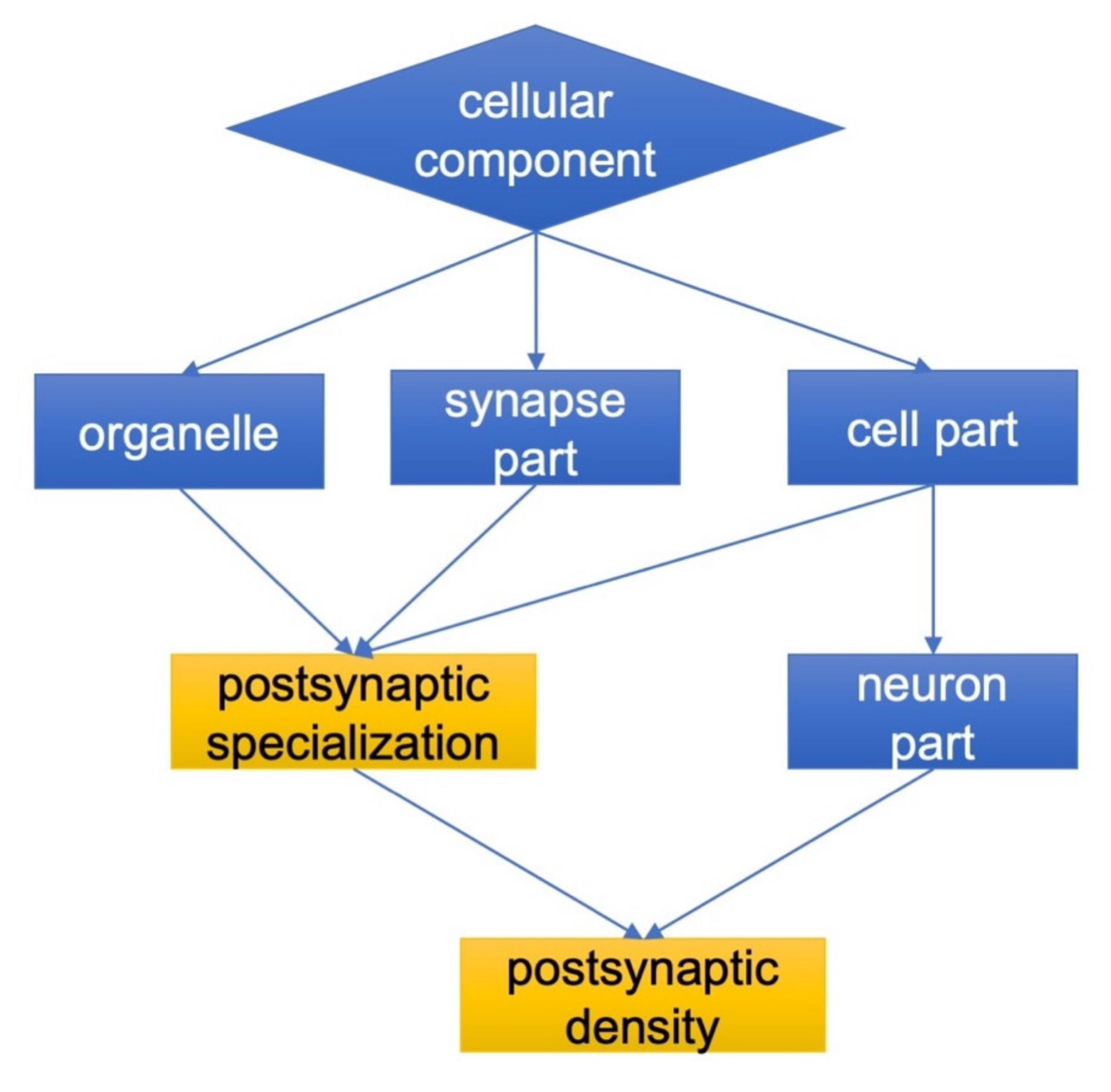
2.1. The Effect of Isatin Administration to Mice on Differently Expressed Proteins in Brain Hemispheres
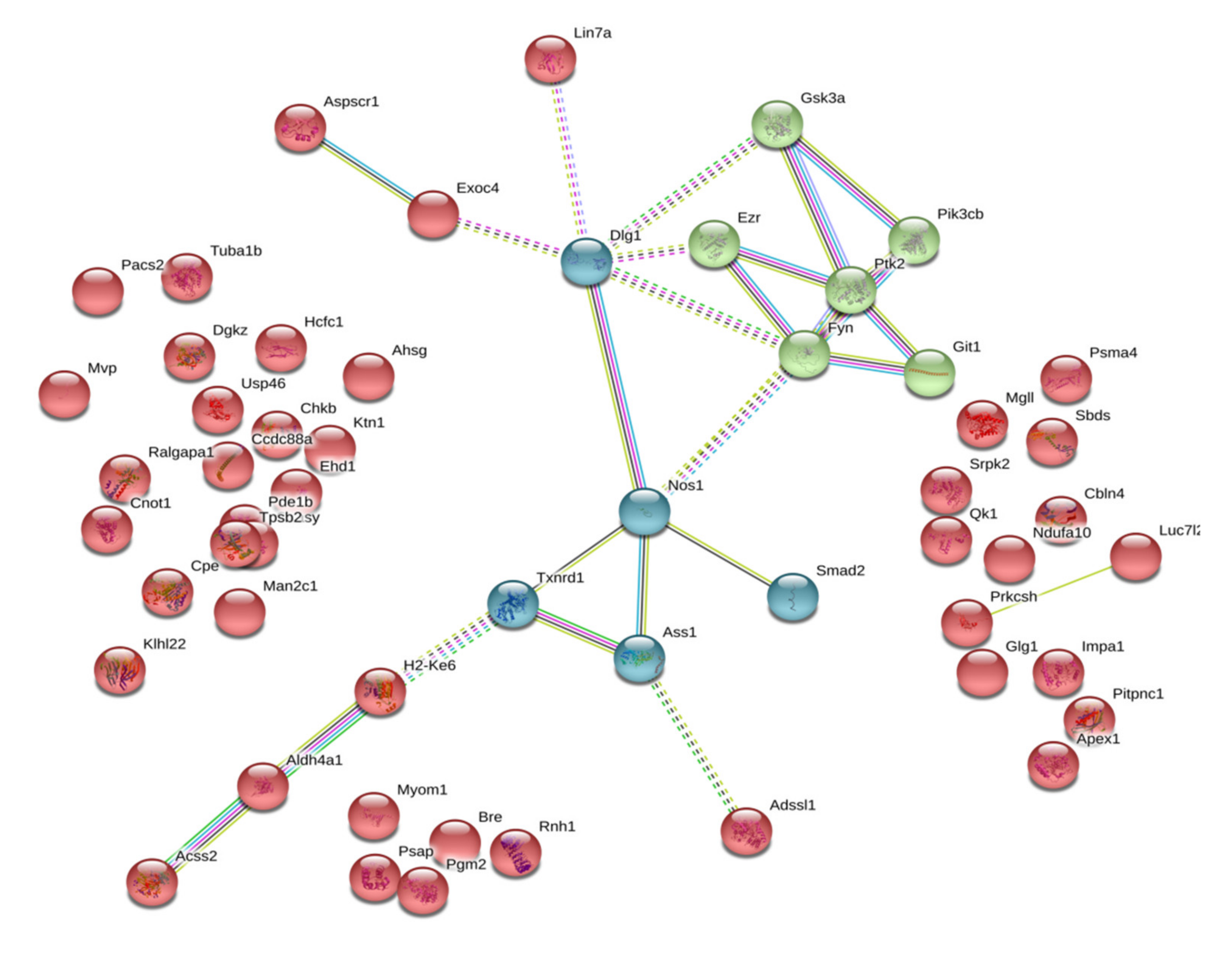
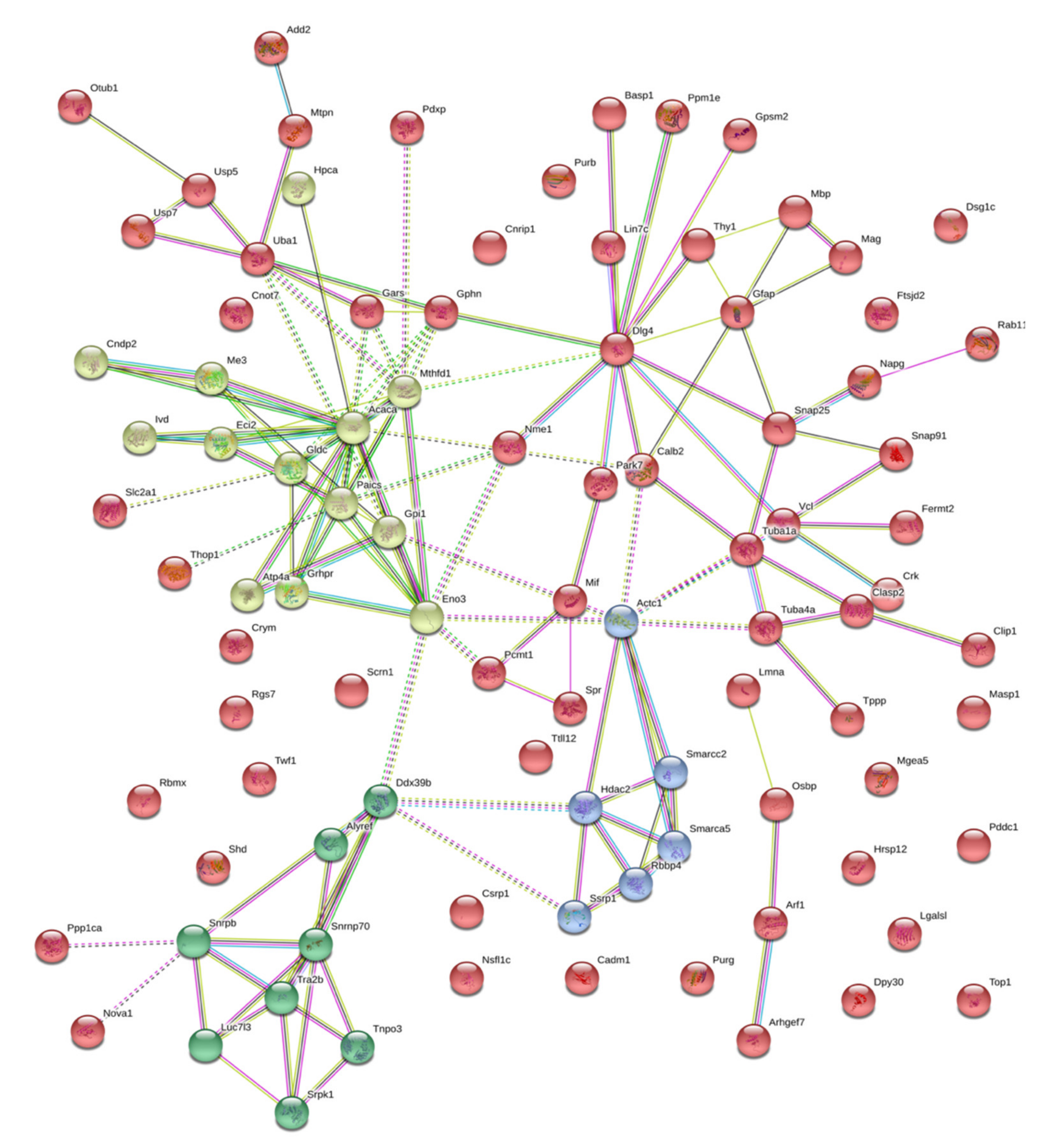
2.2. Isatin-Binding Proteins
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Isatin Administration
4.3. Sample Preparation for Transcriptome Analysis and Proteomic Profiling of Isatin-Binding Proteins
4.3.1. Transcriptome Analysis
4.3.2. Functional Analysis of Differentially Expressed Genes
4.3.3. Affinity Chromatography and Sample Preparation for Mass Spectrometric Analysis
4.3.4. Mass Spectrometric Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Medvedev, A.; Buneeva, O.; Gnedenko, O.; Ershov, P.; Ivanov, A. Isatin, an endogenous non-peptide biofactor: A review of its molecular targets, mechanisms of actions and their biomedical implications. Biofactors 2018, 44, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Phogat, P.; Singh, P. A mini review on central nervous system potential of isatin derivatives. Cent. Nerv. Syst. Agents Med. Chem. 2015, 15, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.; Buneeva, O.; Glover, V. Biological targets for isatin and its analogues: Implications for therapy. Biologics 2007, 1, 151–162. [Google Scholar] [PubMed]
- Medvedev, A.; Igosheva, N.; Crumeyrolle-Arias, M.; Glover, V. Isatin: Role in stress and anxiety. Stress 2005, 8, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.E.; Clow, A.; Sandler, M.; Glover, V. Isatin—A link between natriuretic peptides and monoamines? Biochem. Pharmacol. 1996, 52, 385–391. [Google Scholar] [CrossRef]
- Chatterjee, S.; Lee, L.Y.; Kawahara, R.; Abrahams, J.L.; Adamczyk, M.A.; Anugraham, M.; Ashwood, C.; Sumer-Bayraktar, Z.; Briggs, M.T.; Chik, J.H.L.; et al. Protein Paucimannosylation Is an Enriched N ‐Glycosylation Signature of Human Cancers. Proteomics 2010, 19. [Google Scholar] [CrossRef]
- Pandeya, S.N.; Smitha, S.; Jyoti, M.; Sridhar, S.K. Biological activities of isatin and its derivatives. Acta Pharm. 2005, 55, 27–46. [Google Scholar]
- Vine, K.L.; Matesic, L.; Locke, J.M.; Ranson, M.; Skropeta, D. Cytotoxic and anticancer activities of isatin and its derivatives: A comprehensive review from 2000–2008. Anticancer Agents Med. Chem. 2009, 9, 397–414. [Google Scholar] [CrossRef]
- Pakravan, P.; Kashanian, S.; Khodaei, M.M.; Harding, F.J. Biochemical and pharmacological characterization of isatin and its derivatives: From structure to activity. Pharmacol. Rep. 2013, 65, 313–335. [Google Scholar] [CrossRef]
- Verma, M.; Pandeya, S.N.; Singh, K.N.; Stables, J.P. Anticonvulsant activity of Schiff bases of isatin derivatives. Acta Pharm. 2004, 54, 49–56. [Google Scholar]
- Chohan, Z.H.; Pervez, H.; Rauf, A.; Khan, K.M.; Supuran, C.T. Isatin-derived antibacterial and antifungal compounds and their transition metal complexes. J. Enzyme Inhib. Med. Chem. 2004, 19, 417–423. [Google Scholar] [CrossRef]
- Sriram, D.; Bal, T.R.; Yogeeswari, P. Design, synthesis and biological evaluation of novel non-nucleoside HIV-1 reverse transcriptase inhibitors with broad-spectrum chemotherapeutic properties. Bioorg. Med. Chem. 2004, 12, 5865–5873. [Google Scholar] [CrossRef] [PubMed]
- Mak, N.K.; Leung, C.Y.; Wei, X.Y.; Shen, X.L.; Wong, R.N.; Leung, K.N.; Fung, M.C. Inhibition of RANTES expression by indirubin in influenza virus-infected human bronchial epithelial cells. Biochem. Pharmacol. 2004, 67, 167–174. [Google Scholar] [CrossRef]
- Selvam, P.; Murgesh, N.; Chandramohan, M.; De Clercq, E.; Keyaerts, E.; Vijgen, L.; Maes, P.; Neyts, J.; Ranst, M.V. In vitro antiviral activity of some novel isatin derivatives against HCV and SARS-CoV viruses. Indian J. Pharm. Sci. 2008, 70, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.G.; Magee, W.P.; Stukenbrok, H.A.; Beckius, G.E.; Milici, A.J.; Tracey, W.R. A novel nonpeptidic caspase-3/7 inhibitor, (S)-(+)-5-[1-(2-methoxymethylpyrrolidinyl)sulfonyl]isatin reduces myocardial ischemic injury. Eur. J. Pharmacol. 2002, 456, 59–68. [Google Scholar] [CrossRef]
- Limpachayaporn, P.; Schäfers, M.; Haufe, G. Isatin sulfonamides: Potent caspases-3 and -7 inhibitors, and promising pet and spect radiotracers for apoptosis imaging. Future Med. Chem. 2015, 7, 1173–1196. [Google Scholar] [CrossRef] [PubMed]
- Motzer, R.J.; Escudier, B.; Gannon, A.; Figlin, R.A. Sunitinib: Ten Years of Successful Clinical Use and Study in Advanced Renal Cell Carcinoma. Oncologist 2017, 22, 41–52. [Google Scholar] [CrossRef]
- Watkins, P.; Clow, A.; Glover, V.; Halket, J.; Przyborowska, A.; Sandler, M. Isatin, regional distribution in rat brain and tissues. Neurochem. Int. 1990, 17, 321–323. [Google Scholar] [CrossRef]
- Manabe, S.; Gao, Q.; Yuan, J.; Takahashi, T.; Ueki, A. Determination of isatin in urine and plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1997, 691, 197–202. [Google Scholar] [CrossRef]
- Mawatari, K.; Segawa, M.; Masatsuka, R.; Hanawa, Y.; Iinuma, F.; Watanabe, M. Fluorimetric determination of isatin in human urine and serum by liquid chromatography postcolumn photoirradiation. Analyst 2001, 126, 33–36. [Google Scholar] [CrossRef]
- Igosheva, N.; Matta, S.; Glover, V. Effect of acute stress and gender on isatin in rat tissues and serum. Physiol. Behav. 2004, 80, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Tozawa, Y.; Ueki, A.; Manabe, S.; Matsushima, K. Stress-induced increase in urinary isatin excretion in rats: Reversal by both dexamethasone and alpha-methyl-P-tyrosine. Biochem. Pharmacol. 1998, 56, 1041–1046. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Clow, A.; Przyborowska, A.; Halket, J.; Glover, V.; Sandler, M. Effect of aromatic amino acids, pentylenetetrazole and yohimbine on isatin and tribulin activity in rat brain. Neurosci. Lett. 1991, 132, 44–46. [Google Scholar] [CrossRef]
- Sandler, M.; Przyborowska, A.; Halket, J.; Watkins, P.; Glover, V.; Coates, M.E. Urinary but not brain isatin levels are reduced in germ-free rats. J. Neurochem. 1991, 57, 1074–1075. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Mitra, S.K.; Acharya, S.B. Anxiogenic activity of isatin, a putative biological factor, in rodents. J. Psychopharmacol. 1991, 5, 202–206. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Acharya, S.B. Further investigations on the anxiogenic effects of isatin. Biog. Amines 1993, 9, 453–463. [Google Scholar]
- Abel, E.L. Behavioral effects of isatin on open field activity and immobility in the forced swim test in rats. Physiol. Behav. 1995, 57, 611–613. [Google Scholar] [CrossRef]
- Chocholova, L.; Kolinova, M. Effect of isatin (2,3-Dioxoindoline) on audiogenic seizures in rats and its relationship to electragraphic and behavioural phenomena. Physiol. Bohemoslov. 1979, 28, 495–502. [Google Scholar]
- Medvedev, A.E.; Gorkin, V.Z.; Fedotova, I.B.; Semiokhina, A.F.; Sandler, M.; Glover, V. Antiseizure effect of isatin and reduction of monoamine oxidase activity in rats with experimental audiogenic seizures. Med. Sci. Res. 1994, 22, 555–557. [Google Scholar]
- Bhattacharya, S.K.; Charkrabarti, A. Dose-related proconvulsant and anticonvulsant activity of isatin, a putative biological factor in rats. Ind. J. Exp. Biol. 1998, 36, 118–121. [Google Scholar]
- Zhou, Y.; Zhao, Z.Q.; Xie, J.X. Effect of isatin on rotational behavior and DA levels in caudate putamen in Parkinsonian rats. Brain Res. 2001, 917, 127–132. [Google Scholar] [CrossRef]
- Hamaue, N.; Minami, M.; Terado, M.; Hirafuji, M.; Endo, T.; Machida, M.; Hiroshige, T.; Ogata, A.; Tashiro, K.; Saito, H.; et al. Comparative study of the effects of isatin, an endogenous MAO-inhibitor, and selegiline on bradykinesia and dopamine levels in a rat model of Parkinson’s disease induced by the Japanese encephalitis virus. Neurotoxicology 2004, 25, 205–213. [Google Scholar] [CrossRef]
- Minami, M.; Hamaue, N.; Hirafuji, M.; Saito, H.; Hiroshige, T.; Ogata, A.; Tashiro, K.; Parvez, S.H. Isatin, an endogenous MAO inhibitor, and a rat model of Parkinson’s disease induced by the Japanese encephalitis virus. J. Neural Transm. 2006, 71, 87–95. [Google Scholar]
- Medvedev, A.E.; Buneeva, O.A.; Kopylov, A.T.; Tikhonova, O.V.; Medvedeva, M.V.; Nerobkova, L.N.; Kapitsa, I.G.; Zgoda, V.G. Brain mitochondrial subproteome of Rpn10-binding proteins and its changes induced by the neurotoxin MPTP and the neuroprotector isatin. Biochemistry 2017, 82, 330–339. [Google Scholar] [CrossRef]
- Buneeva, O.; Kopylov, A.; Kapitsa, I.; Ivanova, E.; Zgoda, V.; Medvedev, A. The effect of neurotoxin MPTP and neuroprotector isatin on the profile of ubiquitinated brain mitochondrial proteins. Cells 2018, 7, 91. [Google Scholar] [CrossRef]
- Crumeyrolle-Arias, M.; Buneeva, O.; Zgoda, V.; Kopylov, A.; Cardona, A.; Tournaire, M.-C.; Pozdnev, V.; Glover, V.; Medvedev, A. Isatin binding proteins in rat brain: In situ imaging, quantitative characterization of specific [3H]isatin binding, and proteomic profiling. J. Neurosci. Res. 2009, 87, 2763–2772. [Google Scholar] [CrossRef]
- Buneeva, O.; Gnedenko, O.; Zgoda, V.; Kopylov, A.; Glover, V.; Ivanov, A.; Medvedev, A.; Archakov, A. Isatin binding proteins of rat and mouse brain: Proteomic identification and optical biosensor validation. Proteomics 2010, 10, 23–37. [Google Scholar] [CrossRef]
- Buneeva, O.A.; Kopylov, A.T.; Tikhonova, O.V.; Zgoda, V.G.; Medvedev, A.E.; Archakov, A.I. Effect of affinity sorbent on proteomic profiling of isatin-binding proteins of mouse brain. Biochemistry 2012, 77, 1584–1599. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Buneeva, O.A.; Kopylov, A.T.; Gnedenko, O.V.; Medvedeva, M.V.; Kozin, S.A.; Ivanov, A.S.; Zgoda, V.G.; Makarov, A.A. The effects of an endogenous non-peptide molecule isatin and hydrogen peroxide on proteomic profiling of rat brain amyloid-beta binding proteins: Relevance to Alzheimer’s disease? Int. J. Mol. Sci. 2015, 16, 476–495. [Google Scholar] [CrossRef]
- Medvedev, A.; Buneeva, O.; Gnedenko, O.; Fedchenko, V.; Medvedeva, M.; Ivanov, Y.; Glover, V.; Sandler, M. Isatin interaction with glyceraldehyde-3-phosphate dehydrogenase, a putative target of neuroprotective drugs: Partial agonism with deprenyl. J. Neural Transm. 2006, 71, 195–203. [Google Scholar]
- Cane, A.; Tournaire, M.C.; Barritault, D.; Crumeyrolle-Arias, M. The endogenous oxindoles 5-hydroxyoxindole and isatin are antiproliferative and proapoptotic. Biochem. Biophys. Res. Commun. 2000, 276, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Igosheva, N.; Lorz, C.; O’Conner, E.; Glover, V.; Mehmet, H. Isatin, an endogenous monoamine oxidase inhibitor, triggers a dose- and time-dependent switch from apoptosis to necrosis in human neuroblastoma cells. Neurochem. Int. 2005, 47, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Ju, C.; Zhang, J.; Song, J.; Ge, Y.; Yue, W. Antitumor effects of isatin on human neuroblastoma cell line (SH-SY5Y) and the related mechanism. Eur. J. Pharmacol. 2008, 589, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Hou, L.; Ju, C.; Zhang, J.; Ge, Y.; Yue, W. Isatin inhibits proliferation and induces apoptosis of SH-SY5Y neuroblastoma cells in vitro and in vivo. Eur. J. Pharmacol. 2013, 702, 235–241. [Google Scholar] [CrossRef]
- Ma, Z.; Hou, L.; Jiang, Y.; Chen, Y.; Song, J. The endogenous oxindole isatin induces apoptosis of MCF 7 breast cancer cells through a mitochondrial pathway. Oncol. Rep. 2014, 32, 2111–2117. [Google Scholar] [CrossRef][Green Version]
- Sun, W.; Zhang, L.; Hou, L.; Ju, C.; Zhao, S.; Wei, Y. Isatin inhibits SH-SY5Y neuroblastoma cell invasion and metastasis through MAO/HIF-1α/CXCR4 signaling. Anticancer Drugs 2017, 28, 645–653. [Google Scholar] [CrossRef]
- Fogaça, M.V.; Cândido-Bacani, P.M.; Benicio, L.M.; Zapata, L.M.; Cardoso, P.F.; de Oliveira, M.T.; Calvo, T.R.; Varanda, E.A.; Vilegas, W.; de Syllos Colus, I.M. Effects of indirubin and isatin on cell viability, mutagenicity, genotoxicity and BAX/ERCC1 gene expression. Pharm. Biol. 2017, 55, 2005–2014. [Google Scholar] [CrossRef]
- Buneeva, O.A.; Kapitsa, I.G.; Ivanova, E.A.; Kopylov, A.T.; Zgoda, V.G.; Medvedev, A.E. The effect of a neuroprotective dose of isatin or deprenyl to mice on the profile of brain isatin-binding proteins. Biomed. Khim. 2019, 65, 407–417. [Google Scholar] [CrossRef]
- Panova, N.G.; Zemskova, M.A.; Axenova, L.N.; Medvedev, A.E. Does isatin interact with rat brain monoamine oxidases in vivo? Neurosci. Lett. 1997, 233, 58–60. [Google Scholar] [CrossRef]
- Xu, P.; Hou, L.; Ju, C.; Zhang, Z.; Sun, W.; Zhang, L.; Song, J.; Lv, Y.; Liu, L.; Chen, Z.; et al. Isatin inhibits the proliferation and invasion of SHSY5Y neuroblastoma cells. Mol. Med. Rep. 2016, 13, 2757–2762. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, W.; Cao, Y.; Hou, L.; Ju, C.; Wang, X. Isatin inhibits the invasion of human SHSY5Y neuroblastoma cells based on microarray analysis. Mol. Med. Rep. 2019, 20, 1700–1706. [Google Scholar] [PubMed]
- Fedchenko, V.; Globa, A.; Kaloshin, A.; Kapitsa, I.; Nerobkova, L.; Val’dman, E.; Buneeva, O.; Glover, V.; Medvedev, A. The effect of short-term administration of (-)-deprenyl and isatin on expression of some genes in the mouse brain cortex. Med. Sci. Monit. 2008, 14, BR269–BR273. [Google Scholar] [PubMed]
- Rehni, A.K.; Singh, T.G.; Jaggi, A.S.; Singh, N. Pharmacological preconditioning of the brain: A possible interplay between opioid and calcitonin gene related peptide transduction systems. Pharmacol. Rep. 2008, 60, 904–913. [Google Scholar]
- Snaebjornsson, M.T.; Schulze, A. Non-canonical functions of enzymes facilitate cross-talk between cell metabolic and regulatory pathways. Exp. Mol. Med. 2018, 50, 34. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Walch, L. Emerging Role of the Scaffolding Protein Dlg1 in Vesicle Trafficking. Traffic 2013, 14, 964–973. [Google Scholar] [CrossRef]
- Von Ossowski, L.; Li, L.-L.; MoÈykkynen, T.; Coleman, S.K.; Courtney, M.J.; KeinaÈnen, K. Cysteine 893 is a target of regulatory thiol modifications of GluA1 AMPA receptors. PLoS ONE 2017, 12, e0171489. [Google Scholar] [CrossRef]
- Saura, M.; Zaragoza, C.; Herranz, B.; Griera, M.; Diez-Marqués, L.; Rodriguez-Puyol, D.; Rodriguez-Puyol, M. Nitric oxide regulates transforming growth factor-beta signaling in endothelial cells. Circ. Res. 2005, 97, 1115–11123. [Google Scholar] [CrossRef]
- Denis, J.-F.; Sader, F.; Gatien, S.; Villiard, É.; Philip, A.; Roy, S. Activation of Smad2 but not Smad3 is required to mediate TGF-β signaling during axolotl limb regeneration. Development 2016, 143, 3481–3490. [Google Scholar] [CrossRef]
- Viswanatha, R.; Wayt, J.; Ohouo, P.Y.; Smolka, M.B.; Bretscher, A. Interactome analysis reveals ezrin can adopt multiple conformational states. J. Biol. Chem. 2013, 288, 35437–35451. [Google Scholar] [CrossRef]
- McClatchey, A.I. ERM proteins at a glance. J. Cell Sci. 2014, 127, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Poullet, P.; Gautreau, A.; Kadare, G.; Girault, J.-A.; Louvard, D.; Arpin, M. Ezrin interacts with focal adhesion kinase and induces its activation independently of cell-matrix adhesion. J. Biol. Chem. 2001, 276, 37686–37691. [Google Scholar] [CrossRef] [PubMed]
- Tapial Martínez, P.; López Navajas, P.; Lietha, D. FAK structure and regulation by membrane interactions and force in focal adhesions. Biomolecules 2020, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Baillat, G.; Siret, C.; Delamarre, E.; Luis, J. Early adhesion induces interaction of FAK and Fyn in lipid domains and activates raft-dependent Akt signaling in SW480 colon cancer cells. Biochim. Biophys. Acta 2008, 1783, 2323–2331. [Google Scholar] [CrossRef][Green Version]
- Liu, S.; Premont, R.T.; Rockey, D. G-protein-coupled receptor kinase interactor-1 (GIT1) is a new endothelial nitric-oxide synthase (eNOS) interactor with functional effects on vascular homeostasis. J. Biol. Chem. 2012, 287, 12309–12320. [Google Scholar] [CrossRef]
- Zhang, H.; Webb, D.J.; Asmussen, H.; Horwitz, A.F. Synapse formation is regulated by the signaling adaptor GIT1. J. Cell Biol. 2003, 161, 131–142. [Google Scholar] [CrossRef]
- Beurel, E.; Grieco, S.F.; Jope, R.S. Glycogen synthase kinase-3 (GSK3): Regulation, actions, and diseases. Pharmacol. Ther. 2015, 148, 114–131. [Google Scholar] [CrossRef]
- Pilot-Storck, F.; Chopin, E.; Rual, J.-F.; Baudot, A.; Dobrokhotov, P.; Robinson-Rechavi, M.; Brun, C.; Cusick, M.E.; Hill, D.E.; Schaeffer, L.; et al. Interactome mapping of the phosphatidylinositol 3-kinase-mammalian target of rapamycin pathway identifies deformed epidermal autoregulatory factor-1 as a new glycogen synthase kinase-3 interactor. Mol. Cell. Proteom. 2010, 9, 1578–1593. [Google Scholar] [CrossRef]
- Popkie, A.P.; Zeidner, L.C.; Albrecht, A.M.; D’Ippolito, A.; Eckardt, S.; Newsom, D.E.; Groden, J.; Doble, B.W.; Aronow, B.; McLaughlin, K.J.; et al. Phosphatidylinositol 3-kinase (PI3K) signaling via glycogen synthase kinase-3 (GSK-3) regulates DNA methylation of imprinted loci. J. Biol. Chem. 2010, 285, 41337–41347. [Google Scholar] [CrossRef]
- Karamanou, S.; Bariami, V.; Papanikou, E.; Kalodimos, C.G.; Economou, A. Assembly of the translocase motor onto the preprotein-conducting channel. Mol. Microbiol. 2008, 70, 311–322. [Google Scholar] [CrossRef]
- Rondeau, P.; Bourdon, E. The glycation of albumin: Structural and functional impacts. Biochimie 2011, 93, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Stoll, P.; Bassler, N.; Hagemeyer, C.E.; Eisenhardt, S.U.; Chen, Y.C.; Schmidt, R.; Schwarz, M.; Ahrens, I.; Katagiri, Y.; Pannen, B.; et al. Targeting ligand-induced binding sites on GPIIb/IIIa via single-chain antibody allows effective anticoagulation without bleeding time prolongation. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1206–1212. [Google Scholar] [CrossRef]
- Ivanov, A.; Medvedev, A.; Ershov, P.; Molnar, A.; Mezentsev, Y.; Yablokov, E.; Kaluzhsky, L.; Gnedenko, O.; Buneeva, O.; Haidukevich, I.; et al. Protein interactomics based on direct molecular fishing on paramagnetic particles: Practical realization and further SPR validation. Proteomics 2014, 14, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Ershov, P.; Mezentsev, Y.; Gilep, A.; Usanov, S.; Buneeva, O.; Medvedev, A.; Ivanov, A. Isatin-induced increase in the affinity of human ferrochelatase and adrenodoxin reductase interaction. Protein Sci. 2017, 26, 2458–2462. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Ennajdaoui, H.; Edmondson, C.; Wirth, B.; Sanford, J.; Chen, B. Splicing factor TRA2B is required for neural progenitor survival. J. Comp. Neurol. 2014, 522, 372–392. [Google Scholar] [CrossRef]
- Liu, J.; Bian, T.; Feng, J.; Qian, L.; Zhang, J.; Jiang, D.; Zhang, Q.; Li, X.; Liu, Y.; Shi, J. miR-335 inhibited cell proliferation of lung cancer cells by target Tra2b. Cancer Sci. 2018, 109, 289–296. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, K.I.; Parisien, M.; Dai, Q.; Diatchenko, L.; Pan, T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017, 45, 6051–6063. [Google Scholar] [CrossRef]
- Correa, B.R.; de Araujo, P.R.; Qiao, M.; Burns, S.C.; Chen, C.; Schlegel, R.; Agarwal, S.; Galante, P.A.F.; Penalva, L.O.F. Functional genomics analyses of RNA-binding proteins reveal the splicing regulator SNRPB as an oncogenic candidate in glioblastoma. Genome Biol. 2016, 17, 125. [Google Scholar] [CrossRef]
- Motley, W.W.; Talbot, K.; Fischbeck, K.H. GARS axonopathy: Not every neuron’s cup of tRNA. Trends Neurosci. 2010, 33, 59–66. [Google Scholar] [CrossRef]
- Yao, P.; Fox, P.L. 2013 Aminoacyl-tRNA synthetases in medicine and disease. EMBO Mol. Med. 2013, 5, 332–343. [Google Scholar] [CrossRef]
- Hou, Y.; Shang, C.; Wang, H.; Yun, J. Isatin-azole hybrids and their anticancer activities. Arch. Pharm. 2020, 353, e1900272. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Tulsyan, S.; Bharadwaj, M.; Mehrota, R. Systematic review on cytotoxic and anticancer potential of n-substituted isatins as novel class of compounds useful in multidrug-resistant cancer therapy: In silico and in vitro analysis. Top. Curr. Chem. 2019, 377, 15. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, M.; Eldehna, W.M.; Almahli, H.; Elgez, A.; Fares, M.; Elaasser, M.M.; Abdel-Aziz, H.A. Novel thiazolidinone/thiazolo[3,2-a]benzimidazolone-isatin conjugates as apoptotic anti-proliferative agents towards breast cancer: One-pot synthesis and in vitro biological evaluation. Molecules 2018, 23, 1420. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.E.; Goodwin, B.L.; Sandler, M.; Glover, V. Efficacy of isatin analogues as antagonists of rat brain and heart atrial natriuretic peptide receptors coupled to particulate guanylyl cyclase. Biochem. Pharmacol. 1999, 57, 913–915. [Google Scholar] [CrossRef]
- GeneXplain platform, release 3.0. Available online: http://platform.genexplain.com/ (accessed on 16 March 2020).
- Vaudel, M.; Barsnes, H.; Berven, F.S.; Sickmann, A.; Martens, L. SearchGUI: An open-source graphical user interface for simultaneous OMSSA and X!Tandem searches. Proteomics 2011, 11, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.E.; Gygi, S.P. Target-decoy search strategy for increased confidence in large-scale protein identifications by mass spectrometry. Nat. Methods 2007, 4, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Vaudel, M.; Burkhart, J.M.; Zahedi, R.P.; Oveland, E.; Berven, F.S.; Sickmann, A.; Martens, L.; Barsnes, H. PeptideShaker enables reanalysis of MS-derived proteomics data sets. Nat. Biotechnol. 2015, 33, 22–24. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, S.; Mathews, M.; Bereman, M.S.; Rubel, E.W.; MacCoss, M.J.; Noble, W.S. Estimating relative abundances of proteins from shotgun proteomics data. BMC Bioinform. 2012, 13, 308. [Google Scholar] [CrossRef]
- Glover, V.; Halket, J.M.; Watkins, P.J.; Clow, A.; Goodwin, B.L.; Sandler, M. Isatin: Identity with the purified endogenous monoamine oxidase inhibitor tribulin. J. Neurochem. 1988, 51, 656–659. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Entry Name | ANOVA (p) | Fold Change | Description | Function | |
|---|---|---|---|---|---|---|
| 1 | Q9JLV5 | CUL3 | 0 | −2.681 | Cullin-3 | E3 ubiquitin ligase of the ubiquitin-proteasome system |
| 2 | O35143 | ATIF1 | 1.44 × 10−15 | −2.655 | ATPase inhibitor, mitochondrial | Regulation of ATP synthesis |
| 3 | P34022 | RANG | 2.00 × 10−15 | −2.736 | Ran-specific GTPase-activating protein | Regulation of nucleocytoplasmic transport |
| 4 | Q921I1 | TRFE | 1.11 × 10−14 | −2.062 | Serotransferrin | Binding of iron ions and stimulation of cell proliferation |
| 5 | Q8C8N2 | SCAI | 9.71 × 10−14 | −2.521 | Protein SCAI | Tumor suppressor |
| 6 | Q9Z1R2 | BAG6 | 1.32 × 10−13 | −2.275 | Large proline-rich protein BAG6 | BAG family molecular chaperone regulator 6 is involved in DNA damage-induced apoptosis |
| 7 | Q61543 | GSLG1 | 9.94 × 10−12 | −2.164 | Golgi apparatus protein 1 | E-selectin ligand 1, cysteine-rich fibroblast growth factor receptor |
| 8 | Q9D2M8 | UB2V2 | 1.38 × 10−11 | −2.224 | Ubiquitin-conjugating enzyme E2 variant 2 | The component of the ubiquitin-proteasome system |
| 9 | P70670 | NACAM | 4.60 × 10−11 | −2.293 | Nascent polypeptide-associated complex subunit alpha, muscle-specific form | Molecular chaperone |
| 10 | Q8BGD9 | IF4B | 1.88 × 10−10 | −2.453 | Eukaryotic translation initiation factor 4B | Protein translation |
| 11 | Q00623 | APOA1 | 4.70 × 10−10 | −2.363 | Apolipoprotein A-I | In CNS, participates in reverse cholesterol transport |
| 12 | P84309 | ADCY5 | 6.87 × 10−10 | −2.72 | Adenylate cyclase type 5 | Cell signaling |
| 13 | Q99MN9 | PCCB | 7.83 × 10−10 | −2.249 | Propionyl-CoA carboxylase beta chain, mitochondrial | Fatty acid beta-oxidation |
| 14 | P07759 | SPA3K | 4.90 × 10−9 | −4.156 | Serine protease inhibitor A3K | A member of serine protease inhibitors, required for normal synaptic plasticity and regulation of serine protease mediated cell death |
| 15 | Q9EPN1 | NBEA | 5.62 × 10−9 | −3.393 | Neurobeachin | A protein required for central synapses formation and functioning |
| 16 | P22599 | A1AT2 | 5.78 × 10−9 | −2.595 | Alpha-1-antitrypsin 1-2 | A member of serine protease inhibitors, required for normal synaptic plasticity and regulation of serine protease-mediated cell death |
| 17 | Q62059 | CSPG2 | 2.58 × 10−8 | −2.126 | Versican core protein | A component of the extracellular matrix component of the brain |
| 18 | D3YXK2 | SAFB1 | 3.06 × 10−8 | −2.383 | Scaffold attachment factor B1 | Binds to specific sequences of DNA needed for genome organization into functional units in the cell nucleus |
| 19 | Q9D1X0 | NOL3 | 3.43 × 10−7 | −3.614 | Nucleolar protein 3 | An anti-apoptotic protein implicated in down-regulation of activities of caspases-2 and -8 and tumor protein p53 |
| 20 | P08414 | KCC4 | 7.85 × 10−7 | –10.97 | Calcium/calmodulin-dependent protein kinase type IV | Cell signaling |
| 21 | Q9D2N4 | DTNA | 1.05 × 10−6 | −2.133 | Dystrobrevin alpha | Required for stability of synapses and clustering of nicotinic acetylcholine receptors |
| 22 | P52760 | UK114 | 3.24 × 10−6 | −2.077 | Ribonuclease UK114 | Endoribonuclease causing translation inhibition by cleaving mRNA |
| 23 | Q922D8 | C1TC | 4.95 × 10−6 | −2.019 | C-1-tetrahydrofolate synthase, cytoplasmic | Crucial for de novo synthesis of purines |
| 24 | Q8C0E2 | VP26B | 9.11 × 10−6 | −2.027 | Vacuolar protein sorting-associated protein 26B | A component of the retromer cargo-selective complex (CSC) |
| 25 | Q6NZL0 | SOGA3 | 1.70 × 10−5 | −2.171 | Protein SOGA3 | A member of the SOGA (suppressor of glucose by autophagy) family involved in regulation of autophagy |
| 26 | Q61029 | LAP2B | 2.69 × 10−5 | −2.127 | Lamina-associated polypeptide 2, isoforms beta/delta/epsilon/gamma | Involved in maintenance of the structural organization of the nuclear envelope and DNA binding |
| 27 | O55091 | IMPCT | 2.85 × 10−5 | −2.608 | Protein IMPACT | Translational regulator that helps to maintain constant high levels of translation under stress conditions |
| 28 | P35235 | PTN11 | 2.89 × 10−5 | −2,467 | Tyrosine-protein phosphatase non-receptor type 11 | Participates in the signal transduction from the cell surface to the nucleus |
| 29 | P23492 | PNPH | 4.66 × 10−5 | −2.104 | Purine nucleoside phosphorylase | The enzyme involved in purine metabolism |
| 30 | Q9QXD6 | F16P1 | 1.31 × 10−4 | −8.295 | Fructose-1,6-bisphosphatase 1 | Glycolysis and non-canonical functions including blockade of the transcriptional activity of HIF-1α and activation of the RAS/RAF/MEK/ERK cascade |
| 31 | O88456 | CPNS1 | 6.14 × 10−3 | −2.523 | Calpain small subunit 1 | The regulatory subunit of the protease calpain, responsible for limited proteolysis of substrates involved in cytoskeleton remodeling and signal transduction |
| Gene Symbol | Gene Description | −Log (p value) | LogFold Change | Role/Effect |
|---|---|---|---|---|
| Upregulated genes | ||||
| Ctla2a | cytotoxic T lymphocyte-associated protein 2 alpha | 4.154 | 3.132 | Induces apoptosis in some cells |
| Nfkbia | alpha, nuclear factor of kappa light polypeptide gene enhancer in B cells inhibitor | 4.191 | 2.876 | Negative regulator of cell proliferation |
| Ctla2b | cytotoxic T lymphocyte-associated protein 2 beta | 4.376 | 2.735 | Induces apoptosis in some cells |
| Cdkn1a | cyclin-dependent kinase inhibitor 1A (P21) | 3.165 | 2.730 | Delays or stops the cell cycle |
| Errfi1 | ERBB receptor feedback inhibitor 1 | 4.676 | 2.236 | Negative regulator of several EGFR family members |
| Sgk1 | serum/glucocorticoid regulated kinase 1 | 3.642 | 1.940 | Regulation of vascular cell proliferation |
| Trp53inp1 | transformation related protein 53 inducible nuclear protein 1 | 3.301 | 1.887 | One of the key elements in p53-mediated cell cycle arrest and apoptosis in different cell types |
| Mtor | mechanistic target of rapamycin (serine/threonine kinase) | 4.091 | 1.787 | Involved in the regulation of cell growth, proliferation, autophagy, and protein synthesis |
| Cdt1 | chromatin licensing and DNA replication factor 1 | 4.554 | 1.747 | DNA replication |
| Xpo7 | exportin 7 | 4.626 | 1.722 | A member of the exportin family, responsible for nucleocytoplasmic trafficking of regulatory proteins |
| Gch1 | GTP cyclohydrolase 1 | 4.055 | 1.716 | Enzyme involved in tetrahydrobiopterin (BH4) biosynthesis; its expression is increased in pathological proliferation |
| Epc2 | enhancer of polycomb homolog 2 (Drosophila) | 3.578 | 1.691 | Epc plays an important role in cellular differentiation and development |
| Xpo4 | exportin 4 | 4.535 | 1.608 | A member of the exportin family, responsible for nucleocytoplasmic trafficking of regulatory proteins |
| Rbbp6 | retinoblastoma binding protein 6 | 4.721 | 1.606 | Acts as an E3 ubiquitin ligase; is a negative regulator of p53 and promotes cell proliferation |
| Ubqln1 | ubiquilin 1 | 3.187 | 1.596 | Involved in regulation of the protein quality control system |
| Recql4 | RecQ protein-like 4 | 3.030 | 1.575 | DNA helicase that belongs to the RecQ helicase family |
| Sap30 | sin3 associated polypeptide 30 kDa | 3.040 | 1.575 | Involved in the functional recruitment of the Sin3-histone deacetylase complex |
| Stac3 | SH3 and cysteine rich domain 3 | 4.708 | 1.573 | Involved in neuron specific signal transduction |
| Srsf6 | serine/arginine-rich splicing factor 6 | 3.077 | 1.457 | A member of the conserved family of RNA-binding proteins, which play an important role in the regulation of gene expression |
| Pnrc1 | proline-rich nuclear receptor coactivator 1 | 3.679 | 1.453 | Interacts with various nuclear receptors and functions as a tumor suppressor |
| Downregulated genes | ||||
| Cx3cr1 | chemokine (C-X3-C) receptor 1 | −3.709 | −2.796 | Has a significant impact on cell proliferation |
| Tiam2 | T cell lymphoma invasion and metastasis 2 | −5.154 | −2.105 | Plays an important role in neuron development and malignant proliferation in humans |
| Slc38a5 | member 5, solute carrier family 38 | −3.087 | −2.103 | Members of this superfamily of solute carriers are crucial for brain physiology and are implicated in various brain disorders |
| Lyl1 | lymphoblastomic leukemia 1 | −4.446 | −1.956 | Regulates expression of CREB1 target genes |
| Ier5l | immediate early response 5-like | −3.599 | −1.910 | Regulates the cellular response to mitogenic signals |
| Cyth4 | cytohesin 4 | −5.331 | −1.882 | This guanine nucleotide-exchange protein is an important regulator of signal transduction |
| Limd2 | LIM domain containing 2 | −3.070 | −1.867 | Is necessary for growth, development, and adaptive responses |
| Gpr17 | G protein-coupled receptor 17 | −3.345 | −1.853 | Cell signaling |
| Rasl10a | RAS-like, family 10, member A | −3.816 | −1.760 | A Ras-related protein that belongs to the superfamily of small GTPases and exhibits tumor suppressor potential. |
| Bcl11a | B cell CLL/lymphoma 11A (zinc finger protein) | −3.983 | −1.759 | DNA binding protein |
| Tgfb1i1 | transforming growth factor beta 1 induced transcript 1 | −5.006 | −1.748 | A molecular adapter involved in coordination of multiple protein–protein interactions |
| Cxcl12 | chemokine (C-X-C motif) ligand 12 | −3.073 | −1.685 | A ligand for CXC chemokine receptor 4 involved in neuronal regeneration in the adult brain |
| Cckbr | cholecystokinin B receptor | −3.012 | −1.683 | In the brain it is crucial for cortical development |
| Tm4sf1 | transmembrane 4 superfamily member 1 | −3.198 | −1.681 | A cell surface glycoprotein involved in cell–cell interactions |
| Tbcc | tubulin-specific chaperone C | −5.848 | −1.629 | Involved in structural rearrangements of tubulin and actin during the cell cycle |
| Aif1 | allograft inflammatory factor 1 | −3.263 | −1.627 | A calcium-binding adapter molecule specifically expressed in microglia. |
| Trim59 | tripartite motif-containing 59 | −4.371 | −1.602 | Functions as a ubiquitination ligase; its overexpression stimulates cell proliferation and migration |
| Pbx2 | pre B cell leukemia homeobox 2 | −5.499 | −1.579 | A transcription factor required for normal brain development |
| Kif21b | kinesin family member 21B | −4.166 | −1.497 | An ATP-dependent microtubule-based motor protein involved in the intracellular transport processes |
| Rapgef3 | Rap guanine nucleotide exchange factor (GEF) 3 | −3.312 | −1.488 | Involved in brain development and brain diseases |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medvedev, A.; Kopylov, A.; Buneeva, O.; Kurbatov, L.; Tikhonova, O.; Ivanov, A.; Zgoda, V. A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research. Int. J. Mol. Sci. 2020, 21, 4187. https://doi.org/10.3390/ijms21114187
Medvedev A, Kopylov A, Buneeva O, Kurbatov L, Tikhonova O, Ivanov A, Zgoda V. A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research. International Journal of Molecular Sciences. 2020; 21(11):4187. https://doi.org/10.3390/ijms21114187
Chicago/Turabian StyleMedvedev, Alexei, Arthur Kopylov, Olga Buneeva, Leonid Kurbatov, Olga Tikhonova, Alexis Ivanov, and Victor Zgoda. 2020. "A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research" International Journal of Molecular Sciences 21, no. 11: 4187. https://doi.org/10.3390/ijms21114187
APA StyleMedvedev, A., Kopylov, A., Buneeva, O., Kurbatov, L., Tikhonova, O., Ivanov, A., & Zgoda, V. (2020). A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research. International Journal of Molecular Sciences, 21(11), 4187. https://doi.org/10.3390/ijms21114187