Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus


 and
and
Abstract
1. Introduction
2. Results
2.1. Specificity of Anti-CD4-2 mAb (3C8)
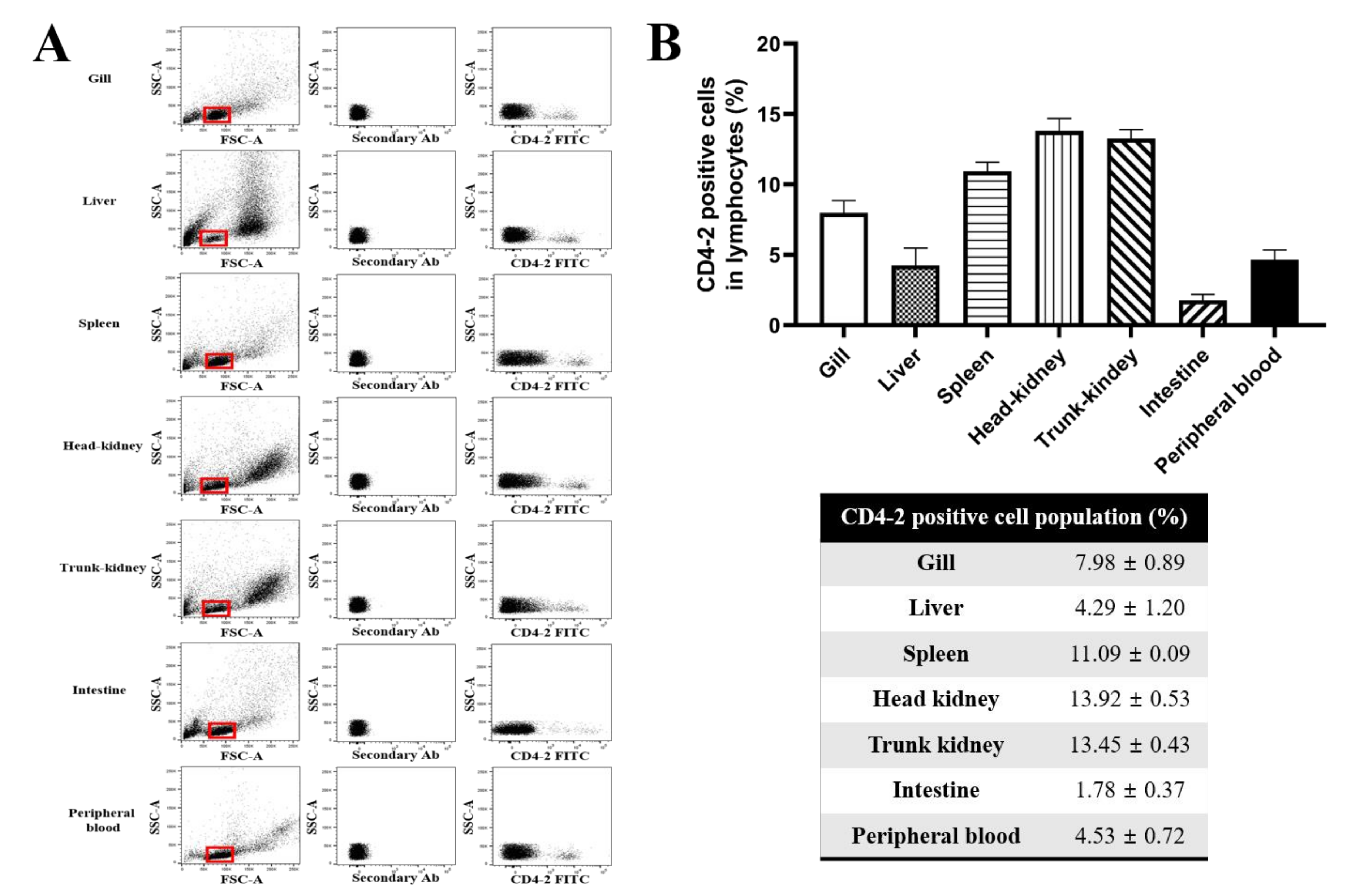
2.2. Tissue Distribution of CD4-2-Positive Lymphocytes Analyzed by Flow Cytometry
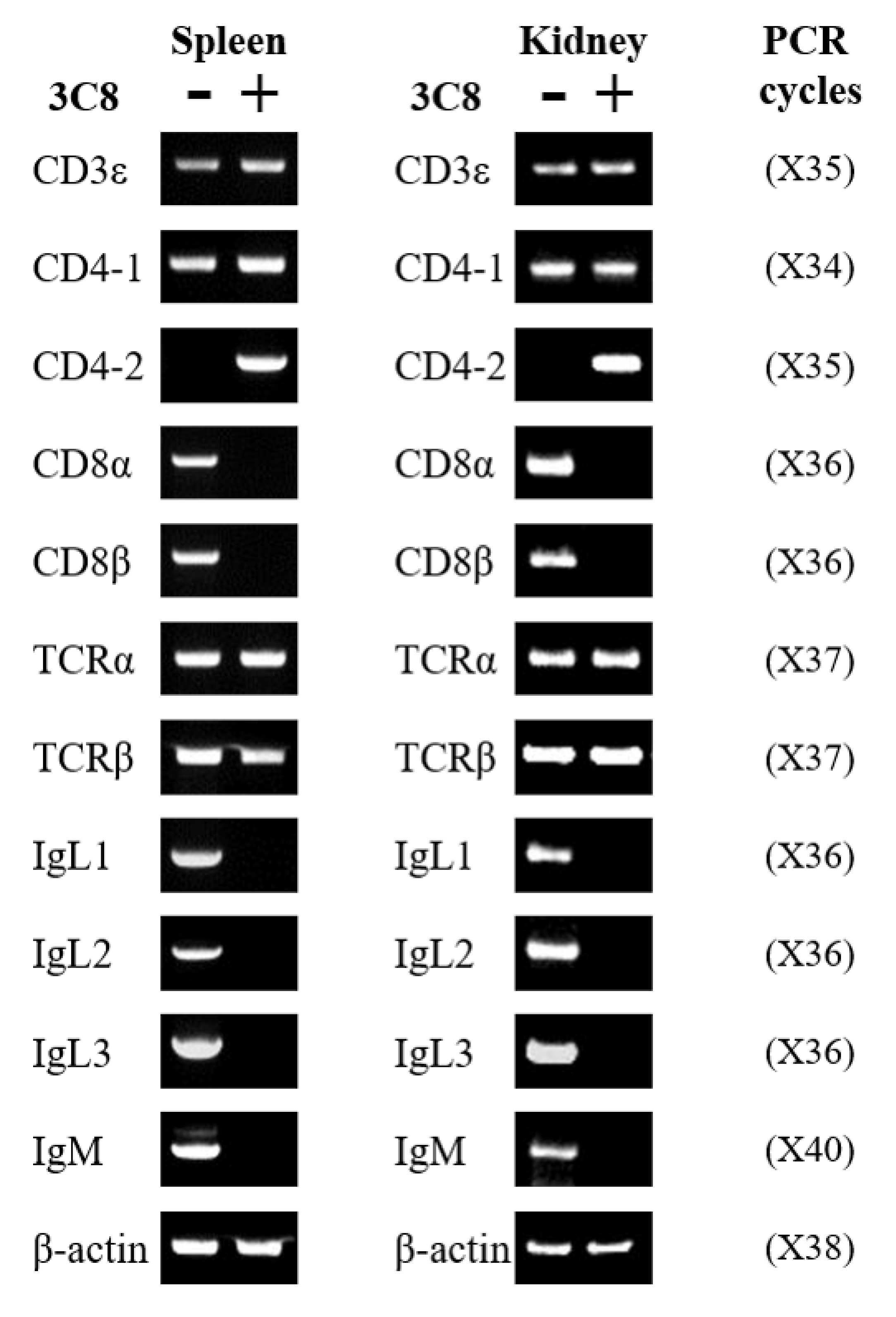
2.3. Expression of mRNA Profiles in CD4-2-Positive Lymphocytes
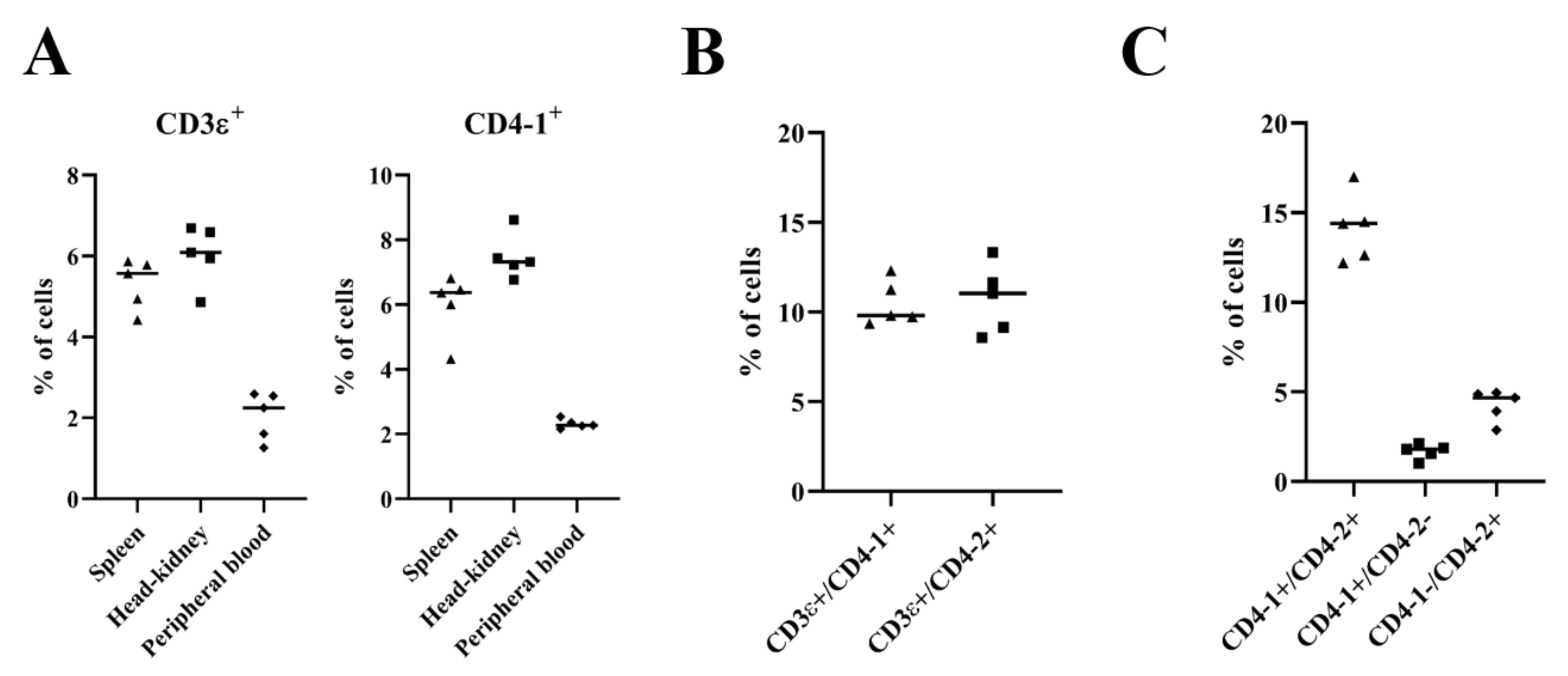
2.4. Two-Color Flow Cytometry to Confirm That the Staining of mAb 3C8 Was Specific for CD4-2 Lymphocytes in Olive Flounder
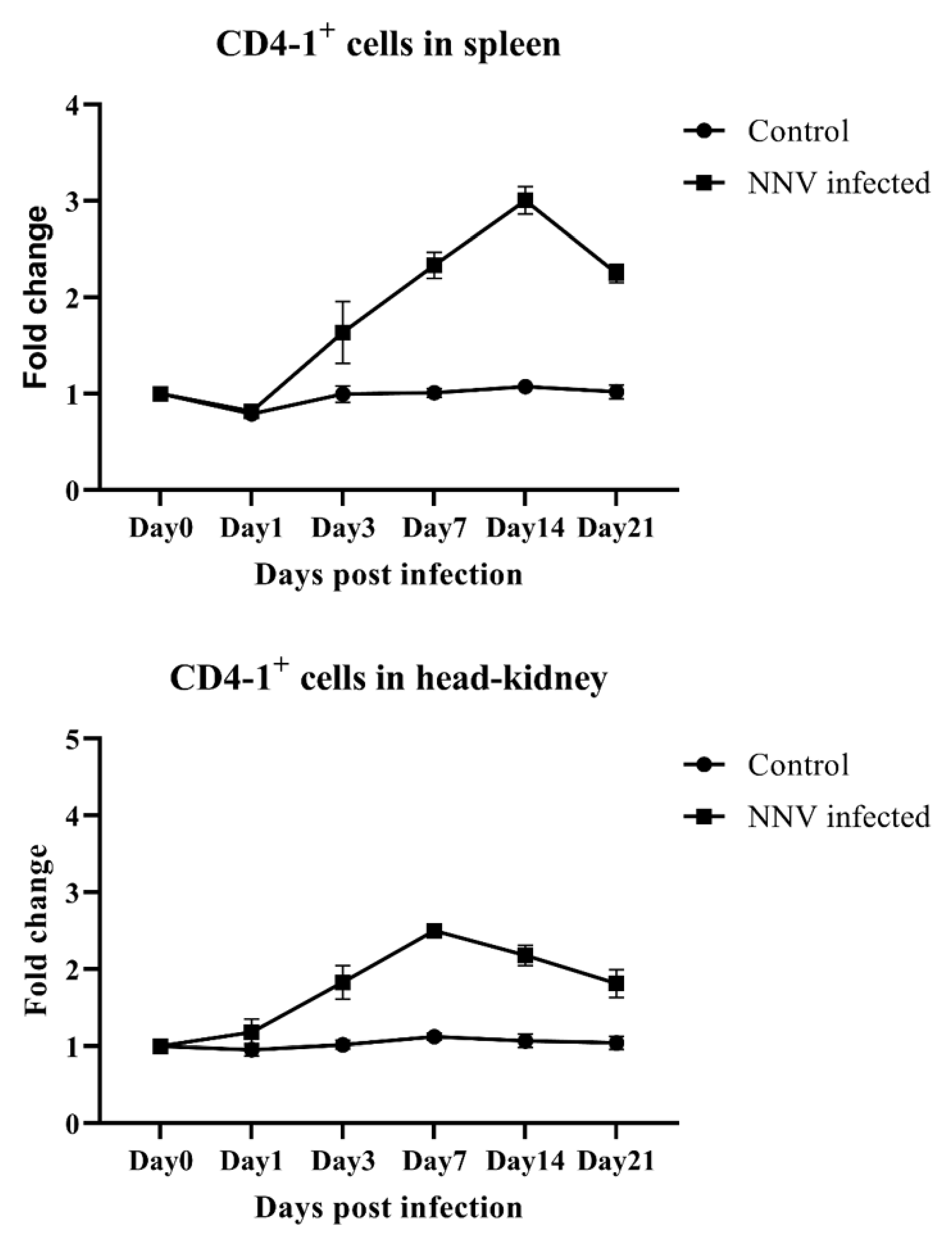
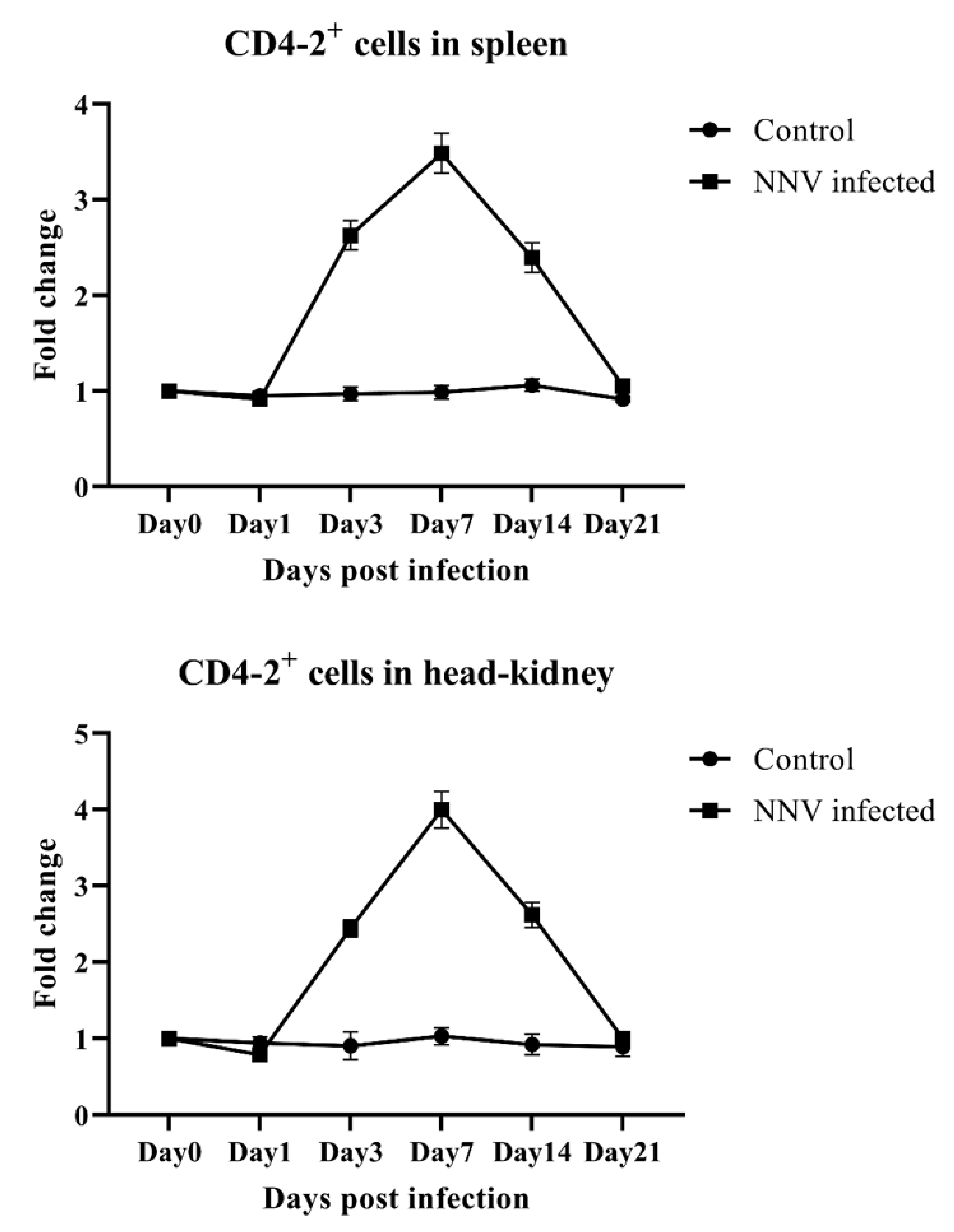
2.5. Response of CD4-1- and CD4-2-Positive Lymphocytes During an NNV Infection in Olive Flounder Determined by Flow Cytometry
3. Discussion
4. Materials and Methods
4.1. Expression and Purification of the Recombinant CD4-2 Antigen
4.2. Production of mAb (3C8) Specific to CD4-2 Lymphocytes from Olive Flounder
4.3. Preparation of Olive Flounder Leukocytes
4.4. Transfection
4.5. Western Blotting
4.6. Flow Cytometry
4.7. Immunofluorescence Staining
4.8. RT-PCR with Flow Cytometry Sorted Leukocytes
4.9. Assessing Populations of CD4-1- and CD4-2-Positive Lymphocytes in Olive Flounder Infected with NNV
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jung, J.W.; Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, Y.R.; Chun, J.H.; Ha, M.W.; Kim, N.N. Involvement of CD4-1 T cells in the cellular immune response of olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) and nervous necrosis virus (NNV) infection. Dev. Comp. Immunol. 2020, 103, 103518. [Google Scholar] [CrossRef]
- Takizawa, F.; Dijkstra, J.M.; Kotterba, P.; Korytář, T.; Kock, H.; Köllner, B.; Jaureguiberry, B.; Nakanishi, T.; Fischer, U. The expression of CD8α discriminates distinct T cell subsets in teleost fish. Dev. Comp. Immunol. 2011, 35, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Somamoto, T.; Kondo, M.; Nakanishi, T.; Nakao, M. Helper function of CD4+ lymphocytes in antiviral immunity in ginbuna crucian carp, Carassius auratus langsdorfii. Dev. Comp. Immunol. 2014, 44, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Laurent, C.; Fazilleau, N.; Brousset, P. A novel subset of T-helper cells: Follicular T-helper cells and their markers. Haematologica 2010, 95, 356. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J.M.; Somamoto, T.; Moore, L.; Hordvik, I.; Ototake, M.; Fischer, U. Identification and characterization of a second CD4-like gene in teleost fish. Mol. Immunol. 2006, 43, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Suetake, H.; Araki, K.; Suzuki, Y. Cloning, expression, and characterization of fugu CD4, the first ectothermic animal CD4. Immunogenetics 2004, 56, 368–374. [Google Scholar] [CrossRef]
- Laing, K.J.; Wang, T.; Zou, J.; Holland, J.; Hong, S.; Bols, N.; Hirono, I.; Aoki, T.; Secombes, C.J. Cloning and expression analysis of rainbow trout Oncorhynchus mykiss tumour necrosis factor-α. Eur. J. Biochem. 2001, 268, 1315–1322. [Google Scholar] [CrossRef]
- Sun, X.-F.; Shang, N.; Hu, W.; Wang, Y.-P.; Guo, Q.-L. Molecular cloning and characterization of carp (Cyprinus carpio L.) CD8β and CD4-like genes. Fish Shellfish Immunol. 2007, 23, 1242–1255. [Google Scholar] [CrossRef]
- Patel, S.; Øvergård, A.-C.; Nerland, A.H. A CD4 homologue in Atlantic halibut (Hippoglossus hippoglossus): Molecular cloning and characterisation. Fish Shellfish Immunol. 2009, 26, 377–384. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Casani, D.; Guerra, L.; Picchietti, S.; Costantini, S.; Facchiano, A.M.; Zou, J.; Secombes, C.J.; Scapigliati, G. A CD4 homologue in sea bass (Dicentrarchus labrax): Molecular characterization and structural analysis. Mol. Immunol. 2008, 45, 3168–3177. [Google Scholar] [CrossRef][Green Version]
- Maisey, K.; Montero, R.; Corripio-Miyar, Y.; Toro-Ascuy, D.; Valenzuela, B.; Reyes-Cerpa, S.; Sandino, A.M.; Zou, J.; Wang, T.; Secombes, C.J. Isolation and characterization of salmonid CD4 + T cells. J. Immunol. 2016, 196, 4150–4163. [Google Scholar] [CrossRef] [PubMed]
- Maddon, P.J.; Molineaux, S.M.; Maddon, D.E.; Zimmerman, K.A.; Godfrey, M.; Alt, F.W.; Chess, L.; Axel, R. Structure and expression of the human and mouse T4 genes. Proc. Natl. Acad. Sci. USA 1987, 84, 9155–9159. [Google Scholar] [CrossRef]
- Kono, T.; Korenaga, H. Cytokine gene expression in CD4 positive cells of the Japanese pufferfish, Takifugu rubripes. PLoS ONE 2013, 8, e66364. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, J.S.; Kim, Y.R.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, J.; Thompson, K.D.; Suh, J.P.; Jung, T.S. Development of a monoclonal antibody against the CD3ε of olive flounder (Paralichthys olivaceus) and its application in evaluating immune response related to CD3ε. Fish Shellfish Immunol. 2017, 65, 179–185. [Google Scholar] [CrossRef]
- Randelli, E.; Buonocore, F.; Scapigliati, G. Cell markers and determinants in fish immunology. Fish Shellfish Immunol. 2008, 25, 326–340. [Google Scholar] [CrossRef]
- Toda, H.; Saito, Y.; Koike, T.; Takizawa, F.; Araki, K.; Yabu, T.; Somamoto, T.; Suetake, H.; Suzuki, Y.; Ototake, M. Conservation of characteristics and functions of CD4 positive lymphocytes in a teleost fish. Dev. Comp. Immunol. 2011, 35, 650–660. [Google Scholar] [CrossRef]
- Xing, J.; Ma, J.; Tang, X.; Sheng, X.; Zhan, W. Characterizations of CD4-1, CD4-2 and CD8β T cell subpopulations in peripheral blood leucocytes, spleen and head kidney of Japanese flounder (Paralichthys olivaceus). Mol. Immunol. 2017, 85, 155–165. [Google Scholar] [CrossRef]
- Xing, J.; Wang, L.; Zhen, M.; Tang, X.; Zhan, W. Variations of T and B lymphocytes of flounder (Paralichthys olivaceus) after Hirame novirhabdovirus infection and immunization. Mol. Immunol. 2018, 96, 19–27. [Google Scholar] [CrossRef]
- Xing, J.; Luo, K.; Tang, X.; Zhan, W. Influence of CD4-1+, CD4-2+ and CD8+ T lymphocytes subpopulations on the immune response of B lymphocytes in flounder (Paralichthys olivaceus) immunized with thymus-dependent or thymus-independent antigen. Fish Shellfish Immunol. 2019, 84, 979–986. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, Y.R.; Chun, J.H.; Ha, M.W.; Kim, H.S.; et al. Characterization of Hagfish (Eptatretus burgeri) Variable Lymphocyte Receptor–Based Antibody and Its Potential Role in the Neutralization of Nervous Necrosis Virus. J. Immunol. 2020, 204, 718–725. [Google Scholar] [CrossRef]
- Kato, G.; Goto, K.; Akune, I.; Aoka, S.; Kondo, H.; Hirono, I. CD4 and CD8 homologues in Japanese flounder, Paralichthys olivaceus: Differences in the expressions and localizations of CD4-1, CD4-2, CD8α and CD8β. Dev. Comp. Immunol. 2013, 39, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Laing, K.J.; Zou, J.J.; Purcell, M.K.; Phillips, R.; Secombes, C.J.; Hansen, J.D. Evolution of the CD4 family: Teleost fish possess two divergent forms of CD4 in addition to lymphocyte activation gene-3. J. Immunol. 2006, 177, 3939–3951. [Google Scholar] [CrossRef] [PubMed]
- Husband, A. Overview of the mammalian immune system. In Advances in Nutritional Research; Springer: Berlin/Heidelberg, Germany, 2001; pp. 3–14. [Google Scholar]
- Laing, K.J.; Hansen, J.D. Fish T cells: Recent advances through genomics. Dev. Comp. Immunol. 2011, 35, 1282–1295. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, R.; Matsuura, Y.; Shibasaki, Y.; Imamura, S.; Nakanishi, T. Cross-reactivity of monoclonal antibodies against CD4-1 and CD8α of ginbuna crucian carp with lymphocytes of zebrafish and other cyprinid species. Dev. Comp. Immunol. 2018, 80, 15–23. [Google Scholar] [CrossRef]
- Kurata, O.; Iwasaki, T.; Matsuyama, T.; Nakayasu, C.; Wada, S.; Hatai, K. Lymphocytes with T-cell-like properties express the Fas ligand in the Japanese flounder Paralichthys olivaceus. Fish Shellfish Immunol. 2011, 30, 509–514. [Google Scholar] [CrossRef]
- Shao, R.; Li, C.S.; Fang, Y.; Zhao, L.; Hang, C. Low B and T lymphocyte attenuator expression on CD4 + T cells in the early stage of sepsis is associated with the severity and mortality of septic patients: A prospective cohort study. Crit. Care 2015, 19, 308. [Google Scholar] [CrossRef]
- Lyons, A.B.; Parish, C.R. Determination of lymphocyte division by flow cytometry. J. Immunol. Methods 1994, 171, 131–137. [Google Scholar] [CrossRef]
- Ellis, A.E. The leucocytes of fish: A review. J. Fish Biol. 1977, 11, 453–491. [Google Scholar] [CrossRef]
- Handin, R.I.; Lux, S.E.; Stossel, T.P. Blood: Principles and Practice of Hematology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2003; Volume 1. [Google Scholar]
- Workenhe, S.T.; Rise, M.L.; Kibenge, M.J.; Kibenge, F.S. The fight between the teleost fish immune response and aquatic viruses. Mol. Immunol. 2010, 47, 2525–2536. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, S.; Jiang, G.; Yang, D.; Lian, J.; Yang, Y. The development of the lymphoid organs of flounder, Paralichthys olivaceus, from hatching to 13 months. Fish Shellfish Immunol. 2004, 16, 621–632. [Google Scholar] [CrossRef]
- Schrøder, M.B.; Villena, A.J.; Jørgensen, T.Ø. Ontogeny of lymphoid organs and immunoglobulin producing cells in Atlantic cod (Gadus morhua L.). Dev. Comp. Immunol. 1998, 22, 507–517. [Google Scholar]
- Petrie-Hanson, L.; Ainsworth, A.J. Ontogeny of channel catfish lymphoid organs. Vet. Immunol. Immunopathol. 2001, 81, 113–127. [Google Scholar] [CrossRef]
- Suresh, N.; Ranganathan, L. Ontogeny of lymphoid organs-thymus, spleen and kidney in Catla catla (Hamilton). Indian J. Fish 2008, 55, 317–322. [Google Scholar]
- Westermann, J.; Pabst, R. Distribution of lymphocyte subsets and natural killer cells in the human body. Clin. Investig. 1992, 70, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, F.; Magadan, S.; Parra, D.; Xu, Z.; Korytář, T.; Boudinot, P.; Sunyer, J.O. Novel teleost CD4-bearing cell populations provide insights into the evolutionary origins and primordial roles of CD4+ lymphocytes and CD4+ macrophages. J. Immunol. 2016, 196, 4522–4535. [Google Scholar] [CrossRef]
- Hannet, I.; Erkeller-Yuksel, F.; Lydyard, P.; Deneys, V.; DeBruyère, M. Developmental and maturational changes in human blood lymphocyte subpopulations. Immunol. Today 1992, 13, 215–218. [Google Scholar] [CrossRef]
- Koppang, E.O.; Fischer, U.; Moore, L.; Tranulis, M.A.; Dijkstra, J.M.; Köllner, B.; Aune, L.; Jirillo, E.; Hordvik, I. Salmonid T cells assemble in the thymus, spleen and in novel interbranchial lymphoid tissue. J. Anat. 2010, 217, 728–739. [Google Scholar] [CrossRef]
- Boardman, T.; Warner, C.; Ramirez-Gomez, F.; Matrisciano, J.; Bromage, E. Characterization of an anti-rainbow trout (Oncorhynchus mykiss) CD3ε monoclonal antibody. Vet. Immunol. Immunopathol. 2012, 145, 511–515. [Google Scholar] [CrossRef]
- Edholm, E.-S.; Stafford, J.L.; Quiniou, S.M.; Waldbieser, G.; Miller, N.W.; Bengtén, E.; Wilson, M. Channel catfish, Ictalurus punctatus, CD4-like molecules. Dev. Comp. Immunol. 2007, 31, 172–187. [Google Scholar] [CrossRef]
- Chan, M.M.; Chen, C.; Ager, L.L.; Cooper, M.D. Identification of the avian homologues of mammalian CD4 and CD8 antigens. J. Immunol. 1988, 140, 2133–2138. [Google Scholar]
- Yong, C.Y.; Yeap, S.K.; Omar, A.R.; Tan, W.S. Advances in the study of nodavirus. Peer J. 2017, 5, e3841. [Google Scholar] [CrossRef]
- López-Muñoz, A.; Sepulcre, M.P.; García-Moreno, D.; Fuentes, I.; Béjar, J.; Manchado, M.; Álvarez, M.C.; Meseguer, J.; Mulero, V. Viral nervous necrosis virus persistently replicates in the central nervous system of asymptomatic gilthead seabream and promotes a transient inflammatory response followed by the infiltration of IgM + B lymphocytes. Dev. Comp. Immunol. 2012, 37, 429–437. [Google Scholar] [CrossRef]
- Guidotti, L.G.; Chisari, F.V. Noncytolytic control of viral infections by the innate and adaptive immuneresponse. Ann. Rev. Immunol. 2001, 19, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C. Will advances in fish immunology change vaccination strategies? Fish Shellfish Immunol. 2008, 25, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, D.A.; Lewinsohn, D.M.; Scriba, T.J. Polyfunctional CD4 + T cells as targets for tuberculosis vaccination. Front. Immunol. 2017, 8, 1262. [Google Scholar] [CrossRef]
- Panagioti, E.; Klenerman, P.; Lee, L.N.; Van Der Burg, S.H.; Arens, R. Features of effective T cell-inducing vaccines against chronic viral infections. Front. Immunol. 2018, 9, 276. [Google Scholar] [CrossRef]
- Harlow, E.; Lane, D. A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1988; p. 579. [Google Scholar]
- Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Jung, J.W.; Lazarte, J.M.S.; Lee, J.H.; Thompson, K.D.; Jung, T.S. Dual functionality of lamprey VLRB C-terminus (LC) for multimerization and cell surface display. Mol. Immunol. 2018, 104, 54–60. [Google Scholar] [CrossRef]
- Kim, J.; Im, S.P.; Lee, J.S.; Lazarte, J.M.S.; Kim, S.W.; Jung, J.W.; Kim, J.Y.; Kim, Y.R.; Lee, S.; Kim, G.J. Globular-shaped variable lymphocyte receptors B antibody multimerized by a hydrophobic clustering in hagfish. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | |
|---|---|---|
| CD3ε (AB081751.1) | Forward | ATGAAAATCAACACCATGGATGTC |
| Reverse | TCCCGTCCTGTTCACAATAGA | |
| CD4-1 (AB643634.1) | Forward | ATGAATCCCAGAGGAGAGATAATG |
| Reverse | CACGTAGTCTCCTCCGTCTTC | |
| CD4-2 (AB640684.1) | Forward | GTGATCCTAACAAAACCCAGGCAG |
| Reverse | AGCAGGTTCTTCAACTTTGATCTT | |
| CD8α (AB082957.1) | Forward | ATGGACCAAAAGTGGATTCAGATG |
| Reverse | AACATGTGTGTTGTTCTTCATCTG | |
| CD8β (AB643633.1) | Forward | ATGAACCCGCTGCCGCTG |
| Reverse | GGGCATCTGTCTCATCTTCTG | |
| TCRα (AB053227.1) | Forward | ATGCTCTCACTGCATCTTGGT |
| Reverse | GACTCTGTGACTGAGCCACAG | |
| TCRβ (AB053228.1) | Forward | ATGATTCCAAGCCTCAACACC |
| Reverse | GTGGTTCTGCTTCTCAGCTGA | |
| IgL1 (AB819734.1) | Forward | ATGAGCTTTACCTCCGTCCTC |
| Reverse | GGACTGGGAACACTGGTCTCT | |
| IgL2 (AB819735.1) | Forward | ATGATGGTTTTTCTGAGTCAGGAG |
| Reverse | CTCCGAGCAGCGGTCAGG | |
| IgL3 (AB819736.1) | Forward | ATGCTGGGGACCCTCTGC |
| Reverse | GTGGTACAGACGGACTTGTTG | |
| IgM (AB052744.1) | Forward | ATGTTTCCTGTAGCTGTGCTG |
| Reverse | CTGGGCCTTGCATGGTATGTT | |
| β-actin (HQ386788.1) | Forward | ATGGAAGATGAAATCGCCGCA |
| Reverse | GAAGCATTTGCGGTGGACGAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.W.; Chun, J.H.; Lee, J.S.; Kim, S.W.; Lee, A.R.; Kim, J.; Lazarte, J.M.S.; Kim, Y.R.; Kim, H.J.; Thompson, K.D.; et al. Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus. Int. J. Mol. Sci. 2020, 21, 4180. https://doi.org/10.3390/ijms21114180
Jung JW, Chun JH, Lee JS, Kim SW, Lee AR, Kim J, Lazarte JMS, Kim YR, Kim HJ, Thompson KD, et al. Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus. International Journal of Molecular Sciences. 2020; 21(11):4180. https://doi.org/10.3390/ijms21114180
Chicago/Turabian StyleJung, Jae Wook, Jin Hong Chun, Jung Seok Lee, Si Won Kim, Ae Rin Lee, Jaesung Kim, Jassy Mary S. Lazarte, Young Rim Kim, Hyoung Jun Kim, Kim D. Thompson, and et al. 2020. "Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus" International Journal of Molecular Sciences 21, no. 11: 4180. https://doi.org/10.3390/ijms21114180
APA StyleJung, J. W., Chun, J. H., Lee, J. S., Kim, S. W., Lee, A. R., Kim, J., Lazarte, J. M. S., Kim, Y. R., Kim, H. J., Thompson, K. D., & Jung, T. S. (2020). Characterization of CD4-Positive Lymphocytes in the Antiviral Response of Olive Flounder (Paralichthys oliveceus) to Nervous Necrosis Virus. International Journal of Molecular Sciences, 21(11), 4180. https://doi.org/10.3390/ijms21114180