Profound Nanoscale Structural and Biomechanical Changes in DNA Helix upon Treatment with Anthracycline Drugs

 ,
,  , , , , , ,
, , , , , ,
Abstract

1. Introduction
2. Results and Discussion
2.1. In Silico Analysis Reveals Diverse Modes of Interactions of DNA with Anthracyclines
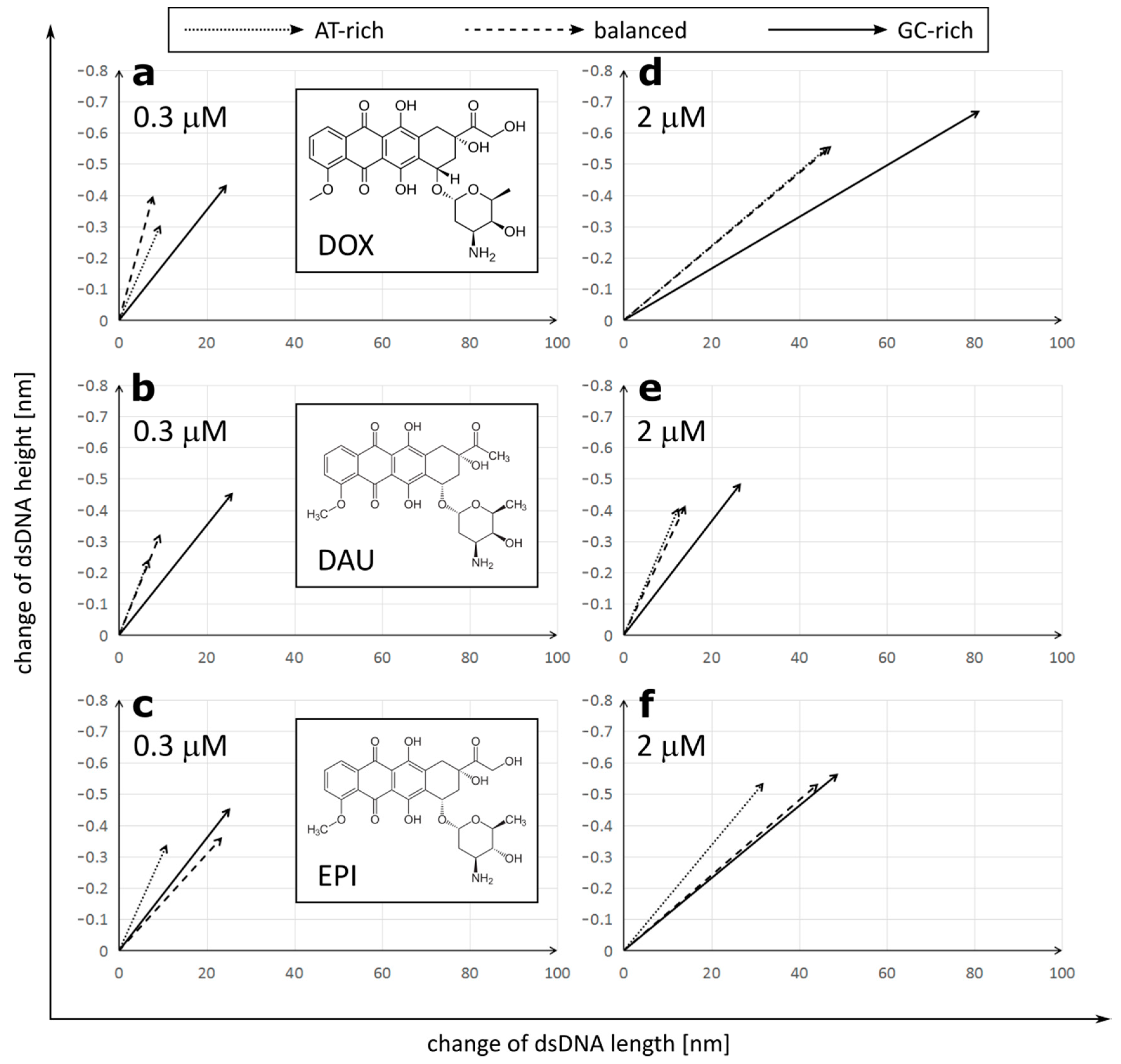
2.2. Anthracyclines Increase Length and Decrease Height of dsDNA Molecule
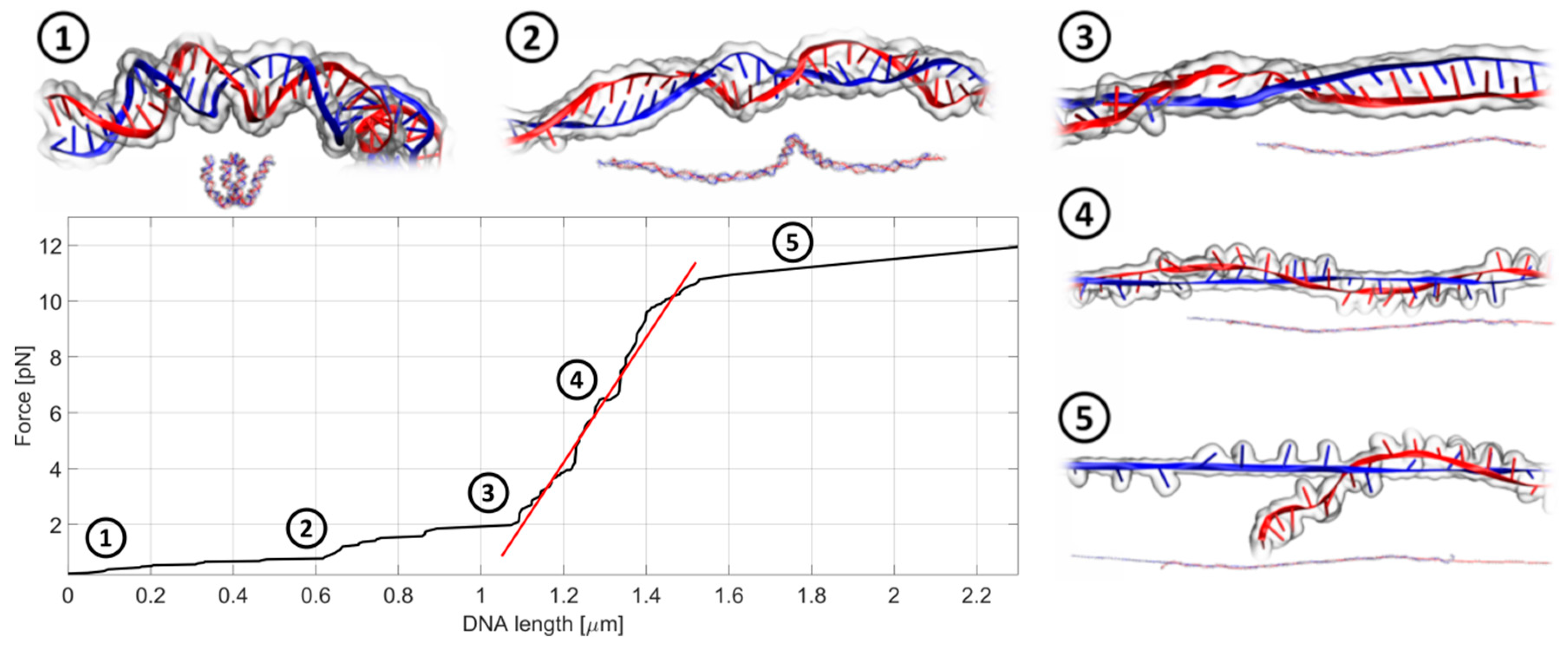
2.3. Anthracyclines Induce Substantial Decrease of dsDNA Stiffness
2.4. Biophysical Changes Are Reflected by Changes of Nuclei Size and Stiffness
3. Materials and Methods
3.1. Materials
3.2. The Structural Study of the Anthracycline Interactions with Double Helix of DNA
3.2.1. Preparation of Samples for Studying Structural Changes of DNA Using AFM
3.2.2. AFM Imaging Technique
3.2.3. AFM Data Analysis
3.3. Changes in DNA Stiffness upon Treatment with Anthracycline Antibiotics
3.3.1. Preparation of Samples for Studying Mechanical Changes of DNA Using Optical Tweezers
3.3.2. OT Technique
3.4. Cell Nuclei Isolation for Optical Tweezers Studies
3.5. Molecular Dynamics
4. Conclusions
- All anthracyclines affected the topology of the dsDNA molecule, lengthening and decreasing the height of the dsDNA molecule;
- The effect that anthracyclines exert on the dsDNA topology was dependent on the GC/AT ratio in dsDNA, and was strongest for the GC-rich dsDNA, suggesting preferential intercalation for GC pairs;
- Despite the high structural similarity of the DOX, DAU and EPI anthracyclines, they differ in the extent of their effect on the topology and their dependence on the composition of dsDNA. DOX exerts the strongest effect on dsDNA topology. DOX and DAU had a preference for GC-rich DNA, while EPI caused similar disturbances in GC-rich and balanced dsDNA topology;
- The malformations of the DNA topology were accompanied by a reduction in DNA stiffness and its susceptibility to break. The worsening of the biomechanical properties of dsDNA is dependent on the dsDNA composition and structure of the interacting molecule. The GC-rich dsDNA was especially affected by anthracyclines, once more indicating a preferential interaction with GC pairs. This is especially important because DNA regulatory elements are often located in GC-rich regions of genomes.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cortes-Funes, H.; Coronado, C. Role of anthracyclines in the era of targeted therapy. Cardiovasc. Toxicol. 2007, 7, 56–60. [Google Scholar] [CrossRef]
- Edwardson, D.W.; Narendrula, R.; Chewchuk, S.; Mispel-Beyer, K.; Mapletoft, J.P.; Parissenti, A.M. Role of drug metabolism in the cytotoxicity and clinical efficacy of anthracyclines. Curr. Drug Metab. 2015, 16, 412–426. [Google Scholar] [CrossRef]
- Tomankova, K.; Polakova, K.; Pizova, K.; Binder, S.; Havrdova, M.; Kolarova, M.; Kriegova, E.; Zapletalova, J.; Malina, L.; Horakova, J.; et al. In Vitro cytotoxicity analysis of doxorubicin-loaded/superparamagnetic iron oxide colloidal nanoassemblies on MCF7 and NIH3T3 cell lines. Int. J. Nanomed. Nanosurg. 2015, 10, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Bose, R.; Verheij, M.; Haimovitz-Friedman, A.; Scotto, K.; Fuks, Z.; Kolesnick, R. Ceramide synthase mediates daunorubicin-induced apoptosis: An alternative mechanism for generating death signals. Cell 1995, 82, 405–414. [Google Scholar] [CrossRef]
- Jaffrézou, J.P.; Levade, T.; Bettaïeb, A.; Andrieu, N.; Bezombes, C.; Maestre, N.; Vermeersch, S.; Rousse, A.; Laurent, G. Daunorubicin-induced apoptosis: Triggering of ceramide generation through sphingomyelin hydrolysis. EMBO J. 1996, 15, 2417–2424. [Google Scholar] [CrossRef]
- Kiyomiya, K.I.; Matsuo, S.; Kurebe, M. Mechanism of specific nuclear transport of adriamycin. Cancer Res. 2001, 61, 2467. [Google Scholar] [PubMed]
- Capelôa, T.; Benyahia, Z.; Zampieri, L.X.; Blackman, M.C.N.M.; Sonveaux, P. Metabolic and non-metabolic pathways that control cancer resistance to anthracyclines. Semin. Cell Dev. Bio 2020, 98, 181–191. [Google Scholar] [CrossRef]
- Pilch, D.S.; Kirolos, M.A.; Liu, X.; Plum, G.E.; Breslauer, K.J. Berenil [1,3-bis(4′-amidinophenyl)triazene] binding to DNA duplexes and to a RNA duplex: Evidence for both intercalative and minor groove binding properties. Biochemistry 1995, 34, 9962–9976. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, A.; Finn, R.M.; Ausio, J. The anthracycline antibiotics: Antitumor drugs that alter chromatin structure. BioEssays 2004, 27, 50–56. [Google Scholar] [CrossRef]
- Li, L.; Zhang, P.; Li, J.; Wang, Y.; Wei, Y.; Hu, J.; Zhou, X.; Xu, B.; Li, B. Measurement of nanomechanical properties of DNA molecules by PeakForce atomic force microscopy based on DNA origami. Nanoscale 2019, 11, 4707–4711. [Google Scholar] [CrossRef] [PubMed]
- Strekowski, L.; Wilson, B. Noncovalent interactions with DNA: An overview. Mutat. Res. 2007, 623, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Arnaiz, C.; Busto, N.; Leal, J.M.; García, B. New insights into the mechanism of the DNA/doxorubicin interaction. J. Phys. Chem. B. 2014, 118, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Xu, S. Sequence and Chiral Selectivity of Drug-DNA Interactions Revealed by Force Spectroscopy. Angew. Chem. Int. Ed. Engl. 2014, 53, 14135–14138. [Google Scholar] [CrossRef] [PubMed]
- Edwardson, J.M.; Henderson, R.M. Atomic force microscopy and drug discovery. Drug Discov. Today 2004, 9, 64–71. [Google Scholar] [CrossRef]
- Sirajuddin, M.; Haider, A.; Ali, S. Analytical Techniques for The Study of Drug-DNA Interactions. Int. J. Adv. Res. 2014, 1, 510–530. [Google Scholar]
- Berge, T.; Haken, E.L.; Waring, M.J.; Henderson, R.M. The binding mode of the DNA bisintercalator luzopeptin investigated using atomic force microscopy. J. Struct. Biol. 2003, 142, 241–246. [Google Scholar] [CrossRef]
- Kopaczyńska, M.; Schulz, A.; Frączkowska, K.; Kraszewski, S.; Podbielska, H.; Fuhrhop, J.H. Selective condensation of DNA by aminoglycoside antibiotics. Eur. Biophys. J. 2016, 45, 287–299. [Google Scholar] [CrossRef]
- Heller, I.; Hoekstra, T.P.; King, G.A.; Peterman, E.J.G.; Wuite, G.J.L. Optical tweezers analysis of DNA-protein complexes. Chem. Rev. 2014, 114, 3087–3119. [Google Scholar] [CrossRef]
- Salerno, D.; Beretta, G.L.; Zanchetta, G.; Brioschi, S.; Cristofalo, M.; Missana, N.; Nardo, L.; Cassina, V.; Tempestini, A.; Giovannoni, R.; et al. Platinum-Based Drugs and DNA Interactions Studied by Single-Molecule and Bulk Measurements. Biophys. J. 2016, 110, 2151–2161. [Google Scholar] [CrossRef]
- Wang, M.D.; Yin, H.; Landick, R.; Gelles, J.; Block, S.M. Stretching DNA with optical tweezers. Biophys. J. 1997, 72, 1335–1346. [Google Scholar] [CrossRef]
- Smith, S.; Cui, Y.; Bustamante, C. Overstretching B-DNA: The Elastic Response of Individual Double-Stranded and Single-Stranded DNA Molecules. Science 1996, 271, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Bennink, M.L.; Leuba, S.H.; Leno, G.H.; Zlatanova, J.; de Grooth, B.G.; Greve, J. Unfolding individual nucleosomes by stretching single chromatin fibers with optical tweezers. Nat. Struct. Mol. Biol. 2001, 8, 606. [Google Scholar] [CrossRef] [PubMed]
- Strick, T.; Allemand, J.F.; Croquette, V.; Bensimon, D. Twisting and stretching single DNA molecules. Prog. Biophys. Mol. Biol. 2000, 74, 115–140. [Google Scholar] [CrossRef]
- Frączkowska, K.; Bacia, M.; Przybyło, M.; Drabik, D.; Kaczorowska, A.; Rybka, J.; Stefanko, E.; Drobczyński, S.; Masajada, J.; Podbielska, H.; et al. Alterations of biomechanics in cancer and normal cells induced by doxorubicin. Biomed. Pharmacother. 2018, 97, 1195–1203. [Google Scholar] [CrossRef]
- Ren, J.; Chaires, J.B. Sequence and Structural Selectivity of Nucleic Acid Binding Ligands. Biochemistry 1999, 38, 16067–16075. [Google Scholar] [CrossRef]
- Cheng, C.M.; Lee, Y.J.; Wang, W.T.; Hsu, C.T.; Tsai, J.S.; Wu, C.M.; Ou, K.L.; Yang, T.S. Determining the binding mode and binding affinity constant of tyrosine kinase inhibitor PD153035 to DNA using optical tweezers. Biochem. Bioph Res. Co 2011, 404, 297–301. [Google Scholar] [CrossRef]
- Crisafuli, F.A.P.; Cesconetto, E.C.; Ramos, E.B.; Rocha, M.S. DNA–cisplatin interaction studied with single molecule stretching experiments. Integr. Biol. 2012, 4, 568–574. [Google Scholar] [CrossRef]
- Maaloum, M.; Muller, P.; Harlepp, S. DNA-intercalator interactions: Structural and physical analysis using atomic force microscopy in solution. Soft Matter 2013, 9, 11233–11240. [Google Scholar] [CrossRef]
- Bosire, R.; Nánási, P.; Imre, L.; Dienes, B.; Szöőr, Á.; Mázló, A.; Kovács, A.; Seidel, R.; Vámosi, G.; Szabó, G. Intercalation of small molecules into DNA in chromatin is primarily controlled by superhelical constraint. PLoS ONE 2019, 14, e0224936. [Google Scholar] [CrossRef]
- Lamperska, W.; Masajada, J.; Drobczyński, S.; Gusin, P. Two-laser optical tweezers with a blinking beam. Opt. Laser Eng. 2017, 94, 82–89. [Google Scholar] [CrossRef]
- Drobczyński, S.; Duś-Szachniewicz, K. Real-time force measurement in double wavelength optical tweezers. JOSA B 2017, 34, 38–43. [Google Scholar] [CrossRef]
- Make-Na Server. Available online: http://structure.usc.edu/make-na/server.html (accessed on 30 September 2019).
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision B.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNA Stiffness [pN/µm] | |||
|---|---|---|---|
| Sample | AT-Rich | Balanced | GC-Rich |
| Untreated | 24 ± 5 | 21 ± 4 | 22 ± 5 |
| DOX | 18 ± 5 | 14 ± 3 | 8 ± 3 |
| DAU | 12 ± 1 | 19 ± 3 | 11 ± 4 |
| EPI | 12 ± 1 | 15 ± 4 | 10 ± 3 |
| DNA Breakage Force [pN] | |||
| Sample | AT-Rich | Balanced | GC-Rich |
| Untreated | - | 8.0 ± 1.5 | 12.6 ± 1.5 |
| DOX | - | 11.0 ± 2.5 | 9.0 ± 1.8 |
| DAU | - | 9.0 ± 1.5 | 9.6 ± 1.5 |
| EPI | 8.5 ± 4.5 | 11.0 ± 0.5 | 9.3 ± 1.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczorowska, A.; Lamperska, W.; Frączkowska, K.; Masajada, J.; Drobczyński, S.; Sobas, M.; Wróbel, T.; Chybicka, K.; Tarkowski, R.; Kraszewski, S.; et al. Profound Nanoscale Structural and Biomechanical Changes in DNA Helix upon Treatment with Anthracycline Drugs. Int. J. Mol. Sci. 2020, 21, 4142. https://doi.org/10.3390/ijms21114142
Kaczorowska A, Lamperska W, Frączkowska K, Masajada J, Drobczyński S, Sobas M, Wróbel T, Chybicka K, Tarkowski R, Kraszewski S, et al. Profound Nanoscale Structural and Biomechanical Changes in DNA Helix upon Treatment with Anthracycline Drugs. International Journal of Molecular Sciences. 2020; 21(11):4142. https://doi.org/10.3390/ijms21114142
Chicago/Turabian StyleKaczorowska, Aleksandra, Weronika Lamperska, Kaja Frączkowska, Jan Masajada, Sławomir Drobczyński, Marta Sobas, Tomasz Wróbel, Kinga Chybicka, Radosław Tarkowski, Sebastian Kraszewski, and et al. 2020. "Profound Nanoscale Structural and Biomechanical Changes in DNA Helix upon Treatment with Anthracycline Drugs" International Journal of Molecular Sciences 21, no. 11: 4142. https://doi.org/10.3390/ijms21114142
APA StyleKaczorowska, A., Lamperska, W., Frączkowska, K., Masajada, J., Drobczyński, S., Sobas, M., Wróbel, T., Chybicka, K., Tarkowski, R., Kraszewski, S., Podbielska, H., Kałas, W., & Kopaczyńska, M. (2020). Profound Nanoscale Structural and Biomechanical Changes in DNA Helix upon Treatment with Anthracycline Drugs. International Journal of Molecular Sciences, 21(11), 4142. https://doi.org/10.3390/ijms21114142