Regulation of Deubiquitinating Enzymes by Post-Translational Modifications


Abstract
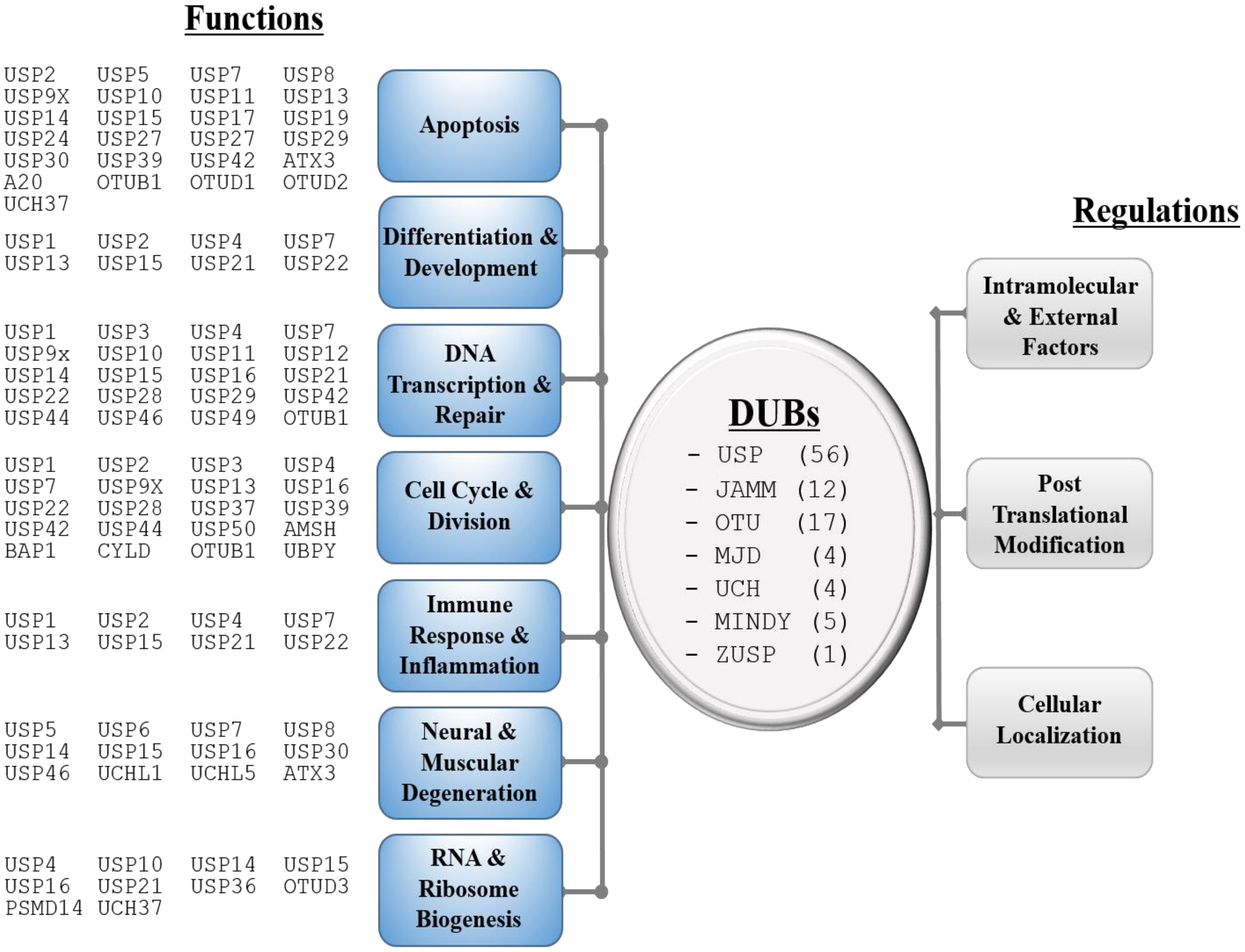
1. Introduction
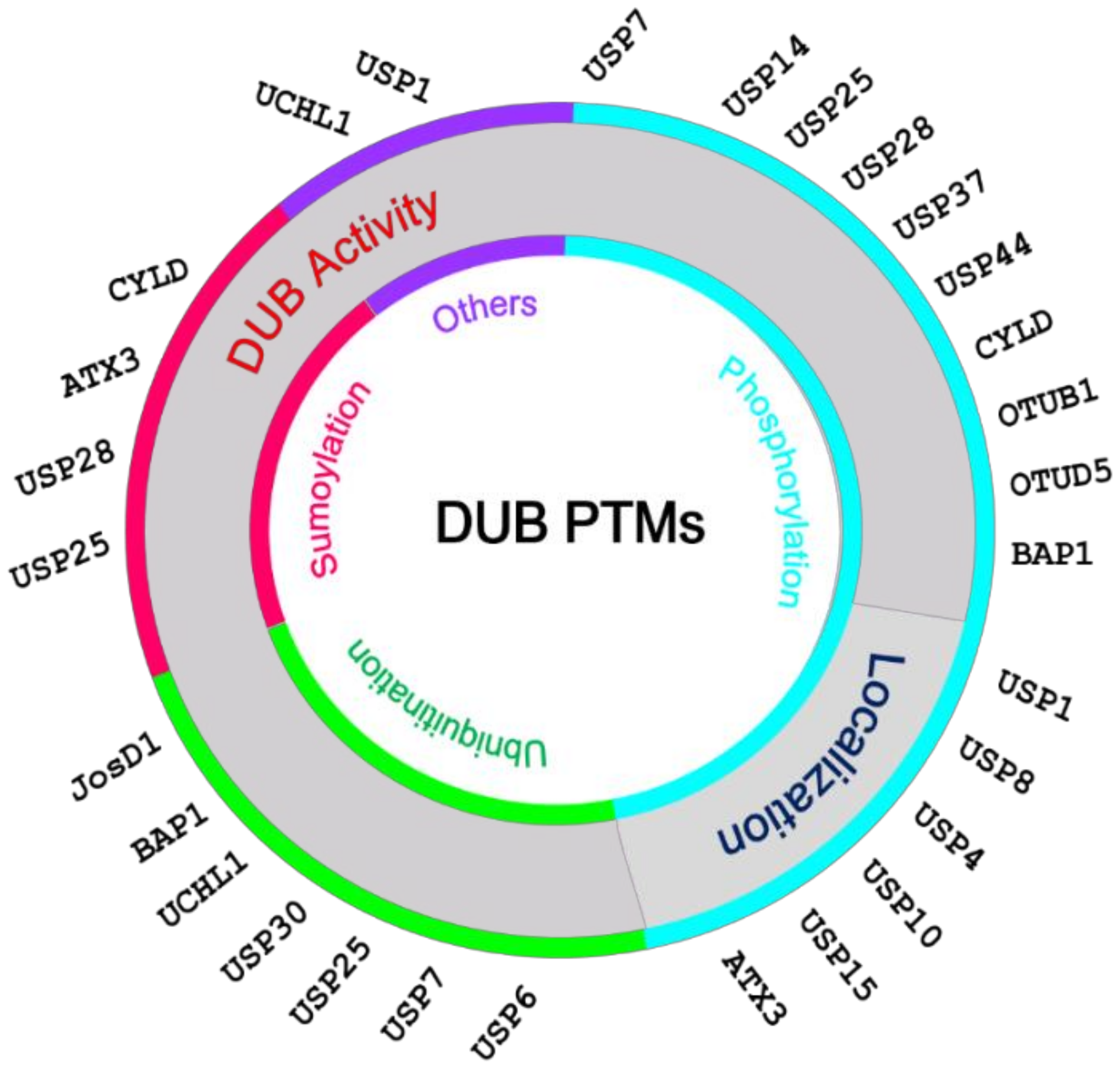
2. Post-Translational Modifications of DUBs
2.1. PTMs Regulating the Catalytic Activity of DUBs
2.2. PTMs Regulating the Subcellular Localization of DUBs
2.3. PTMs Regulating DUBs Interaction with Partner Proteins
2.4. PTMs Regulating DUB Stability and Abundance
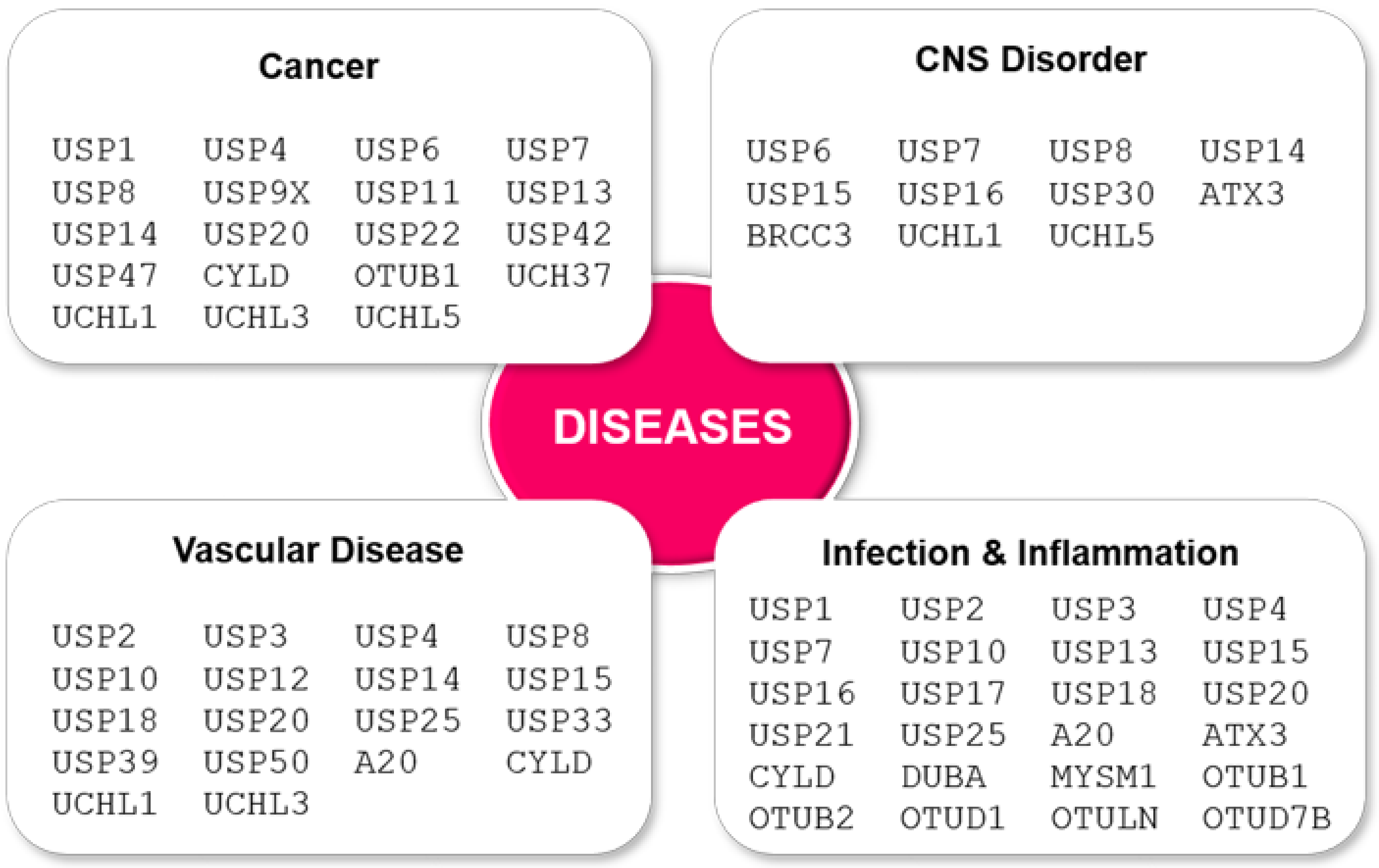
3. Misregulation of DUB PTM May Lead to Disease
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PTM | post translational modification |
| DUB | deubiquitinating enzyme |
| Ub | ubiquitin |
| USP | ubiquitin-specific protease |
| JAMM | JAB1/MPN/Mov34 metalloenzymes |
| OTU | ovarian tumor protease |
| UCH | ubiquitin C-terminal hydrolase |
| MJP | Josephin and JAB1/MPN+ |
| MINDY | MIU-containing novel DUB |
| ZUP1 | zinc finger-containing ubiquitin peptidase 1 |
| SUMO | small ubiquitin-like modifier |
| UBA | ubiquitin associated domain |
| UBL | ubiquitin-like |
| UIM | ubiquitin interacting motif |
| MJD | Machado-Josephin domain protease |
References
- Pickart, C.M. Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta 2004, 1695, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Clague, M.J.; Barsukov, I.; Coulson, J.M.; Liu, H.; Rigden, D.J.; Urbe, S. Deubiquitylases from genes to organism. Physiol. Rev. 2013, 93, 1289–1315. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Tcherpakov, M. Will the ubiquitin system furnish as many drug targets as protein kinases? Cell 2010, 143, 686–693. [Google Scholar] [CrossRef]
- D’Andrea, A.; Pellman, D. Deubiquitinating enzymes: A new class of biological regulators. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 337–352. [Google Scholar] [CrossRef]
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell. Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef]
- Clague, M.J.; Urbe, S.; Komander, D. Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 2019, 20, 338–352. [Google Scholar] [CrossRef]
- Mevissen, T.E.T.; Komander, D. Mechanisms of Deubiquitinase Specificity and Regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef]
- Sahtoe, D.D.; Sixma, T.K. Layers of DUB regulation. Trends Biochem. Sci. 2015, 40, 456–467. [Google Scholar] [CrossRef]
- Kessler, B.M.; Edelmann, M.J. PTMs in conversation: Activity and function of deubiquitinating enzymes regulated via post-translational modifications. Cell Biochem. Biophys. 2011, 60, 21–38. [Google Scholar] [CrossRef]
- Leznicki, P.; Kulathu, Y. Mechanisms of regulation and diversification of deubiquitylating enzyme function. J. Cell Sci. 2017, 130, 1997–2006. [Google Scholar] [CrossRef] [PubMed]
- Hunter, T. The age of crosstalk: Phosphorylation, ubiquitination, and beyond. Mol. Cell 2007, 28, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Khoury, G.A.; Baliban, R.C.; Floudas, C.A. Proteome-wide post-translational modification statistics: Frequency analysis and curation of the swiss-prot database. Sci. Rep. 2011, 1. [Google Scholar] [CrossRef] [PubMed]
- Leggett, D.S.; Hanna, J.; Borodovsky, A.; Crosas, B.; Schmidt, M.; Baker, R.T.; Walz, T.; Ploegh, H.; Finley, D. Multiple associated proteins regulate proteasome structure and function. Mol. Cell 2002, 10, 495–507. [Google Scholar] [CrossRef]
- Xu, D.; Shan, B.; Lee, B.H.; Zhu, K.; Zhang, T.; Sun, H.; Liu, M.; Shi, L.; Liang, W.; Qian, L.; et al. Phosphorylation and activation of ubiquitin-specific protease-14 by Akt regulates the ubiquitin-proteasome system. Elife 2015, 4, e10510. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Cho, J.; Kim, E.E.; Song, E.J. Deubiquitinating Enzymes: A Critical Regulator of Mitosis. Int. J. Mol. Sci. 2019, 20, 5997. [Google Scholar] [CrossRef]
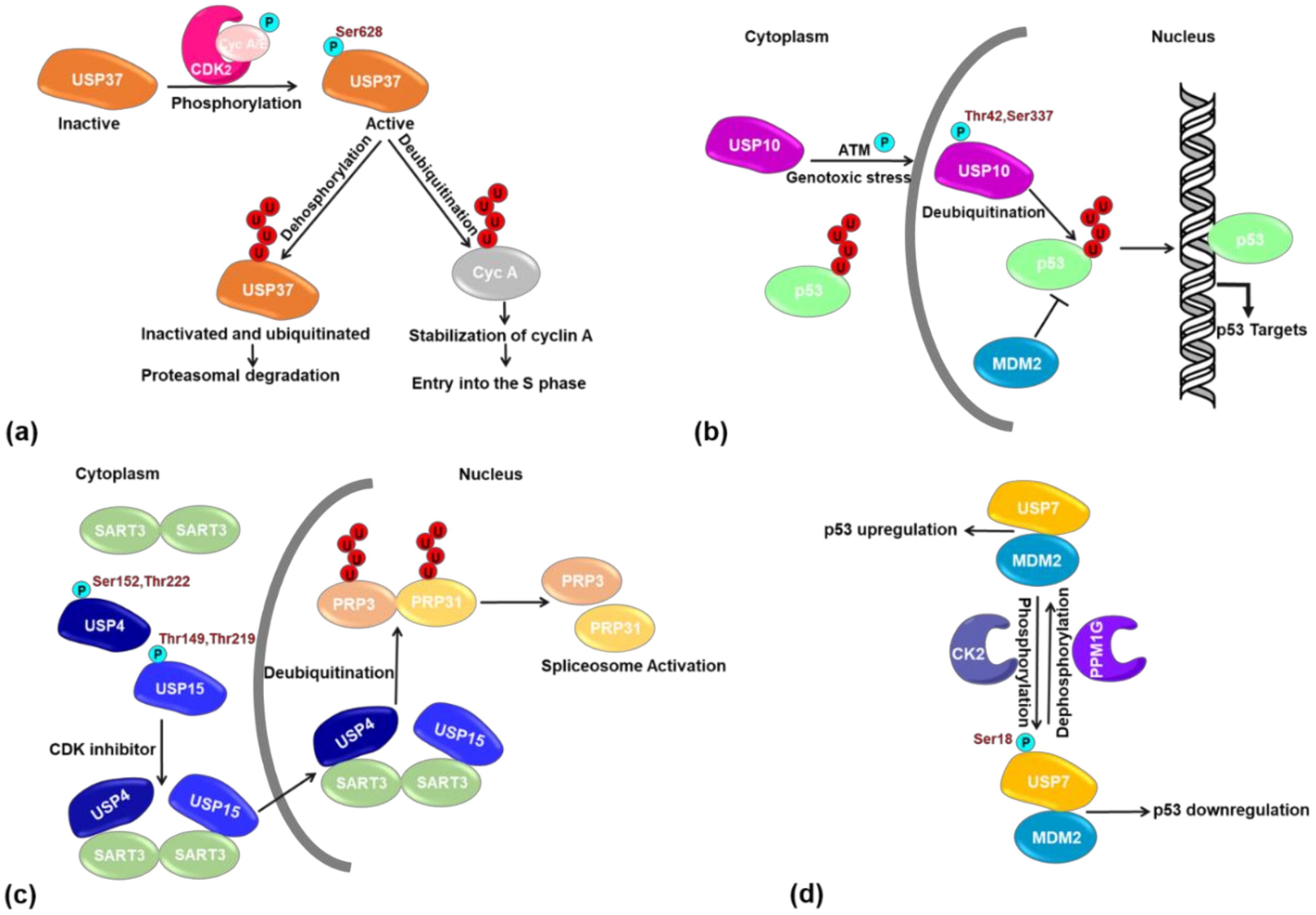
- Huang, X.; Summers, M.K.; Pham, V.; Lill, J.R.; Liu, J.; Lee, G.; Kirkpatrick, D.S.; Jackson, P.K.; Fang, G.; Dixit, V.M. Deubiquitinase USP37 is activated by CDK2 to antagonize APC(CDH1) and promote S phase entry. Mol. Cell 2011, 42, 511–523. [Google Scholar] [CrossRef]
- Stegmeier, F.; Rape, M.; Draviam, V.M.; Nalepa, G.; Sowa, M.E.; Ang, X.L.; McDonald, E.R., 3rd; Li, M.Z.; Hannon, G.J.; Sorger, P.K.; et al. Anaphase initiation is regulated by antagonistic ubiquitination and deubiquitination activities. Nature 2007, 446, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Suresh, B.; Ramakrishna, S.; Lee, H.J.; Choi, J.H.; Kim, J.Y.; Ahn, W.S.; Baek, K.H. K48- and K63-linked polyubiquitination of deubiquitinating enzyme USP44. Cell Biol. Int. 2010, 34, 799–808. [Google Scholar] [CrossRef]
- Mizuno, E.; Kitamura, N.; Komada, M. 14-3-3-dependent inhibition of the deubiquitinating activity of UBPY and its cancellation in the M phase. Exp. Cell Res. 2007, 313, 3624–3634. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.; Jentsch, S. Final stages of cytokinesis and midbody ring formation are controlled by BRUCE. Cell 2008, 132, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Mukai, A.; Mizuno, E.; Kobayashi, K.; Matsumoto, M.; Nakayama, K.I.; Kitamura, N.; Komada, M. Dynamic regulation of ubiquitylation and deubiquitylation at the central spindle during cytokinesis. J. Cell Sci. 2008, 121, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- Meijer, I.M.; Kerperien, J.; Sotoca, A.M.; van Zoelen, E.J.; van Leeuwen, J.E. The Usp8 deubiquitination enzyme is post-translationally modified by tyrosine and serine phosphorylation. Cell Signal. 2013, 25, 919–930. [Google Scholar] [CrossRef]
- Kasahara, K.; Aoki, H.; Kiyono, T.; Wang, S.; Kagiwada, H.; Yuge, M.; Tanaka, T.; Nishimura, Y.; Mizoguchi, A.; Goshima, N.; et al. EGF receptor kinase suppresses ciliogenesis through activation of USP8 deubiquitinase. Nat. Commun. 2018, 9, 758. [Google Scholar] [CrossRef]
- Naik, E.; Dixit, V.M. Usp9X Is Required for Lymphocyte Activation and Homeostasis through Its Control of ZAP70 Ubiquitination and PKCbeta Kinase Activity. J. Immunol. 2016, 196, 3438–3451. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Phung, Q.; Chan, S.; Chaudhari, R.; Quan, C.; O’Rourke, K.M.; Eby, M.; Pietras, E.; Cheng, G.; Bazan, J.F.; et al. DUBA: A deubiquitinase that regulates type I interferon production. Science 2007, 318, 1628–1632. [Google Scholar] [CrossRef]
- Huang, O.W.; Ma, X.; Yin, J.; Flinders, J.; Maurer, T.; Kayagaki, N.; Phung, Q.; Bosanac, I.; Arnott, D.; Dixit, V.M.; et al. Phosphorylation-dependent activity of the deubiquitinase DUBA. Nat. Struct. Mol. Biol. 2012, 19, 171–175. [Google Scholar] [CrossRef]
- Kabra, A.; Rumpa, E.; Li, Y. Modulation of conformational equilibrium by phosphorylation underlies the activation of deubiquitinase A. J. Biol. Chem. 2020, 295, 3945–3951. [Google Scholar] [CrossRef]
- Reiley, W.; Zhang, M.; Wu, X.; Granger, E.; Sun, S.C. Regulation of the deubiquitinating enzyme CYLD by IkappaB kinase gamma-dependent phosphorylation. Mol. Cell. Biol. 2005, 25, 3886–3895. [Google Scholar] [CrossRef]
- Douglas, T.; Saleh, M. Post-translational Modification of OTULIN Regulates Ubiquitin Dynamics and Cell Death. Cell Rep. 2019, 29, 3652–3663.e5. [Google Scholar] [CrossRef]
- Xu, X.; Kalac, M.; Markson, M.; Chan, M.; Brody, J.D.; Bhagat, G.; Ang, R.L.; Legarda, D.; Justus, S.J.; Liu, F.; et al. Reversal of CYLD phosphorylation as a novel therapeutic approach for adult T-cell leukemia/lymphoma (ATLL). Cell Death Dis. 2020, 11, 94. [Google Scholar] [CrossRef]
- Hutti, J.E.; Turk, B.E.; Asara, J.M.; Ma, A.; Cantley, L.C.; Abbott, D.W. IkappaB kinase beta phosphorylates the K63 deubiquitinase A20 to cause feedback inhibition of the NF-kappaB pathway. Mol. Cell Biol. 2007, 27, 7451–7461. [Google Scholar] [CrossRef] [PubMed]
- Zammit, N.W.; Siggs, O.M.; Gray, P.E.; Horikawa, K.; Langley, D.B.; Walters, S.N.; Daley, S.R.; Loetsch, C.; Warren, J.; Yap, J.Y.; et al. Denisovan, modern human and mouse TNFAIP3 alleles tune A20 phosphorylation and immunity. Nat. Immunol. 2019, 20, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Meray, R.K.; Lansbury, P.T., Jr. Reversible monoubiquitination regulates the Parkinson disease-associated ubiquitin hydrolase UCH-L1. J. Biol. Chem. 2007, 282, 10567–10575. [Google Scholar] [CrossRef]
- Leroy, E.; Boyer, R.; Auburger, G.; Leube, B.; Ulm, G.; Mezey, E.; Harta, G.; Brownstein, M.J.; Jonnalagada, S.; Chernova, T.; et al. The ubiquitin pathway in Parkinson’s disease. Nature 1998, 395, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.H.; Maraganore, D.M.; McDonnell, S.K.; Rocca, W.A. Incidence and distribution of parkinsonism in Olmsted County, Minnesota, 1976–1990. Neurology 1999, 52, 1214–1220. [Google Scholar] [CrossRef]
- Todi, S.V.; Winborn, B.J.; Scaglione, K.M.; Blount, J.R.; Travis, S.M.; Paulson, H.L. Ubiquitination directly enhances activity of the deubiquitinating enzyme ataxin-3. EMBO J. 2009, 28, 372–382. [Google Scholar] [CrossRef]
- Seki, T.; Gong, L.; Williams, A.J.; Sakai, N.; Todi, S.V.; Paulson, H.L. JosD1, a membrane-targeted deubiquitinating enzyme, is activated by ubiquitination and regulates membrane dynamics, cell motility, and endocytosis. J. Biol. Chem. 2013, 288, 17145–17155. [Google Scholar] [CrossRef]
- Meulmeester, E.; Kunze, M.; Hsiao, H.H.; Urlaub, H.; Melchior, F. Mechanism and consequences for paralog-specific sumoylation of ubiquitin-specific protease 25. Mol. Cell. 2008, 30, 610–619. [Google Scholar] [CrossRef]
- Zhen, Y.; Knobel, P.A.; Stracker, T.H.; Reverter, D. Regulation of USP28 deubiquitinating activity by SUMO conjugation. J. Biol. Chem. 2014, 289, 34838–34850. [Google Scholar] [CrossRef]
- Denuc, A.; Bosch-Comas, A.; Gonzalez-Duarte, R.; Marfany, G. The UBA-UIM domains of the USP25 regulate the enzyme ubiquitination state and modulate substrate recognition. PLoS ONE 2009, 4, e5571. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sureda-Gomez, M.; Zhen, Y.; Amador, V.; Reverter, D. A quaternary tetramer assembly inhibits the deubiquitinating activity of USP25. Nat. Commun. 2018, 9, 4973. [Google Scholar] [CrossRef]
- Gersch, M.; Wagstaff, J.L.; Toms, A.V.; Graves, B.; Freund, S.M.V.; Komander, D. Distinct USP25 and USP28 Oligomerization States Regulate Deubiquitinating Activity. Mol. Cell 2019, 74, 436–451.e7. [Google Scholar] [CrossRef] [PubMed]
- Sauer, F.; Klemm, T.; Kollampally, R.B.; Tessmer, I.; Nair, R.K.; Popov, N.; Kisker, C. Differential Oligomerization of the Deubiquitinases USP25 and USP28 Regulates Their Activities. Mol. Cell 2019, 74, 421–435.e10. [Google Scholar] [CrossRef] [PubMed]
- Cotto-Rios, X.M.; Bekes, M.; Chapman, J.; Ueberheide, B.; Huang, T.T. Deubiquitinases as a signaling target of oxidative stress. Cell Rep. 2012, 2, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Kulathu, Y.; Garcia, F.J.; Mevissen, T.E.; Busch, M.; Arnaudo, N.; Carroll, K.S.; Barford, D.; Komander, D. Regulation of A20 and other OTU deubiquitinases by reversible oxidation. Nat. Commun. 2013, 4, 1569. [Google Scholar] [CrossRef]
- Lee, J.G.; Baek, K.; Soetandyo, N.; Ye, Y. Reversible inactivation of deubiquitinases by reactive oxygen species in vitro and in cells. Nat. Commun. 2013, 4, 1568. [Google Scholar] [CrossRef]
- Urbe, S.; Liu, H.; Hayes, S.D.; Heride, C.; Rigden, D.J.; Clague, M.J. Systematic survey of deubiquitinase localization identifies USP21 as a regulator of centrosome- and microtubule-associated functions. Mol. Biol. Cell 2012, 23, 1095–1103. [Google Scholar] [CrossRef]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 regulates p53 localization and stability by deubiquitinating p53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef]
- Mueller, T.; Breuer, P.; Schmitt, I.; Walter, J.; Evert, B.O.; Wullner, U. CK2-dependent phosphorylation determines cellular localization and stability of ataxin-3. Hum. Mol. Genet. 2009, 18, 3334–3343. [Google Scholar] [CrossRef]
- Pastori, V.; Sangalli, E.; Coccetti, P.; Pozzi, C.; Nonnis, S.; Tedeschi, G.; Fusi, P. CK2 and GSK3 phosphorylation on S29 controls wild-type ATXN3 nuclear uptake. Biochim. Biophys. Acta 2010, 1802, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Marcelo, A.; Brito, F.; Carmo-Silva, S.; Matos, C.A.; Alves-Cruzeiro, J.; Vasconcelos-Ferreira, A.; Koppenol, R.; Mendonca, L.; de Almeida, L.P.; Nobrega, C. Cordycepin activates autophagy through AMPK phosphorylation to reduce abnormalities in Machado-Joseph disease models. Hum. Mol. Genet. 2019, 28, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Herhaus, L.; Perez-Oliva, A.B.; Cozza, G.; Gourlay, R.; Weidlich, S.; Campbell, D.G.; Pinna, L.A.; Sapkota, G.P. Casein kinase 2 (CK2) phosphorylates the deubiquitylase OTUB1 at Ser16 to trigger its nuclear localization. Sci. Signal 2015, 8, ra35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Drabsch, Y.; Gao, R.; Snaar-Jagalska, B.E.; Mickanin, C.; Huang, H.; Sheppard, K.A.; Porter, J.A.; Lu, C.X.; et al. USP4 is regulated by AKT phosphorylation and directly deubiquitylates TGF-beta type I receptor. Nat. Cell Biol. 2012, 14, 717–726. [Google Scholar] [CrossRef]
- Row, P.E.; Prior, I.A.; McCullough, J.; Clague, M.J.; Urbe, S. The ubiquitin isopeptidase UBPY regulates endosomal ubiquitin dynamics and is essential for receptor down-regulation. J. Biol. Chem. 2006, 281, 12618–12624. [Google Scholar] [CrossRef]
- Blagoev, B.; Ong, S.E.; Kratchmarova, I.; Mann, M. Temporal analysis of phosphotyrosine-dependent signaling networks by quantitative proteomics. Nat. Biotechnol. 2004, 22, 1139–1145. [Google Scholar] [CrossRef]
- Ballif, B.A.; Cao, Z.; Schwartz, D.; Carraway, K.L., 3rd; Gygi, S.P. Identification of 14-3-3epsilon substrates from embryonic murine brain. J. Proteome. Res. 2006, 5, 2372–2379. [Google Scholar] [CrossRef]
- Liu, Z.; Meray, R.K.; Grammatopoulos, T.N.; Fredenburg, R.A.; Cookson, M.R.; Liu, Y.; Logan, T.; Lansbury, P.T., Jr. Membrane-associated farnesylated UCH-L1 promotes alpha-synuclein neurotoxicity and is a therapeutic target for Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2009, 106, 4635–4640. [Google Scholar] [CrossRef]
- Kobayashi, E.; Aga, M.; Kondo, S.; Whitehurst, C.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. C-Terminal Farnesylation of UCH-L1 Plays a Role in Transport of Epstein-Barr Virus Primary Oncoprotein LMP1 to Exosomes. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Bishop, P.; Rubin, P.; Thomson, A.R.; Rocca, D.; Henley, J.M. The ubiquitin C-terminal hydrolase L1 (UCH-L1) C terminus plays a key role in protein stability, but its farnesylation is not required for membrane association in primary neurons. J. Biol. Chem. 2014, 289, 36140–36149. [Google Scholar] [CrossRef]
- Akhavantabasi, S.; Akman, H.B.; Sapmaz, A.; Keller, J.; Petty, E.M.; Erson, A.E. USP32 is an active, membrane-bound ubiquitin protease overexpressed in breast cancers. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2010, 21, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Mevissen, T.E.; Hospenthal, M.K.; Geurink, P.P.; Elliott, P.R.; Akutsu, M.; Arnaudo, N.; Ekkebus, R.; Kulathu, Y.; Wauer, T.; El Oualid, F.; et al. OTU deubiquitinases reveal mechanisms of linkage specificity and enable ubiquitin chain restriction analysis. Cell 2013, 154, 169–184. [Google Scholar] [CrossRef] [PubMed]
- McCullough, J.; Row, P.E.; Lorenzo, O.; Doherty, M.; Beynon, R.; Clague, M.J.; Urbe, S. Activation of the endosome-associated ubiquitin isopeptidase AMSH by STAM, a component of the multivesicular body-sorting machinery. Curr. Biol. 2006, 16, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Row, P.E.; Liu, H.; Hayes, S.; Welchman, R.; Charalabous, P.; Hofmann, K.; Clague, M.J.; Sanderson, C.M.; Urbe, S. The MIT domain of UBPY constitutes a CHMP binding and endosomal localization signal required for efficient epidermal growth factor receptor degradation. J. Biol. Chem. 2007, 282, 30929–30937. [Google Scholar] [CrossRef] [PubMed]
- Cope, G.A.; Suh, G.S.; Aravind, L.; Schwarz, S.E.; Zipursky, S.L.; Koonin, E.V.; Deshaies, R.J. Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 2002, 298, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Aravind, L.; Oania, R.; McDonald, W.H.; Yates, J.R., 3rd; Koonin, E.V.; Deshaies, R.J. Role of Rpn11 metalloprotease in deubiquitination and degradation by the 26S proteasome. Science 2002, 298, 611–615. [Google Scholar] [CrossRef]
- Yao, T.; Song, L.; Jin, J.; Cai, Y.; Takahashi, H.; Swanson, S.K.; Washburn, M.P.; Florens, L.; Conaway, R.C.; Cohen, R.E.; et al. Distinct modes of regulation of the Uch37 deubiquitinating enzyme in the proteasome and in the Ino80 chromatin-remodeling complex. Mol. Cell 2008, 31, 909–917. [Google Scholar] [CrossRef]
- Garcia-Santisteban, I.; Zorroza, K.; Rodriguez, J.A. Two nuclear localization signals in USP1 mediate nuclear import of the USP1/UAF1 complex. PLoS ONE 2012, 7, e38570. [Google Scholar] [CrossRef]
- Olazabal-Herrero, A.; Garcia-Santisteban, I.; Rodriguez, J.A. Structure-function analysis of USP1: Insights into the role of Ser313 phosphorylation site and the effect of cancer-associated mutations on autocleavage. Mol. Cancer 2015, 14, 33. [Google Scholar] [CrossRef][Green Version]
- Villamil, M.A.; Liang, Q.; Chen, J.; Choi, Y.S.; Hou, S.; Lee, K.H.; Zhuang, Z. Serine phosphorylation is critical for the activation of ubiquitin-specific protease 1 and its interaction with WD40-repeat protein UAF1. Biochemistry 2012, 51, 9112–9123. [Google Scholar] [CrossRef]
- Cohn, M.A.; Kowal, P.; Yang, K.; Haas, W.; Huang, T.T.; Gygi, S.P.; D’Andrea, A.D. A UAF1-containing multisubunit protein complex regulates the Fanconi anemia pathway. Mol. Cell 2007, 28, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Moldovan, G.L.; Vinciguerra, P.; Murai, J.; Takeda, S.; D’Andrea, A.D. Regulation of the Fanconi anemia pathway by a SUMO-like delivery network. Genes Dev. 2011, 25, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Schoeffler, A.J.; Wickliffe, K.; Newton, K.; Starovasnik, M.A.; Dueber, E.C.; Harris, S.F. Structural Insights into WD-Repeat 48 Activation of Ubiquitin-Specific Protease 46. Structure 2015, 23, 2043–2054. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhang, T.; Wang, F.; Yang, J.; Ding, J. Structural insights into the activation of USP46 by WDR48 and WDR20. Cell Discov. 2019, 5, 34. [Google Scholar] [CrossRef]
- Li, H.; Lim, K.S.; Kim, H.; Hinds, T.R.; Jo, U.; Mao, H.; Weller, C.E.; Sun, J.; Chatterjee, C.; D’Andrea, A.D.; et al. Allosteric Activation of Ubiquitin-Specific Proteases by beta-Propeller Proteins UAF1 and WDR20. Mol. Cell 2016, 63, 249–260. [Google Scholar] [CrossRef]
- Cohn, M.A.; Kee, Y.; Haas, W.; Gygi, S.P.; D’Andrea, A.D. UAF1 is a subunit of multiple deubiquitinating enzyme complexes. J. Biol. Chem. 2009, 284, 5343–5351. [Google Scholar] [CrossRef]
- Xiao, N.; Li, H.; Luo, J.; Wang, R.; Chen, H.; Chen, J.; Wang, P. Ubiquitin-specific protease 4 (USP4) targets TRAF2 and TRAF6 for deubiquitination and inhibits TNFalpha-induced cancer cell migration. Biochem. J. 2012, 441, 979–986. [Google Scholar] [CrossRef]
- Yun, S.I.; Kim, H.H.; Yoon, J.H.; Park, W.S.; Hahn, M.J.; Kim, H.C.; Chung, C.H.; Kim, K.K. Ubiquitin specific protease 4 positively regulates the WNT/beta-catenin signaling in colorectal cancer. Mol. Oncol. 2015, 9, 1834–1851. [Google Scholar] [CrossRef]
- Zhang, X.; Berger, F.G.; Yang, J.; Lu, X. USP4 inhibits p53 through deubiquitinating and stabilizing ARF-BP1. EMBO J. 2011, 30, 2177–2189. [Google Scholar] [CrossRef]
- Milojevic, T.; Reiterer, V.; Stefan, E.; Korkhov, V.M.; Dorostkar, M.M.; Ducza, E.; Ogris, E.; Boehm, S.; Freissmuth, M.; Nanoff, C. The ubiquitin-specific protease Usp4 regulates the cell surface level of the A2A receptor. Mol. Pharmacol. 2006, 69, 1083–1094. [Google Scholar] [CrossRef]
- Wada, K.; Tanji, K.; Kamitani, T. Oncogenic protein UnpEL/Usp4 deubiquitinates Ro52 by its isopeptidase activity. Biochem. Biophys. Res. Commun. 2006, 339, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Torre, S.; Polyak, M.J.; Langlais, D.; Fodil, N.; Kennedy, J.M.; Radovanovic, I.; Berghout, J.; Leiva-Torres, G.A.; Krawczyk, C.M.; Ilangumaran, S.; et al. USP15 regulates type I interferon response and is required for pathogenesis of neuroinflammation. Nat. Immunol. 2017, 18, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Sun, S.C. Deubiquitinases as pivotal regulators of T cell functions. Front. Med. 2018, 12, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, N.F.; Tian, W.; Wu, T.; Sun, Z.; Lau, A.; Chapman, E.; Fang, D.; Zhang, D.D. USP15 negatively regulates Nrf2 through deubiquitination of Keap1. Mol. Cell 2013, 51, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Herhaus, L.; Al-Salihi, M.A.; Dingwell, K.S.; Cummins, T.D.; Wasmus, L.; Vogt, J.; Ewan, R.; Bruce, D.; Macartney, T.; Weidlich, S.; et al. USP15 targets ALK3/BMPR1A for deubiquitylation to enhance bone morphogenetic protein signalling. Open Biol. 2014, 4, 140065. [Google Scholar] [CrossRef]
- Huang, X.; Langelotz, C.; Hetfeld-Pechoc, B.K.; Schwenk, W.; Dubiel, W. The COP9 signalosome mediates beta-catenin degradation by deneddylation and blocks adenomatous polyposis coli destruction via USP15. J. Mol. Biol. 2009, 391, 691–702. [Google Scholar] [CrossRef]
- Eichhorn, P.J.; Rodon, L.; Gonzalez-Junca, A.; Dirac, A.; Gili, M.; Martinez-Saez, E.; Aura, C.; Barba, I.; Peg, V.; Prat, A.; et al. USP15 stabilizes TGF-beta receptor I and promotes oncogenesis through the activation of TGF-beta signaling in glioblastoma. Nat. Med. 2012, 18, 429–435. [Google Scholar] [CrossRef]
- Song, E.J.; Werner, S.L.; Neubauer, J.; Stegmeier, F.; Aspden, J.; Rio, D.; Harper, J.W.; Elledge, S.J.; Kirschner, M.W.; Rape, M. The Prp19 complex and the Usp4Sart3 deubiquitinating enzyme control reversible ubiquitination at the spliceosome. Genes Dev. 2010, 24, 1434–1447. [Google Scholar] [CrossRef]
- Das, T.; Park, J.K.; Park, J.; Kim, E.; Rape, M.; Kim, E.E.; Song, E.J. USP15 regulates dynamic protein-protein interactions of the spliceosome through deubiquitination of PRP31. Nucleic Acids Res. 2017, 45, 5010–5011. [Google Scholar] [CrossRef][Green Version]
- Park, J.K.; Das, T.; Song, E.J.; Kim, E.E. Structural basis for recruiting and shuttling of the spliceosomal deubiquitinase USP4 by SART3. Nucleic Acids Res. 2016, 44, 5424–5437. [Google Scholar] [CrossRef]
- Soboleva, T.A.; Jans, D.A.; Johnson-Saliba, M.; Baker, R.T. Nuclear-cytoplasmic shuttling of the oncogenic mouse UNP/USP4 deubiquitylating enzyme. J. Biol. Chem. 2005, 280, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Kim, E.E.; Song, E.J. Phosphorylation of USP15 and USP4 Regulates Localization and Spliceosomal Deubiquitination. J. Mol. Biol. 2019, 431, 3900–3912. [Google Scholar] [CrossRef] [PubMed]
- Vlasschaert, C.; Xia, X.; Coulombe, J.; Gray, D.A. Evolution of the highly networked deubiquitinating enzymes USP4, USP15, and USP11. BMC Evol. Biol. 2015, 15, 230. [Google Scholar] [CrossRef] [PubMed]
- Fei, E.; Jia, N.; Zhang, T.; Ma, X.; Wang, H.; Liu, C.; Zhang, W.; Ding, L.; Nukina, N.; Wang, G. Phosphorylation of ataxin-3 by glycogen synthase kinase 3beta at serine 256 regulates the aggregation of ataxin-3. Biochem. Biophys. Res. Commun. 2007, 357, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Khoronenkova, S.V.; Dianova, I.I.; Ternette, N.; Kessler, B.M.; Parsons, J.L.; Dianov, G.L. ATM-dependent downregulation of USP7/HAUSP by PPM1G activates p53 response to DNA damage. Mol. Cell 2012, 45, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Montalvan, A.; Bouwmeester, T.; Joberty, G.; Mader, R.; Mahnke, M.; Pierrat, B.; Schlaeppi, J.M.; Worpenberg, S.; Gerhartz, B. Biochemical characterization of USP7 reveals post-translational modification sites and structural requirements for substrate processing and subcellular localization. FEBS J. 2007, 274, 4256–4270. [Google Scholar] [CrossRef]
- Boutell, C.; Canning, M.; Orr, A.; Everett, R.D. Reciprocal activities between herpes simplex virus type 1 regulatory protein ICP0, a ubiquitin E3 ligase, and ubiquitin-specific protease USP7. J. Virol. 2005, 79, 12342–12354. [Google Scholar] [CrossRef]
- Cai, J.; Crotty, T.M.; Reichert, E.; Carraway, K.L., 3rd; Stafforini, D.M.; Topham, M.K. Diacylglycerol kinase delta and protein kinase C(alpha) modulate epidermal growth factor receptor abundance and degradation through ubiquitin-specific protease 8. J. Biol. Chem. 2010, 285, 6952–6959. [Google Scholar] [CrossRef]
- Faronato, M.; Patel, V.; Darling, S.; Dearden, L.; Clague, M.J.; Urbe, S.; Coulson, J.M. The deubiquitylase USP15 stabilizes newly synthesized REST and rescues its expression at mitotic exit. Cell Cycle 2013, 12, 1964–1977. [Google Scholar] [CrossRef]
- Fielding, A.B.; Concannon, M.; Darling, S.; Rusilowicz-Jones, E.V.; Sacco, J.J.; Prior, I.A.; Clague, M.J.; Urbe, S.; Coulson, J.M. The deubiquitylase USP15 regulates topoisomerase II alpha to maintain genome integrity. Oncogene 2018, 37, 2326–2342. [Google Scholar] [CrossRef]
- Cao, Z.; Wu, X.; Yen, L.; Sweeney, C.; Carraway, K.L., 3rd. Neuregulin-induced ErbB3 downregulation is mediated by a protein stability cascade involving the E3 ubiquitin ligase Nrdp1. Mol. Cell. Biol. 2007, 27, 2180–2188. [Google Scholar] [CrossRef] [PubMed]
- Cholay, M.; Reverdy, C.; Benarous, R.; Colland, F.; Daviet, L. Functional interaction between the ubiquitin-specific protease 25 and the SYK tyrosine kinase. Exp. Cell Res. 2010, 316, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zaugg, K.; Mak, T.W.; Elledge, S.J. A role for the deubiquitinating enzyme USP28 in control of the DNA-damage response. Cell 2006, 126, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Hutti, J.E.; Shen, R.R.; Abbott, D.W.; Zhou, A.Y.; Sprott, K.M.; Asara, J.M.; Hahn, W.C.; Cantley, L.C. Phosphorylation of the tumor suppressor CYLD by the breast cancer oncogene IKKepsilon promotes cell transformation. Mol. Cell 2009, 34, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Edelmann, M.J.; Kramer, H.B.; Altun, M.; Kessler, B.M. Post-translational modification of the deubiquitinating enzyme otubain 1 modulates active RhoA levels and susceptibility to Yersinia invasion. FEBS J. 2010, 277, 2515–2530. [Google Scholar] [CrossRef]
- Yu, H.; Pak, H.; Hammond-Martel, I.; Ghram, M.; Rodrigue, A.; Daou, S.; Barbour, H.; Corbeil, L.; Hebert, J.; Drobetsky, E.; et al. Tumor suppressor and deubiquitinase BAP1 promotes DNA double-strand break repair. Proc. Natl. Acad. Sci. USA 2014, 111, 285–290. [Google Scholar] [CrossRef]
- Shen, C.; Ye, Y.; Robertson, S.E.; Lau, A.W.; Mak, D.O.; Chou, M.M. Calcium/calmodulin regulates ubiquitination of the ubiquitin-specific protease TRE17/USP6. J. Biol. Chem. 2005, 280, 35967–35973. [Google Scholar] [CrossRef]
- Bingol, B.; Tea, J.S.; Phu, L.; Reichelt, M.; Bakalarski, C.E.; Song, Q.; Foreman, O.; Kirkpatrick, D.S.; Sheng, M. The mitochondrial deubiquitinase USP30 opposes parkin-mediated mitophagy. Nature 2014, 510, 370–375. [Google Scholar] [CrossRef]
- Todi, S.V.; Scaglione, K.M.; Blount, J.R.; Basrur, V.; Conlon, K.P.; Pastore, A.; Elenitoba-Johnson, K.; Paulson, H.L. Activity and cellular functions of the deubiquitinating enzyme and polyglutamine disease protein ataxin-3 are regulated by ubiquitination at lysine 117. J. Biol. Chem. 2010, 285, 39303–39313. [Google Scholar] [CrossRef]
- Mashtalir, N.; Daou, S.; Barbour, H.; Sen, N.N.; Gagnon, J.; Hammond-Martel, I.; Dar, H.H.; Therrien, M.; Affar el, B. Autodeubiquitination protects the tumor suppressor BAP1 from cytoplasmic sequestration mediated by the atypical ubiquitin ligase UBE2O. Mol. Cell 2014, 54, 392–406. [Google Scholar] [CrossRef]
- Kobayashi, T.; Masoumi, K.C.; Massoumi, R. Deubiquitinating activity of CYLD is impaired by SUMOylation in neuroblastoma cells. Oncogene 2015, 34, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.F.; Liao, S.S.; Luo, Y.Y.; Tang, J.G.; Wang, J.L.; Lei, L.F.; Chi, J.W.; Du, J.; Jiang, H.; Xia, K.; et al. SUMO-1 modification on K166 of polyQ-expanded ataxin-3 strengthens its stability and increases its cytotoxicity. PLoS ONE 2013, 8, e54214. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.N.; Hart, G.W. Cytosolic O-glycosylation is abundant in nerve terminals. J. Neurochem. 2001, 79, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, P.; Wang, X.; Linder, S. Deubiquitinase inhibition as a cancer therapeutic strategy. Pharm. 2015, 147, 32–54. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.P.; Chen, J.; Tse, W.K.F. Role of Deubiquitinases in Human Cancers: Potential Targeted Therapy. Int. J. Mol. Sci. 2020, 21, 2548. [Google Scholar] [CrossRef]
- Ruan, J.; Schluter, D.; Wang, X. Deubiquitinating enzymes (DUBs): DoUBle-edged swords in CNS autoimmunity. J. Neuroinflamm. 2020, 17, 102. [Google Scholar] [CrossRef]
- Ebner, P.; Versteeg, G.A.; Ikeda, F. Ubiquitin enzymes in the regulation of immune responses. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 425–460. [Google Scholar] [CrossRef]
- Wang, B.; Cai, W.; Ai, D.; Zhang, X.; Yao, L. The Role of Deubiquitinases in Vascular Diseases. J. Cardiovasc. Transl. Res. 2020, 13, 131–141. [Google Scholar] [CrossRef]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef]
- Reyes-Turcu, F.E.; Ventii, K.H.; Wilkinson, K.D. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu. Rev. Biochem. 2009, 78, 363–397. [Google Scholar] [CrossRef]
- Heideker, J.; Wertz, I.E. DUBs, the regulation of cell identity and disease. Biochem. J. 2015, 467, 191. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Cho, S.; Park, B.C.; Lee, D.H. Oxidative stress-enhanced SUMOylation and aggregation of ataxin-1: Implication of JNK pathway. Biochem. Biophys. Res. Commun. 2010, 393, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.J.; Magin, R.S.; Liu, X.; Doherty, L.M.; Buhrlage, S.J. Advances in Discovering Deubiquitinating Enzyme (DUB) Inhibitors. J. Med. Chem. 2020, 63, 2731–2750. [Google Scholar] [CrossRef] [PubMed]
- Wertz, I.E.; Wang, X. From Discovery to Bedside: Targeting the Ubiquitin System. Cell Chem. Biol. 2019, 26, 156–177. [Google Scholar] [CrossRef]
- Wu, H.Q.; Baker, D.; Ovaa, H. Small molecules that target the ubiquitin system. Biochem. Soc. Trans. 2020. [Google Scholar] [CrossRef]
- Wertz, I.E.; Murray, J.M. Structurally-defined deubiquitinase inhibitors provide opportunities to investigate disease mechanisms. Drug Discov. Today Technol. 2019, 31, 109–123. [Google Scholar] [CrossRef]
- Su, M.G.; Weng, J.T.; Hsu, J.B.; Huang, K.Y.; Chi, Y.H.; Lee, T.Y. Investigation and identification of functional post-translational modification sites associated with drug binding and protein-protein interactions. BMC Syst. Biol. 2017, 11, 132. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DUBs | Modified Sites | Effect in Cells | References |
|---|---|---|---|
| Phosphorylation | |||
| USP1 | Ser313 | Promote DUB catalytic activity by enhancing the interaction with cofactor UFA1 | [70] |
| USP4 | Ser445 | Alter the subcellular localization from the nucleus to cytoplasm | [54] |
| USP7 | Ser18 | Stabilization of USP7 for MDM2 deubiquitination | [95] |
| Ser963 | Unknown (possibly protein-protein interaction) | [96] | |
| USP8 | Ser680 | Inhibit catalytic activity by promoting USP8 association with the 14-3-3 family proteins | [20] |
| Tyr717, Tyr810 | Elevates activity leading to inhibition of ciliogenesis | [24] | |
| Thr907 | Increase Stability | [101] | |
| USP9X | Ser1600 | Enhance catalytic activity | [25] |
| USP10 | Thr42, Ser337 | Increase stability by inducing nuclear translocation | [49] |
| USP14 | Ser432 | Increase catalytic activity | [15] |
| USP15 | Thr149, Thr219 | Regulate interaction, localization and DUB activity towards its substrate PRP31 | [92] |
| Ser229 | Abrogate USP15 function in maintaining TOP2A mediated genomic stability | [100] | |
| USP25 | Tyr740 | Decrease USP25 cellular level and stability | [102] |
| USP28 | Ser67, Ser714 | Regulate the complex-formation with the DNA checkpoint proteins | [103] |
| USP37 | Ser628 | Enhance catalytic activity in G1/S during cell cycle. | [17] |
| USP44 | Unknown | Activation of USP44 | [18] |
| CYLD | Ser418 | Decrease DUB activity and induce IKKε-mediated cell transformation | [104] |
| OTUD5 | Ser177 | Required for activation of the enzyme | [27] |
| A20 | Ser381 | Increase the activity of A20 to inhibit NF-κB signaling pathway | [32] |
| ATX3 | Ser340, Ser352 | Increase nuclear localization and aggregation | [50] |
| Ser129 | Promote nuclear uptake | [51] | |
| OTUB1 | Ser16, Ser18, Tyr26 | Increase stability and protein–protein interaction | [105] |
| BAP1 | Thr273, Ser276, Ser592 | Promote DNA repair and cellular recovery from DNA damage | [106] |
| Ubiquitination | |||
| USP6 | Mono-ubiquitination | Promotes its own deubiquitination | [107] |
| USP7 | Lys869 | Decrease stability and modulate protein-protein interaction | [96] |
| USP25 | Mono-ubiquitination at Lys99 | Enhance catalytic activity and substrate recognition | [41] |
| USP30 | Fingers subdomain of the catalytic domain | Induce proteasomal degradation | [108] |
| USP44 | Unknown | Diminish stability by inducing proteasomal degradation. | [19] |
| ATXN3 | Lys117 | Enhance catalytic activity | [109] |
| JosD1 | Unknown | Enhance catalytic activity and regulates membrane dynamics, cell motility, and endocytosis | [38] |
| UCHL1 | Lys4, Lys65, Lys71, Lys157 | Inhibit enzyme activity | [34] |
| BAP1 | Multiple mono-ubiquitination within the NLS region (residues 699–729) | Prevent cytoplasmic accumulation by auto deubiquitination | [110] |
| SUMOylation | |||
| USP25 | Lys99, Lys 141 | Inhibit catalytic activity by decreasing chain hydrolysis | [39] |
| USP28 | Unknown | Negatively regulate the deubiquitinating activity | [40] |
| CLYD | Unknown | Inhibit activity against TRAF2 and TRAF6 and facilitates NFκB signaling | [111] |
| ATXN3 | Lys166 | Enhance stability | [112] |
| Others | |||
| USP1 | Oxidation | Reduce catalytic activity | [45] |
| USP32 | Lipid modification | Association with intracellular membranes | [11] |
| UCHL1 | Farnesylation | Promote intracellular membrane association and increased α-synuclein accumulation | [58] |
| UCHL1 | O-glycosylation | Regulate synaptosome proteins functions playing a vital role in neurodegenerative disease. | [113] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, T.; Shin, S.C.; Song, E.J.; Kim, E.E. Regulation of Deubiquitinating Enzymes by Post-Translational Modifications. Int. J. Mol. Sci. 2020, 21, 4028. https://doi.org/10.3390/ijms21114028
Das T, Shin SC, Song EJ, Kim EE. Regulation of Deubiquitinating Enzymes by Post-Translational Modifications. International Journal of Molecular Sciences. 2020; 21(11):4028. https://doi.org/10.3390/ijms21114028
Chicago/Turabian StyleDas, Tanuza, Sang Chul Shin, Eun Joo Song, and Eunice EunKyeong Kim. 2020. "Regulation of Deubiquitinating Enzymes by Post-Translational Modifications" International Journal of Molecular Sciences 21, no. 11: 4028. https://doi.org/10.3390/ijms21114028
APA StyleDas, T., Shin, S. C., Song, E. J., & Kim, E. E. (2020). Regulation of Deubiquitinating Enzymes by Post-Translational Modifications. International Journal of Molecular Sciences, 21(11), 4028. https://doi.org/10.3390/ijms21114028