Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice

 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
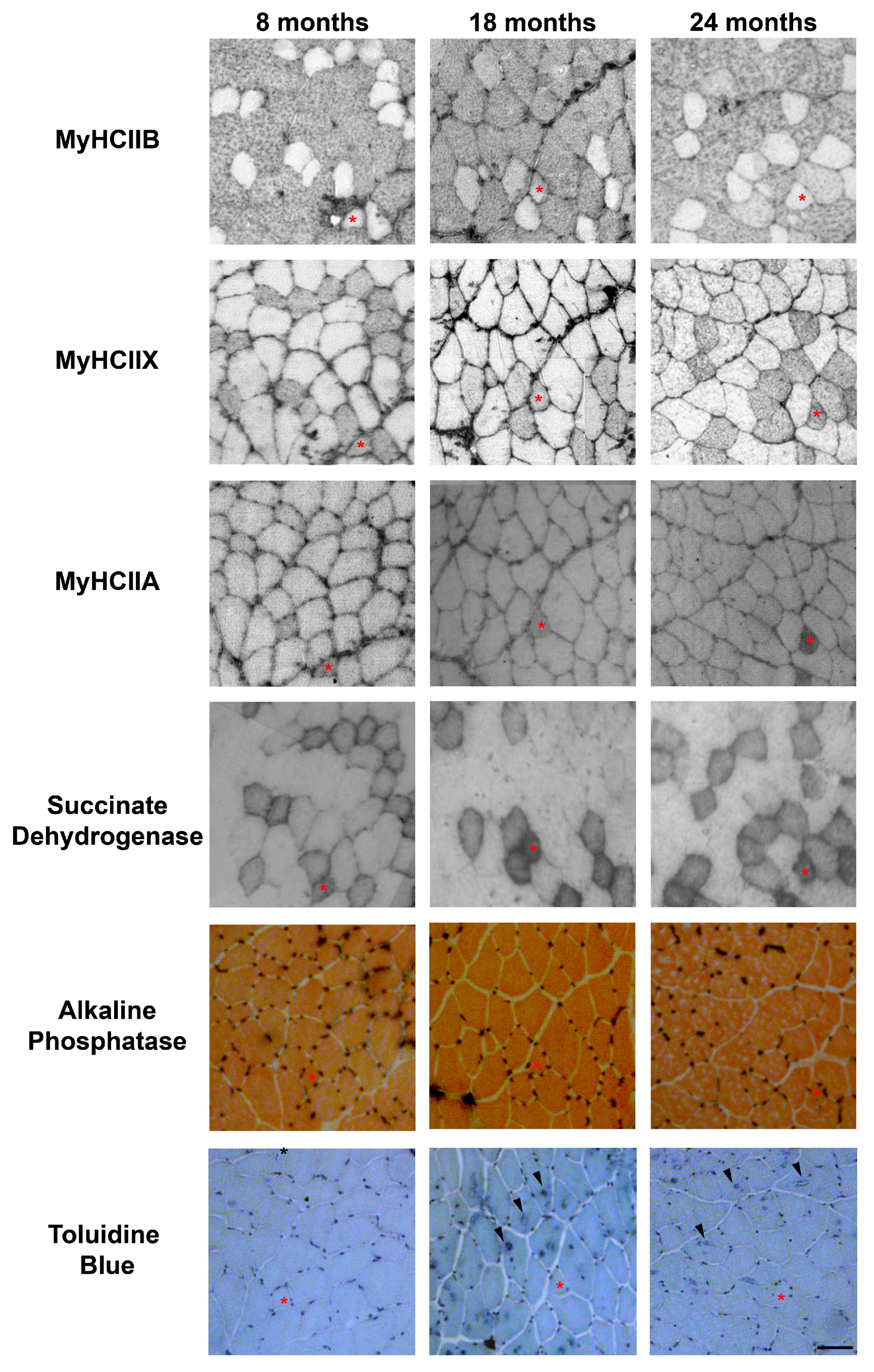
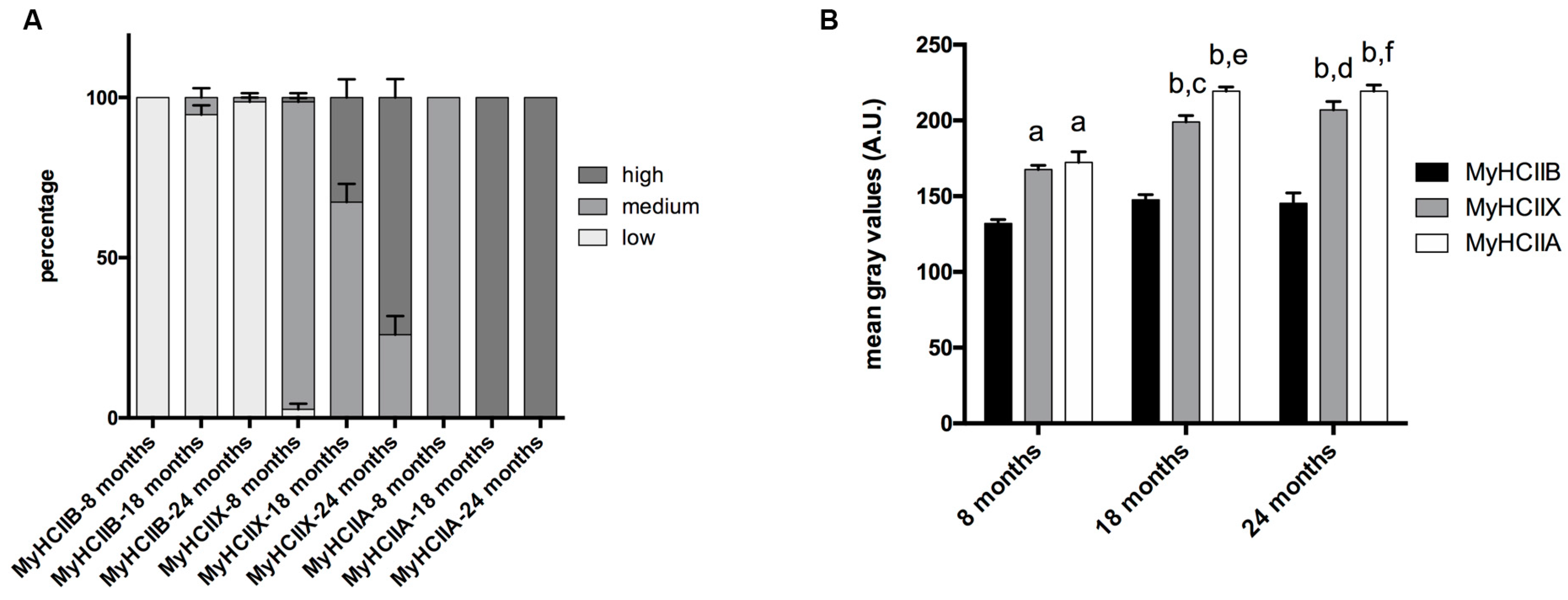
2.1. Fiber-Type Composition in Aging Tibialis Anterior Muscle
2.2. Fiber-Type CSA in Aging Tibialis Anterior Muscle
2.3. Fiber Type and Measurement of SDH Activity
2.4. Capillarization
2.5. Tubular Aggregates
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Antibodies
4.3. Cryostat Sectioning
4.4. Immunohistochemistry Reactions
4.5. SDH Staining
4.6. Alkaline Phosphatase Staining
4.7. Toluidine Blue Staining
4.8. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Schiaffino, S.; Reggiani, C.; Murgia, M. Fiber type diversity in skeletal muscle explored by mass spectrometry-based single fiber proteomics. Histol. Histopathol. 2019, 35, 239–246. [Google Scholar] [PubMed]
- Burke, R.E.; Levine, D.N.; Zajac, F.E., 3rd. Mammalian motor units: Physiological-histochemical correlation in three types in cat gastrocnemius. Science 1971, 174, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed]
- Ortenblad, N.; Nielsen, J.; Boushel, R.; Soderlund, K.; Saltin, B.; Holmberg, H.C. The muscle fiber profiles, mitochondrial content, and enzyme activities of the exceptionally well-trained arm and leg muscles of elite cross-country skiers. Front. Physiol. 2018, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- Kent-Braun, J.A.; Fitts, R.H.; Christie, A. Skeletal muscle fatigue. Compr. Physiol. 2012, 2, 997–1044. [Google Scholar] [PubMed]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-related loss of muscle mass and function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Narici, M.V.; Maganaris, C.N.; Reeves, N.D.; Capodaglio, P. Effect of aging on human muscle architecture. J. Appl. Physiol. 2003, 95, 2229–2234. [Google Scholar] [CrossRef]
- Naro, F.; Venturelli, M.; Monaco, L.; Toniolo, L.; Muti, E.; Milanese, C.; Zhao, J.; Richardson, R.S.; Schena, F.; Reggiani, C. Skeletal muscle fiber size and gene expression in the oldest-old with differing degrees of mobility. Front. Physiol. 2019, 10, 313. [Google Scholar] [CrossRef]
- Delbono, O. Expression and regulation of excitation-contraction coupling proteins in aging skeletal muscle. Curr. Aging Sci. 2011, 4, 248–259. [Google Scholar] [CrossRef]
- Prior, S.J.; Ryan, A.S.; Blumenthal, J.B.; Watson, J.M.; Katzel, L.I.; Goldberg, A.P. Sarcopenia is associated with lower skeletal muscle capillarization and exercise capacity in older adults. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2016, 71, 1096–1101. [Google Scholar] [CrossRef]
- Vielhaber, S.; Schroder, R.; Winkler, K.; Weis, S.; Sailer, M.; Feistner, H.; Heinze, H.J.; Schroder, J.M.; Kunz, W.S. Defective mitochondrial oxidative phosphorylation in myopathies with tubular aggregates originating from sarcoplasmic reticulum. J. Neuropathol. Exp. Neurol. 2001, 60, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S. Tubular aggregates in skeletal muscle: Just a special type of protein aggregates? Neuromuscul. Disord. NMD 2012, 22, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, E.; Quarta, M.; Paolini, C.; Squecco, R.; Fusco, P.; Toniolo, L.; Blaauw, B.; Formoso, L.; Rossi, D.; Birkenmeier, C.; et al. Deletion of small ankyrin 1 (sank1) isoforms results in structural and functional alterations in aging skeletal muscle fibers. Am. J. Physiol. Cell Physiol. 2015, 308, C123–C138. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Standley, R.A.; Murach, K.A.; Raue, U.; Minchev, K.; Coen, P.M.; Newman, A.B.; Cummings, S.; Harris, T.; Kritchevsky, S.; et al. Improved single muscle fiber quality in the oldest-old. J. Appl. Physiol. 2016, 121, 878–884. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Flurkey, K.; Currer, J.M.; Harrison, D.E. Mouse models in aging research. In The Mouse in Biomedical Research, 2nd ed.; Fox, J.G., Davisson, M.T., Quimby, F.W., Barthold, S.W., Newcomer, C.E., Smith, A.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume III, pp. 637–672. [Google Scholar]
- Augusto, V.; Padovani, C.R.; Rocha Campos, G.E. Skeletal muscle fiber types in c57bl6j mice. Braz. J. Morphol. Sci. 2004, 21, 89–94. [Google Scholar]
- Kammoun, M.; Cassar-Malek, I.; Meunier, B.; Picard, B. A simplified immunohistochemical classification of skeletal muscle fibres in mouse. Eur. J. Histochem. EJH 2014, 58, 2254. [Google Scholar] [CrossRef]
- Hamalainen, N.; Pette, D. The histochemical profiles of fast fiber types iib, iid, and iia in skeletal muscles of mouse, rat, and rabbit. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1993, 41, 733–743. [Google Scholar] [CrossRef]
- Hirofuji, C.; Nakatani, T.; Ishihara, A.; Tanaka, M.; Itoh, K.; Itoh, M.; Katsuta, S.; Ibata, Y. Cell size and succinate dehydrogenese activity of different types of fibres in different regions of the tibialis anterior muscle in mice and rats. Acta Histochem. Cytochem. 2000, 33, 295–303. [Google Scholar] [CrossRef][Green Version]
- Agbulut, O.; Destombes, J.; Thiesson, D.; Butler-Browne, G. Age-related appearance of tubular aggregates in the skeletal muscle of almost all male inbred mice. Histochem. Cell Biol. 2000, 114, 477–481. [Google Scholar] [CrossRef]
- Gonzalez, E.; Messi, M.L.; Delbono, O. The specific force of single intact extensor digitorum longus and soleus mouse muscle fibers declines with aging. J. Membr. Biol. 2000, 178, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.V.; Faulkner, J.A. Contractile properties of skeletal muscles from young, adult and aged mice. J. Physiol. 1988, 404, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Lanza, I.R.; Befroy, D.E.; Kent-Braun, J.A. Age-related changes in atp-producing pathways in human skeletal muscle in vivo. J. Appl. Physiol. 2005, 99, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.; Toniolo, L.; Nagaraj, N.; Ciciliot, S.; Vindigni, V.; Schiaffino, S.; Reggiani, C.; Mann, M. Single muscle fiber proteomics reveals fiber-type-specific features of human muscle aging. Cell Rep. 2017, 19, 2396–2409. [Google Scholar] [CrossRef]
- Gueugneau, M.; Coudy-Gandilhon, C.; Meunier, B.; Combaret, L.; Taillandier, D.; Polge, C.; Attaix, D.; Roche, F.; Feasson, L.; Barthelemy, J.C.; et al. Lower skeletal muscle capillarization in hypertensive elderly men. Exp. Gerontol. 2016, 76, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Ballak, S.B.; Buse-Pot, T.; Harding, P.J.; Yap, M.H.; Deldicque, L.; de Haan, A.; Jaspers, R.T.; Degens, H. Blunted angiogenesis and hypertrophy are associated with increased fatigue resistance and unchanged aerobic capacity in old overloaded mouse muscle. Age 2016, 38, 39. [Google Scholar] [CrossRef] [PubMed]
- Toniolo, L.; Giacomello, E. Resveratrol, aging and fatigue. In Aging, 2nd ed.; Preedy, V., Patel, V., Eds.; Elsevier: San Diego, CA, USA, in press.
- Coggan, A.R.; Spina, R.J.; King, D.S.; Rogers, M.A.; Brown, M.; Nemeth, P.M.; Holloszy, J.O. Histochemical and enzymatic comparison of the gastrocnemius muscle of young and elderly men and women. J. Gerontol. 1992, 47, B71–B76. [Google Scholar] [CrossRef]
- Barnouin, Y.; McPhee, J.S.; Butler-Browne, G.; Bosutti, A.; De Vito, G.; Jones, D.A.; Narici, M.; Behin, A.; Hogrel, J.Y.; Degens, H. Coupling between skeletal muscle fiber size and capillarization is maintained during healthy aging. J. Cachexia Sarcopenia Muscle 2017, 8, 647–659. [Google Scholar] [CrossRef]
- Toniolo, L.; Formoso, L.; Torelli, L.; Crea, E.; Bergamo, A.; Sava, G.; Giacomello, E. Long-term resveratrol treatment improves the capillarization in the skeletal muscles of ageing c57bl/6j mice. Int. J. Food Sci. Nutr. 2020, 1–8. [Google Scholar] [CrossRef]
- Toniolo, L.; Fusco, P.; Formoso, L.; Mazzi, A.; Canato, M.; Reggiani, C.; Giacomello, E. Resveratrol treatment reduces the appearance of tubular aggregates and improves the resistance to fatigue in aging mice skeletal muscles. Exp.Gerontol. 2018, 111, 170–179. [Google Scholar] [CrossRef]
- Narici, M.V.; Maffulli, N. Sarcopenia: Characteristics, mechanisms and functional significance. Br. Med. Bull. 2010, 95, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Listrat, A.; Meunier, B.; Gueugneau, M.; Coudy-Gandilhon, C.; Combaret, L.; Taillandier, D.; Polge, C.; Attaix, D.; Lethias, C.; et al. Apoptosis in capillary endothelial cells in ageing skeletal muscle. Aging Cell 2014, 13, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Nakata, R.; Takahashi, S.; Inoue, H. Recent advances in the study on resveratrol. Biol. Pharm. Bull. 2012, 35, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bies, E.; Tung, B.T.; Navas, P.; Lopez-Lluch, G. Resveratrol primes the effects of physical activity in old mice. Br. J. Nutr. 2016, 116, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Kamei, M.; Isogai, S.; Pan, W.; Weinstein, B.M. Imaging blood vessels in the zebrafish. Methods Cell Biol. 2010, 100, 27–54. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giacomello, E.; Crea, E.; Torelli, L.; Bergamo, A.; Reggiani, C.; Sava, G.; Toniolo, L. Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice. Int. J. Mol. Sci. 2020, 21, 3923. https://doi.org/10.3390/ijms21113923
Giacomello E, Crea E, Torelli L, Bergamo A, Reggiani C, Sava G, Toniolo L. Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice. International Journal of Molecular Sciences. 2020; 21(11):3923. https://doi.org/10.3390/ijms21113923
Chicago/Turabian StyleGiacomello, Emiliana, Emanuela Crea, Lucio Torelli, Alberta Bergamo, Carlo Reggiani, Gianni Sava, and Luana Toniolo. 2020. "Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice" International Journal of Molecular Sciences 21, no. 11: 3923. https://doi.org/10.3390/ijms21113923
APA StyleGiacomello, E., Crea, E., Torelli, L., Bergamo, A., Reggiani, C., Sava, G., & Toniolo, L. (2020). Age Dependent Modification of the Metabolic Profile of the Tibialis Anterior Muscle Fibers in C57BL/6J Mice. International Journal of Molecular Sciences, 21(11), 3923. https://doi.org/10.3390/ijms21113923