The Unfolded Protein Response: Neutron-Induced Therapy Autophagy as a Promising Treatment Option for Osteosarcoma

 , ,
, ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Neutrons Induce Autophagy in a Dose-Dependent Manner in OS Cells
2.2. Relationship Between Autophagy and Cell Survival After Exposure to Gamma-Rays and LET Neutrons
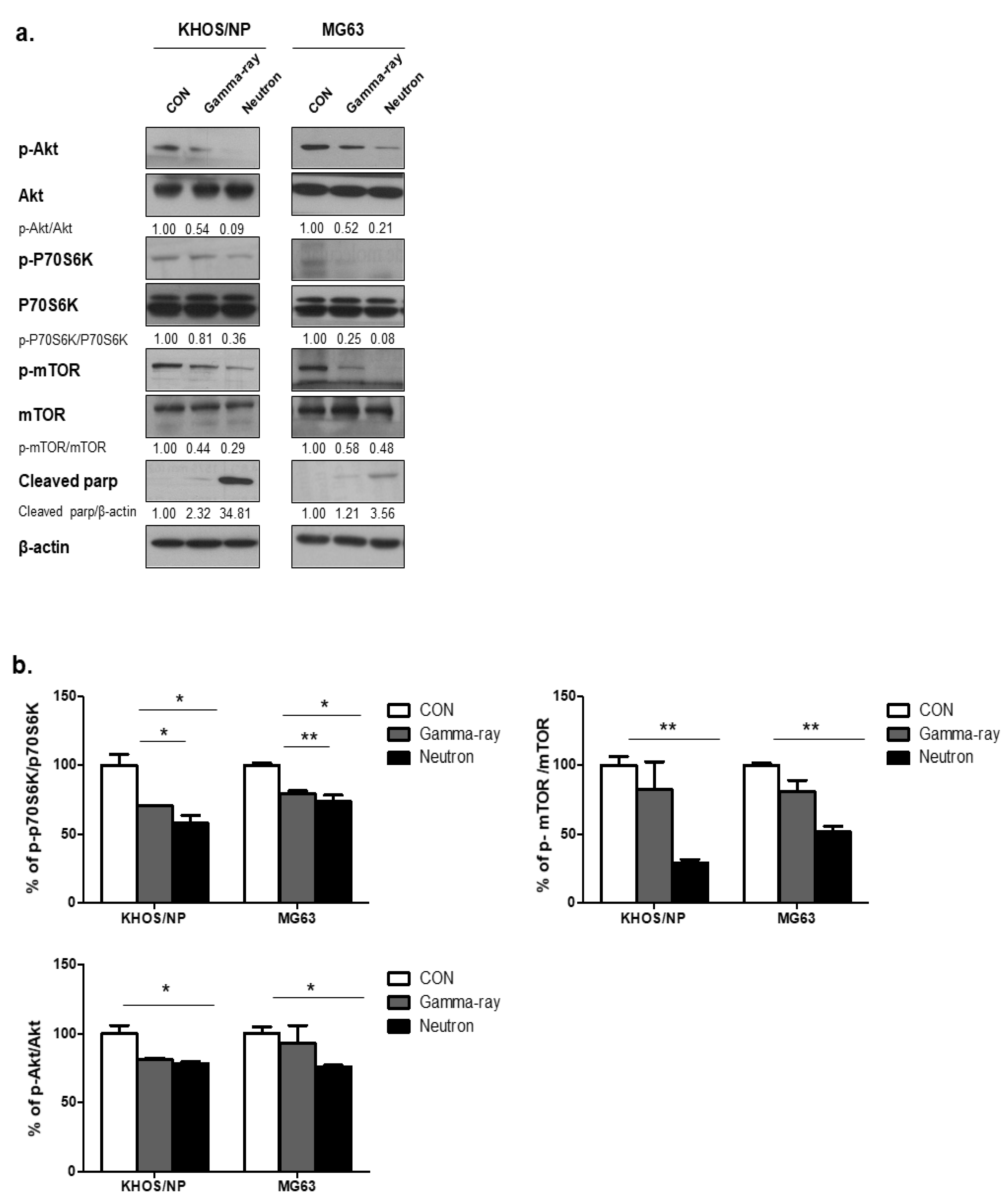
2.3. Neutron Therapy Induces Autophagy in Tumor Cells by Inhibiting the Akt-mTOR Pathway
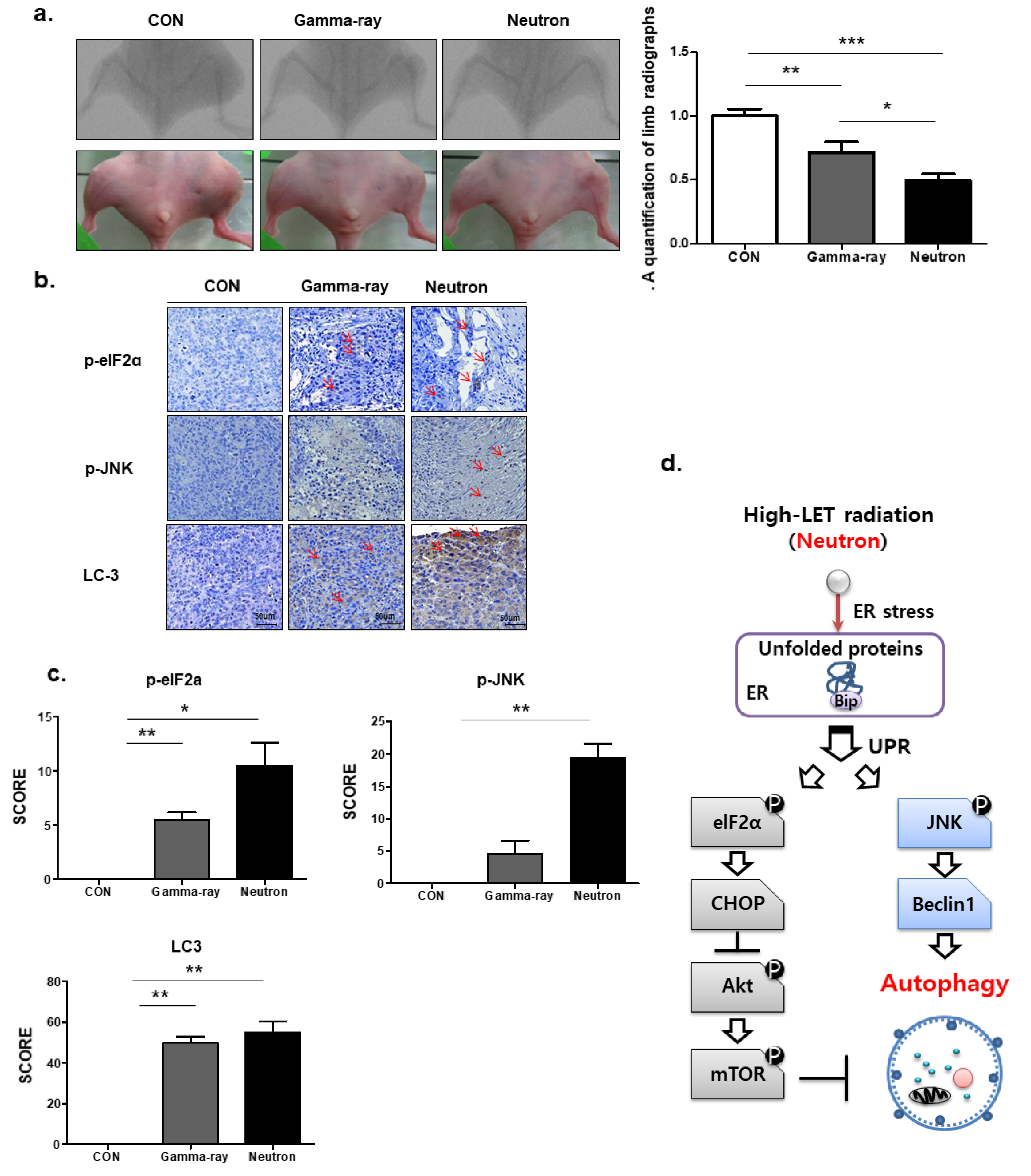
2.4. Neutrons Induce Autophagy Via Activation of Endoplasmic Reticulum (ER) Stress
2.5. Autophagic Effects of Low-LET Gamma-Ray or High-LET Neutron Exposure on Orthotopic Tumors In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Tissue Samples
4.2. Irradiation
4.3. Quantification of Acidic Vacuoles (AVOs) by Acridine Orange (AO) Staining
4.4. ELISA
4.5. Colony Formation Assay
4.6. Detection of Apoptotic Cells by Annexin V Staining
4.7. Western Blotting
4.8. Morphology
4.9. Orthotopic Model and Histological Analysis
4.10. Immunohistochemical Staining
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FNT | Fast neutron therapy |
| LET | Linear energy transfer |
| AVO | Acidic vacuoles |
| AO | Acridine orange |
References
- Chadwick, M.; DeLuca, P., Jr.; Haight, R.C. Nuclear data needs for neutron therapy and radiation protection. Radiat. Prot. Dosim. 1997, 70, 1–12. [Google Scholar] [CrossRef]
- Koh, W.J.; Griffin, T.W.; Rasey, J.S.; Laramore, G.E. Positron emission tomography. A new tool for characterization of malignant disease and selection of therapy. Acta Oncol. 1994, 33, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Fowler, J.F.; Morgan, R.L.; Wood, C.A. Pre-therapeutic experiments with the fast neutron beam from the Medical Research Council cyclotron. I. The biological and physical advantages and problems of neutron therapy. Br. J. Radiol. 1963, 36, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Prott, F.J.; Micke, O.; Haverkamp, U.; Potter, R.; Willich, N. Treatment results of fast neutron irradiation in soft tissue sarcomas. Strahlenther. Onkol. 1999, 175, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.L.; Einck, J.; Bellon, J.; Laramore, G.E. Fast neutron radiotherapy for soft tissue and cartilaginous sarcomas at high risk for local recurrence. Int. J. Radiat. Oncol. Biol. Phys. 2001, 50, 449–456. [Google Scholar] [CrossRef]
- Blake, P.; Catterall, M.; Errington, R. Treatment of malignant melanoma by fast neutrons. Br. J. Surg. 1985, 72, 517–519. [Google Scholar] [CrossRef]
- Tsunemoto, H.; Morita, S.; Mori, S. Characteristics of malignant melanoma cells in the treatment with fast neutrons. Pigment. Cell Res. 1989, 2, 372–378. [Google Scholar] [CrossRef]
- Stelzer, K.J.; Douglas, J.G.; Mankoff, D.A.; Silbergeld, D.L.; Krohn, K.A.; Laramore, G.E.; Spence, A.M. Positron emission tomography-guided conformal fast neutron therapy for glioblastoma multiforme. Neuro. Oncol. 2008, 10, 88–92. [Google Scholar] [CrossRef]
- Kageji, T.; Nagahiro, S.; Matsuzaki, K.; Mizobuchi, Y.; Toi, H.; Nakagawa, Y.; Kumada, H. Boron neutron capture therapy using mixed epithermal and thermal neutron beams in patients with malignant glioma-correlation between radiation dose and radiation injury and clinical outcome. Int. J. Radiat. Oncol. Biol. Phys. 2006, 65, 1446–1455. [Google Scholar] [CrossRef]
- Attix, F.H. Introduction to Radiological Physics and Radiation Dosimetry; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Griffin, T.W. Fast neutron radiation therapy. Crit. Rev. Oncol. Hemat. 1992, 13, 17–31. [Google Scholar] [CrossRef]
- Carrie, C.; Breteau, N.; Negrier, S.; Frappaz, D.; Bouffet, E.; Thiesse, P.; Blay, J.Y.; Clavel, M.; Philip, T.; Biron, P.; et al. The role of fast neutron therapy in unresectable pelvic osteosarcoma: Preliminary report. Med. Pediatr. Oncol. 1994, 22, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C.; Yuan, H.-X.; Guan, K.-L. Autophagy regulation by nutrient signaling. Cell Res. 2014, 24, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Ohsumi, Y. Historical landmarks of autophagy research. Cell Res. 2014, 24, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Jinno-Oue, A.; Shimizu, N.; Hamada, N.; Wada, S.; Tanaka, A.; Shinagawa, M.; Ohtsuki, T.; Mori, T.; Saha, M.N.; Hoque, A.S.; et al. Irradiation with carbon ion beams induces apoptosis, autophagy, and cellular senescence in a human glioma-derived cell line. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Hino, M.; Hamada, N.; Tajika, Y.; Funayama, T.; Morimura, Y.; Sakashita, T.; Yokota, Y.; Fukamoto, K.; Mutou, Y.; Kobayashi, Y.; et al. Heavy ion irradiation induces autophagy in irradiated C2C12 myoblasts and their bystander cells. J. Electron. Microsc. Tokyo 2010, 59, 495–501. [Google Scholar] [CrossRef]
- Jin, X.; Liu, Y.; Ye, F.; Liu, X.; Furusawa, Y.; Wu, Q.; Li, F.; Zheng, X.; Dai, Z.; Li, Q. Role of autophagy in high linear energy transfer radiation-induced cytotoxicity to tumor cells. Cancer Sci. 2014, 105, 770–778. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar]
- Furusawa, Y.; Fukutsu, K.; Aoki, M.; Itsukaichi, H.; Eguchi-Kasai, K.; Ohara, H.; Yatagai, F.; Kanai, T.; Ando, K. Inactivation of aerobic and hypoxic cells from three different cell lines by accelerated (3)He-, (12)C- and (20)Ne-ion beams. Radiat. Res. 2000, 154, 485–496. [Google Scholar] [CrossRef]
- Jin, X.D.; Gong, L.; Guo, C.L.; Hao, J.F.; Wei, W.; Dai, Z.Y.; Li, Q. Survivin expressions in human hepatoma HepG2 cells exposed to ionizing radiation of different LET. Radiat. Environ. Biophys. 2008, 47, 399–404. [Google Scholar] [CrossRef]
- Chaachouay, H.; Ohneseit, P.; Toulany, M.; Kehlbach, R.; Multhoff, G.; Rodemann, H.P. Autophagy contributes to resistance of tumor cells to ionizing radiation. Radiother. Oncol. 2011, 99, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Hu, F.; Zhang, S.S.; Zhao, Q.D.; Zong, C.; Ye, F.; Guo, S.W.; Zhang, J.W.; Li, R.; Wu, M.C.; et al. Inhibition of p53 increases chemosensitivity to 5-FU in nutrient-deprived hepatocarcinoma cells by suppressing autophagy. Cancer Lett. 2014, 346, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Si, S.; Schoen, S.; Chen, J.; Jin, X.B.; Wu, G. Suppression of autophagy enhances preferential toxicity of paclitaxel to folliculin-deficient renal cancer cells. J. Exp. Clin. Cancer Res. 2013, 32, 99. [Google Scholar] [CrossRef] [PubMed]
- Ganley, I.G.; Lam du, H.; Wang, J.; Ding, X.; Chen, S.; Jiang, X. ULK1.ATG13.FIP200 complex mediates mTOR signaling and is essential for autophagy. J. Biol. Chem. 2009, 284, 12297–12305. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Jun, C.B.; Ro, S.H.; Kim, Y.M.; Otto, N.M.; Cao, J.; Kundu, M.; Kim, D.H. ULK-Atg13-FIP200 complexes mediate mTOR signaling to the autophagy machinery. Mol. Biol. Cell 2009, 20, 1992–2003. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Takahashi, A.; Kajihara, A.; Yamakawa, N.; Imai, Y.; Ota, I.; Okamoto, N.; Mori, E.; Noda, T.; Furusawa, Y.; et al. Depression of p53-independent Akt survival signals in human oral cancer cells bearing mutated p53 gene after exposure to high-LET radiation. Biochem. Biophys. Res. Commun. 2012, 423, 654–660. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Y.; Pang, X.; Su, Y.; Ai, G.; Wang, T. ER stress induced by ionising radiation in IEC-6 cells. Int. J. Radiat. Biol. 2010, 86, 429–435. [Google Scholar] [CrossRef]
- Chiu, H.W.; Fang, W.H.; Chen, Y.L.; Wu, M.D.; Yuan, G.F.; Ho, S.Y.; Wang, Y.J. Monascuspiloin enhances the radiation sensitivity of human prostate cancer cells by stimulating endoplasmic reticulum stress and inducing autophagy. PLoS ONE 2012, 7, e40462. [Google Scholar] [CrossRef]
- Verfaillie, T.; Salazar, M.; Velasco, G.; Agostinis, P. Linking ER Stress to Autophagy: Potential Implications for Cancer Therapy. Int. J. Cell Biol. 2010, 2010, 930509. [Google Scholar] [CrossRef]
- Li, T.; Su, L.; Zhong, N.; Hao, X.; Zhong, D.; Singhal, S.; Liu, X. Salinomycin induces cell death with autophagy through activation of endoplasmic reticulum stress in human cancer cells. Autophagy 2013, 9, 1057–1068. [Google Scholar] [CrossRef]
- Huang, H.; Kang, R.; Wang, J.; Luo, G.; Yang, W.; Zhao, Z. Hepatitis C virus inhibits AKT-tuberous sclerosis complex (TSC), the mechanistic target of rapamycin (MTOR) pathway, through endoplasmic reticulum stress to induce autophagy. Autophagy 2013, 9, 175–195. [Google Scholar] [CrossRef]
- Nishitoh, H.; Saitoh, M.; Mochida, Y.; Takeda, K.; Nakano, H.; Rothe, M.; Miyazono, K.; Ichijo, H. ASK1 is essential for JNK/SAPK activation by TRAF2. Mol. Cell 1998, 2, 389–395. [Google Scholar] [CrossRef]
- Shi, J.M.; Bai, L.L.; Zhang, D.M.; Yiu, A.; Yin, Z.Q.; Han, W.L.; Liu, J.S.; Li, Y.; Fu, D.Y.; Ye, W.C. Saxifragifolin D induces the interplay between apoptosis and autophagy in breast cancer cells through ROS-dependent endoplasmic reticulum stress. Biochem. Pharmacol. 2013, 85, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Laschi, M.; Bernardini, G.; Geminiani, M.; Manetti, F.; Mori, M.; Spreafico, A.; Campanacci, D.; Capanna, R.; Schenone, S.; Botta, M.; et al. Differentially activated Src kinase in chemo-naive human primary osteosarcoma cells and effects of a Src kinase inhibitor. Biofactors 2017, 43, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Seo, S.; Ji, Y.J. Characteristics of radiation generated by BNCT irradiator of Hanaro nuclear reactor. J. Korean Soc. Ther. Radiol. Oncol. 2006, 24, s158. [Google Scholar]
- Eom, K.Y.; Wu, H.G.; Park, H.J.; Huh, S.N.; Ye, S.J.; Lee, D.H.; Park, S.W. Oncology, Evaluation of biological characteristics of neutron beam generated from MC50 cyclotron. J. Korean Soc. Ther. Radiol. Oncol. 2006, 24, 280–284. [Google Scholar]
- Gingras, A.C.; Gygi, S.P.; Raught, B.; Polakiewicz, R.D.; Abraham, R.T.; Hoekstra, M.F.; Aebersold, R.; Sonenberg, N. Regulation of 4E-BP1 phosphorylation: A novel two-step mechanism. Genes Dev. 1999, 13, 1422–1437. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, M.S.; Lee, K.H.; Koh, J.S.; Jung, W.G.; Kong, C.B. Zoledronic acid is an effective radiosensitizer in the treatment of osteosarcoma. Oncotarget 2016, 7, 70869–70880. [Google Scholar] [CrossRef]
- Nagelkerke, A.; Bussink, J.; Geurts-Moespot, A.; Sweep, F.C.; Span, P.N. Therapeutic targeting of autophagy in cancer. Part II: Pharmacological modulation of treatment-induced autophagy. Semin. Cancer Biol. 2015, 31, 99–105. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.Y.; Lee, Y.-J.; Sai, S.; Ohno, T.; Kong, C.-B.; Lim, S.H.; Kim, E.H. The Unfolded Protein Response: Neutron-Induced Therapy Autophagy as a Promising Treatment Option for Osteosarcoma. Int. J. Mol. Sci. 2020, 21, 3766. https://doi.org/10.3390/ijms21113766
Oh JY, Lee Y-J, Sai S, Ohno T, Kong C-B, Lim SH, Kim EH. The Unfolded Protein Response: Neutron-Induced Therapy Autophagy as a Promising Treatment Option for Osteosarcoma. International Journal of Molecular Sciences. 2020; 21(11):3766. https://doi.org/10.3390/ijms21113766
Chicago/Turabian StyleOh, Ju Yeon, Yeon-Joo Lee, Sei Sai, Tatsuya Ohno, Chang-Bae Kong, Sun Ha Lim, and Eun Ho Kim. 2020. "The Unfolded Protein Response: Neutron-Induced Therapy Autophagy as a Promising Treatment Option for Osteosarcoma" International Journal of Molecular Sciences 21, no. 11: 3766. https://doi.org/10.3390/ijms21113766
APA StyleOh, J. Y., Lee, Y.-J., Sai, S., Ohno, T., Kong, C.-B., Lim, S. H., & Kim, E. H. (2020). The Unfolded Protein Response: Neutron-Induced Therapy Autophagy as a Promising Treatment Option for Osteosarcoma. International Journal of Molecular Sciences, 21(11), 3766. https://doi.org/10.3390/ijms21113766