Molecular Analysis of the ABCA4 Gene Mutations in Patients with Stargardt Disease Using Human Hair Follicles

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. ABCA4 Gene is Expressed in Human Hair Follicles and Skin
2.2. Hair Follicles and Skin Cells Express the Full-Length ABCA4 Transcript and Protein
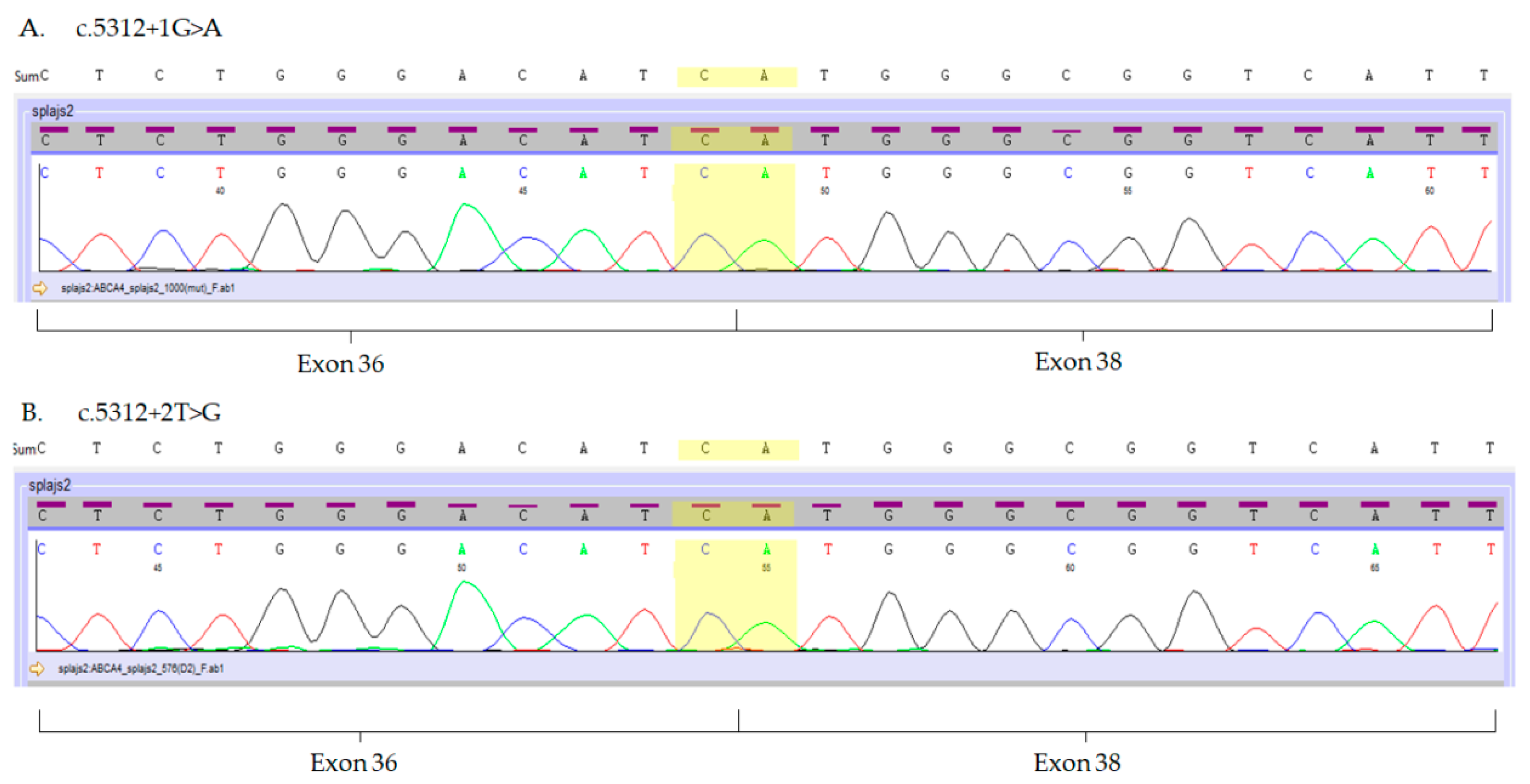
2.3. Analysis of the ABCA4 Splice-Site Variants in Hair Follicles
2.4. Analysis of the ABCA4 Gene Expression in Hair Follicles of Patients with Stargardt Disease
3. Discussion
4. Materials and Methods
4.1. Patients and Controls
4.2. Culture of Primary Keratinocytes, Fibroblasts and Melanocytes
4.3. RNA Isolation and Quantitative mRNA Measurements
4.4. Sanger Sequencing of the ABCA4 Transcripts
4.5. Protein Extract Isolation and Western Blot Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Tsybovsky, Y.; Molday, R.S.; Palczewski, K. The ATP-binding cassette transporter ABCA4: Structural and functional properties and role in retinal disease. Adv. Exp. Med. Biol. 2010, 703, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Sheffield, V.C.; Stone, E.M. Genomics and the eye. N. Engl. J. Med. 2011, 364, 1932–1942. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; White, K.; Stohr, H.; Steiner, K.; Hemmrich, N.; Grimm, T.; Jurklies, B.; Lorenz, B.; Scholl, H.P.; Apfelstedt-Sylla, E.; et al. A comprehensive survey of sequence variation in the ABCA4 (ABCR) gene in Stargardt disease and age-related macular degeneration. Am. J. Hum. Genet. 2000, 67, 800–813. [Google Scholar] [CrossRef] [PubMed]
- Valverde, D.; Riveiro-Alvarez, R.; Bernal, S.; Jaakson, K.; Baiget, M.; Navarro, R.; Ayuso, C. Microarray-based mutation analysis of the ABCA4 gene in Spanish patients with Stargardt disease: Evidence of a prevalent mutated allele. Mol. Vis. 2006, 12, 902–908. [Google Scholar]
- Maugeri, A.; Van Driel, M.A.; Van de Pol, D.J.; Klevering, B.J.; Van Haren, F.J.; Tijmes, N.; Bergen, A.A.; Rohrschneider, K.; Blankenagel, A.; Pinckers, A.J.; et al. The 2588G→C mutation in the ABCR gene is a mild frequent founder mutation in the Western European population and allows the classification of ABCR mutations in patients with Stargardt disease. Am. J. Hum. Genet. 1999, 64, 1024–1035. [Google Scholar] [CrossRef]
- Rosenberg, T.; Klie, F.; Garred, P.; Schwartz, M. N965S is a common ABCA4 variant in Stargardt-related retinopathies in the Danish population. Mol. Vis. 2007, 13, 1962–1969. [Google Scholar]
- Sciezynska, A.; Ozieblo, D.; Ambroziak, A.M.; Korwin, M.; Szulborski, K.; Krawczynski, M.; Stawinski, P.; Szaflik, J.; Szaflik, J.P.; Ploski, R.; et al. Next-generation sequencing of ABCA4: High frequency of complex alleles and novel mutations in patients with retinal dystrophies from Central Europe. Exp. Eye Res. 2016, 145, 93–99. [Google Scholar] [CrossRef]
- Nassisi, M.; Mohand-Said, S.; Dhaenens, C.M.; Boyard, F.; Demontant, V.; Andrieu, C.; Antonio, A.; Condroyer, C.; Foussard, M.; Mejecase, C.; et al. Expanding the Mutation Spectrum in ABCA4: Sixty Novel Disease Causing Variants and Their Associated Phenotype in a Large French Stargardt Cohort. Int. J. Mol. Sci. 2018, 19, 2196. [Google Scholar] [CrossRef]
- Salles, M.V.; Motta, F.L.; Martin, R.; Filippelli-Silva, R.; Dias da Silva, E.; Varela, P.; Costa, K.A.; Chiang, J.P.; Pesquero, J.B.; Sallum, J.F. Variants in the ABCA4 gene in a Brazilian population with Stargardt disease. Mol. Vis. 2018, 24, 546–559. [Google Scholar]
- Chacon-Camacho, O.F.; Granillo-Alvarez, M.; Ayala-Ramirez, R.; Zenteno, J.C. ABCA4 mutational spectrum in Mexican patients with Stargardt disease: Identification of 12 novel mutations and evidence of a founder effect for the common p.A1773V mutation. Exp. Eye Res. 2013, 109, 77–82. [Google Scholar] [CrossRef]
- September, A.V.; Vorster, A.A.; Ramesar, R.S.; Greenberg, L.J. Mutation spectrum and founder chromosomes for the ABCA4 gene in South African patients with Stargardt disease. Invest. Ophthalmol Vis. Sci. 2004, 45, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, S.S.; Bax, N.M.; Zernant, J.; Allikmets, R.; Fritsche, L.G.; Den Dunnen, J.T.; Ajmal, M.; Hoyng, C.B.; Cremers, F.P. In Silico Functional Meta-Analysis of 5,962 ABCA4 Variants in 3,928 Retinal Dystrophy Cases. Hum. Mutat. 2017, 38, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Cremers, F.P.M. ABCA4-Associated Stargardt Disease. Klin. Mon. Fur Augenheilkd. 2020, 237, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Klevering, B.J.; Rohrschneider, K.; Blankenagel, A.; Brunner, H.G.; Deutman, A.F.; Hoyng, C.B.; Cremers, F.P. Mutations in the ABCA4 (ABCR) gene are the major cause of autosomal recessive cone-rod dystrophy. Am. J. Hum. Genet. 2000, 67, 960–966. [Google Scholar] [CrossRef]
- D’Angelo, R.; Donato, L.; Venza, I.; Scimone, C.; Aragona, P.; Sidoti, A. Possible protective role of the ABCA4 gene c.1268A>G missense variant in Stargardt disease and syndromic retinitis pigmentosa in a Sicilian family: Preliminary data. Int. J. Mol. Med. 2017, 39, 1011–1020. [Google Scholar] [CrossRef][Green Version]
- Bauwens, M.; Garanto, A.; Sangermano, R.; Naessens, S.; Weisschuh, N.; De Zaeytijd, J.; Khan, M.; Sadler, F.; Balikova, I.; Van Cauwenbergh, C.; et al. ABCA4-associated disease as a model for missing heritability in autosomal recessive disorders: Novel noncoding splice, cis-regulatory, structural, and recurrent hypomorphic variants. Genet. Med. Off. J. Am. Coll. Med. Genet. 2019, 21, 1761–1771. [Google Scholar] [CrossRef]
- Runhart, E.H.; Sangermano, R.; Cornelis, S.S.; Verheij, J.; Plomp, A.S.; Boon, C.J.F.; Lugtenberg, D.; Roosing, S.; Bax, N.M.; Blokland, E.A.W.; et al. The Common ABCA4 Variant p.Asn1868Ile Shows Nonpenetrance and Variable Expression of Stargardt Disease When Present in trans With Severe Variants. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3220–3231. [Google Scholar] [CrossRef]
- Zernant, J.; Lee, W.; Collison, F.T.; Fishman, G.A.; Sergeev, Y.V.; Schuerch, K.; Sparrow, J.R.; Tsang, S.H.; Allikmets, R. Frequent hypomorphic alleles account for a significant fraction of ABCA4 disease and distinguish it from age-related macular degeneration. J. Med. Genet. 2017, 54, 404–412. [Google Scholar] [CrossRef]
- Zernant, J.; Lee, W.; Nagasaki, T.; Collison, F.T.; Fishman, G.A.; Bertelsen, M.; Rosenberg, T.; Gouras, P.; Tsang, S.H.; Allikmets, R. Extremely hypomorphic and severe deep intronic variants in the ABCA4 locus result in varying Stargardt disease phenotypes. Mol. Case Stud. 2018, 4. [Google Scholar] [CrossRef]
- Cremers, F.P.M.; Lee, W.; Collin, R.W.J.; Allikmets, R. Clinical spectrum, genetic complexity and therapeutic approaches for retinal disease caused by ABCA4 mutations. Prog. Retin. Eye Res. 2020. [Google Scholar] [CrossRef]
- Sciezynska, A.; Ozieblo, D.; Oldak, M. Experimenal studies On cell and gene therapies for retinal dystrophies with a particular focus On ABCA4 retinopathies. Klin Ocz. 2016, 118, 66–71. [Google Scholar]
- Sciezynska, A.; Ozieblo, D.; Oldak, M. Experimental studies on medical treatments of retinal dystrophies with a particular focus on ABCA4 retinopathies. Klin Ocz. 2016, 118, 59–65. [Google Scholar]
- Aukrust, I.; Jansson, R.W.; Bredrup, C.; Rusaas, H.E.; Berland, S.; Jorgensen, A.; Haug, M.G.; Rodahl, E.; Houge, G.; Knappskog, P.M. The intronic ABCA4 c.5461-10T>C variant, frequently seen in patients with Stargardt disease, causes splice defects and reduced ABCA4 protein level. Acta Ophthalmol. 2017, 95, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Sangermano, R.; Khan, M.; Cornelis, S.S.; Richelle, V.; Albert, S.; Garanto, A.; Elmelik, D.; Qamar, R.; Lugtenberg, D.; Van den Born, L.I.; et al. ABCA4 midigenes reveal the full splice spectrum of all reported noncanonical splice site variants in Stargardt disease. Genome Res. 2018, 28, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Desviat, L.R.; Perez, B.; Ugarte, M. Minigenes to confirm exon skipping mutations. Methods Mol. Biol. 2012, 867, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.A.; Mullins, R.F.; Wagner, A.H.; Andorf, J.L.; Johnston, R.M.; Bakall, B.B.; Deluca, A.P.; Fishman, G.A.; Lam, B.L.; Weleber, R.G.; et al. Non-exomic and synonymous variants in ABCA4 are an important cause of Stargardt disease. Hum. Mol. Genet. 2013, 22, 5136–5145. [Google Scholar] [CrossRef] [PubMed]
- Albert, S.; Garanto, A.; Sangermano, R.; Khan, M.; Bax, N.M.; Hoyng, C.B.; Zernant, J.; Lee, W.; Allikmets, R.; Collin, R.W.J.; et al. Identification and Rescue of Splice Defects Caused by Two Neighboring Deep-Intronic ABCA4 Mutations Underlying Stargardt Disease. Am. J. Hum. Genet. 2018, 102, 517–527. [Google Scholar] [CrossRef]
- Fadaie, Z.; Khan, M.; Del Pozo-Valero, M.; Cornelis, S.S.; Ayuso, C.; Cremers, F.P.M.; Roosing, S.; The Abca Study, G. Identification of splice defects due to noncanonical splice site or deep-intronic variants in ABCA4. Hum. Mutat. 2019, 40, 2365–2376. [Google Scholar] [CrossRef]
- Sangermano, R.; Garanto, A.; Khan, M.; Runhart, E.H.; Bauwens, M.; Bax, N.M.; Van den Born, L.I.; Khan, M.I.; Cornelis, S.S.; Verheij, J.; et al. Deep-intronic ABCA4 variants explain missing heritability in Stargardt disease and allow correction of splice defects by antisense oligonucleotides. Genet. Med. Off. J. Am. Coll. Med. Genet. 2019, 21, 1751–1760. [Google Scholar] [CrossRef]
- Khan, M.; Cornelis, S.S.; Pozo-Valero, M.D.; Whelan, L.; Runhart, E.H.; Mishra, K.; Bults, F.; AlSwaiti, Y.; AlTalbishi, A.; De Baere, E.; et al. Resolving the dark matter of ABCA4 for 1054 Stargardt disease probands through integrated genomics and transcriptomics. Genet. Med. Off. J. Am. Coll. Med. Genet. 2020. [Google Scholar] [CrossRef]
- Claassen, J.N.; Zhang, D.; Chen, S.C.; Moon, S.Y.; Lamey, T.; Thompson, J.A.; McLaren, T.; De Roach, J.N.; McLenachan, S.; Chen, F.K. Generation of the induced pluripotent stem cell line from a patient with autosomal recessive ABCA4-mediated Stargardt Macular Dystrophy. Stem Cell Res. 2019, 34, 101352. [Google Scholar] [CrossRef]
- Garanto, A.; Duijkers, L.; Tomkiewicz, T.Z.; Collin, R.W.J. Antisense Oligonucleotide Screening to Optimize the Rescue of the Splicing Defect Caused by the Recurrent Deep-Intronic ABCA4 Variant c.4539+2001G>A in Stargardt Disease. Genes 2019, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Sangermano, R.; Bax, N.M.; Bauwens, M.; Van den Born, L.I.; De Baere, E.; Garanto, A.; Collin, R.W.; Goercharn-Ramlal, A.S.; Den Engelsman-van Dijk, A.H.; Rohrschneider, K.; et al. Photoreceptor Progenitor mRNA Analysis Reveals Exon Skipping Resulting from the ABCA4 c.5461-10T-->C Mutation in Stargardt Disease. Ophthalmology 2016, 123, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Wiszniewski, W.; Zaremba, C.M.; Yatsenko, A.N.; Jamrich, M.; Wensel, T.G.; Lewis, R.A.; Lupski, J.R. ABCA4 mutations causing mislocalization are found frequently in patients with severe retinal dystrophies. Hum. Mol. Genet. 2005, 14, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000198691-ABCA4/tissue (accessed on 2 April 2020).
- Wiley, L.; Kaalberg, E.; Penticoff, J.; Mullins, R.; Stone, E.; Tucker, B. Expression of the retina-specific flippase, ABCA4, in epidermal keratinocytes [version 1; peer review: 2 approved with reservations]. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Haslam, I.S.; El-Chami, C.; Faruqi, H.; Shahmalak, A.; O’Neill, C.A.; Paus, R. Differential expression and functionality of ATP-binding cassette transporters in the human hair follicle. Br. J. Derm. 2015, 172, 1562–1572. [Google Scholar] [CrossRef]
- Riera, M.; Patel, A.; Bures-Jelstrup, A.; Corcostegui, B.; Chang, S.; Pomares, E.; Corneo, B.; Sparrow, J.R. Generation of two iPS cell lines (FRIMOi003-A and FRIMOi004-A) derived from Stargardt patients carrying ABCA4 compound heterozygous mutations. Stem Cell Res. 2019, 36, 101389. [Google Scholar] [CrossRef]
- Takenaka, S.; Itoh, T.; Fujiwara, R. Expression pattern of human ATP-binding cassette transporters in skin. Pharm. Res. Perspect. 2013, 1, e00005. [Google Scholar] [CrossRef]
- Makelainen, S.; Godia, M.; Hellsand, M.; Viluma, A.; Hahn, D.; Makdoumi, K.; Zeiss, C.J.; Mellersh, C.; Ricketts, S.L.; Narfstrom, K.; et al. An ABCA4 loss-of-function mutation causes a canine form of Stargardt disease. PLoS Genet. 2019, 15, e1007873. [Google Scholar] [CrossRef]
- Pasello, M.; Giudice, A.M.; Scotlandi, K. The ABC subfamily A transporters: Multifaceted players with incipient potentialities in cancer. Semin. Cancer Biol. 2020, 60, 57–71. [Google Scholar] [CrossRef]
- Schulz, H.L.; Grassmann, F.; Kellner, U.; Spital, G.; Ruther, K.; Jagle, H.; Hufendiek, K.; Rating, P.; Huchzermeyer, C.; Baier, M.J.; et al. Mutation Spectrum of the ABCA4 Gene in 335 Stargardt Disease Patients From a Multicenter German Cohort-Impact of Selected Deep Intronic Variants and Common SNPs. Investig. Ophthalmol. Vis. Sci. 2017, 58, 394–403. [Google Scholar] [CrossRef]
- Guo, Z.; Draheim, K.; Lyle, S. Isolation and culture of adult epithelial stem cells from human skin. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]
- Sciezynska, A.; Nogowska, A.; Sikorska, M.; Konys, J.; Karpinska, A.; Komorowski, M.; Oldak, M.; Malejczyk, J. Isolation and culture of human primary keratinocytes-a methods review. Exp. Derm. 2019, 28, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exon Number | 3–4 | 10–11 | 20–21 | 29–30 | 35–36 | 40–41 | 49–50 |
|---|---|---|---|---|---|---|---|
| Hair follicles (n = 8) | 33.72 | 28.18 | 28.53 | 27.65 | 27.22 | 27.05 | 27.28 |
| Total skin samples (n = 4) | 37.77 | 32.51 | 33.17 | Not tested | 33.48 | Not tested | 31.92 |
| Keratinocytes (n = 4) | 35.07 | 34.42 | 34.15 | Not tested | 33.02 | Not tested | 34.12 |
| Fibroblasts (n = 4) | 33.67 | 32.75 | 32.54 | Not tested | 32.15 | Not tested | 31.48 |
| Melanocytes (n = 4) | 37.87 | 37.07 | 37.01 | Not tested | 36.29 | Not tested | 38.46 |
| TaqMan® Probes | Exon Boundary |
|---|---|
| Hs00979588_m1 | 3–4 |
| Hs00979567_m1 | 10–11 |
| Hs00979574_m1 | 20–21 |
| Hs00979581_m1 | 29–30 |
| Hs00979586_m1 | 35–36 |
| Hs00979589_m1 | 40–41 |
| Hs00979594_m1 | 49–50 |
| ABCA4 Exons | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Amplicon Length | Ta |
|---|---|---|---|---|
| 2–4 | GGAACTCGTGTGGCCTTTATCTTTATT | GTCCATGAATTGGGACAAGATGTGTAG | 331 | 58.2 |
| 4–8 | AGCTACACATCTTGTCCCAATTCATGG | TGTCTTCATACCAGTTGAAGGAGAGCA | 645 | 58.2 |
| 8–12 | CTCGGGTGCTCTCCTTCAACTGGTAT | TGGTAGAGAGCTGGTCCAGGGATACAT | 698 | 61.3 |
| 12–15 | GTGCCCTCTCTCTACTGGAGGAAAACA | AGACTGGCCTTGGAGAAGAAGGTGCT | 670 | 61.3 |
| 15–20 | CACCTTCTTCTCCAAGGCCAGTCTG | CAGGATGTTGTGCTGTGGACACATG | 787 | 61 |
| 20–25 | CACAGCACAACATCCTGTTCCAC | GAAATTCCAAAACTGCTGAGACCAA | 769 | 57.1 |
| 25–30 | GGCTGACCTTGGTCTCAGCAGT | CCTGCAGGATGGTGAAGGGTTG | 706 | 58.6 |
| 30–36 | CACAGGTCAACCCTTCACCATCCTG | TTGGATTTGTTCACCCGCTCCTGGATCA | 691 | 61 |
| 36–41 | ATCTGCGTGATTTTCTCCATGTCCTT | ATCATCTTCATCAACAATGGGCTCCT | 708 | 56.4 |
| 40–46 | GAAGGGGTGGTGTACTTCCTCCTGAC | CCTTCTCTGATGATGCTCACGATGAC | 707 | 62.7 |
| 44–49 | GTCCATCAAAATATGGGCTACTGTCCT | CTGGTCCAGTGTGGTCTGTGTGACT | 702 | 61 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ścieżyńska, A.; Soszyńska, M.; Komorowski, M.; Podgórska, A.; Krześniak, N.; Nogowska, A.; Smolińska, M.; Szulborski, K.; Szaflik, J.P.; Noszczyk, B.; et al. Molecular Analysis of the ABCA4 Gene Mutations in Patients with Stargardt Disease Using Human Hair Follicles. Int. J. Mol. Sci. 2020, 21, 3430. https://doi.org/10.3390/ijms21103430
Ścieżyńska A, Soszyńska M, Komorowski M, Podgórska A, Krześniak N, Nogowska A, Smolińska M, Szulborski K, Szaflik JP, Noszczyk B, et al. Molecular Analysis of the ABCA4 Gene Mutations in Patients with Stargardt Disease Using Human Hair Follicles. International Journal of Molecular Sciences. 2020; 21(10):3430. https://doi.org/10.3390/ijms21103430
Chicago/Turabian StyleŚcieżyńska, Aneta, Marta Soszyńska, Michał Komorowski, Anna Podgórska, Natalia Krześniak, Aleksandra Nogowska, Martyna Smolińska, Kamil Szulborski, Jacek P. Szaflik, Bartłomiej Noszczyk, and et al. 2020. "Molecular Analysis of the ABCA4 Gene Mutations in Patients with Stargardt Disease Using Human Hair Follicles" International Journal of Molecular Sciences 21, no. 10: 3430. https://doi.org/10.3390/ijms21103430
APA StyleŚcieżyńska, A., Soszyńska, M., Komorowski, M., Podgórska, A., Krześniak, N., Nogowska, A., Smolińska, M., Szulborski, K., Szaflik, J. P., Noszczyk, B., Ołdak, M., & Malejczyk, J. (2020). Molecular Analysis of the ABCA4 Gene Mutations in Patients with Stargardt Disease Using Human Hair Follicles. International Journal of Molecular Sciences, 21(10), 3430. https://doi.org/10.3390/ijms21103430