Abstract
Astrocytes play a major role in the pathogenesis of a range of neurodegenerative diseases, including Alzheimer’s disease (AD), undergoing dramatic morphological and molecular changes that can cause potentially both beneficial and detrimental effects. They comprise a heterogeneous population, requiring a panel of specific phenotype markers to identify astrocyte subtypes, changes in function and their relation to pathology. This study aimed to characterise expression of the astrocyte marker N-myc downstream regulated gene 2 (NDRG2) in the ageing brain, investigate the relationship between NDRG2 and a panel of astrocyte markers, and relate NDRG2 expression to pathology. NDRG2 specifically immunolabelled the cell body and radiating processes of astrocytes in the temporal cortex of the Cognitive Function and Ageing Study (CFAS) neuropathology cohort. Expression of NDRG2 did not correlate with other astrocyte markers, including glial fibrillary acidic protein (GFAP), excitatory amino acid transporter 2 (EAAT2) and glutamine synthetase (GS). NDRG2 showed a relationship to AT8+ neurofibrillary tangles (p = 0.001) and CD68+ microglia (p = 0.047), but not β-amyloid plaques or astrocyte nuclear γH2AX immunoreactivity, a marker of DNA damage response. These findings provide new insight into the astrocyte response to pathology in the ageing brain, and suggest NDRG2 may be a potential target to modulate this response.
1. Introduction
While much of the focus in dementia research has been on the neuron, other cell types are also important. Astrocytes, the most abundant cell type in the central nervous system (CNS), play an essential role in homeostasis, including neuronal support, synapse formation and function, maintenance and metabolism [1,2]. They are classified into two major groups on the basis of their morphology and anatomical location; the fibrous astrocytes of the white matter and the protoplasmic astrocytes of the grey matter [2]. Astrocytes comprise a heterogeneous population [3,4], and different markers may reflect different functional and structural aspects of the astrocyte, and their relationship to pathology.
Disruption of the normal astrocyte–neuronal interaction can lead to synaptic dysfunction and contribute to cognitive impairment, as recently reviewed [5]. Production of pro-inflammatory cytokines and loss of neuronal support, such as neurotransmitter recycling, may contribute to neurodegeneration. As part of the tripartite synapse, astrocytes regulate levels of neurotransmitters, including the excitatory transmitter glutamate, and play a major role in preventing excitotoxicity caused by the accumulation of excess glutamate [6,7]. Astrocytes remove excess glutamate from the extracellular space via excitatory amino acid transporter 2 (EAAT2), and contain the enzyme glutamine synthetase (GS) which converts glutamate to glutamine [8]. In addition to regulating neurotransmission, astrocytes are responsible for providing metabolic support to neurons via the lactate shuttle [9].
Astrocytes react to a variety of CNS pathologies, including age-associated neurodegenerative diseases such as Alzheimer’s disease (AD), by upregulating expression of the intermediate filament glial fibrillary acidic protein (GFAP). Astrogliosis is one of the earliest changes in response to accumulating AD pathology, β-amyloid (Aβ) plaques and neurofibrillary tangles (NFT) of hyperphosphorylated tau [10]. This reactive astrogliosis is considered to play a neuroprotective role, protecting against oxidative stress and promoting the clearance of Aβ [11].
N-myc downstream regulated gene 2 (NDRG2) mRNA and its corresponding protein are principally expressed by protoplasmic and fibrous astrocytes throughout the brain [12,13,14]. Increased expression of NDRG2 has been reported in neurodegenerative diseases, including AD [15,16], frontotemporal lobar degeneration [17] and stroke [18]. The exact functions of NDRG2 are unknown but it has been shown to have an inhibitory effect on tumour proliferation [19,20], reduce production of reactive oxygen species and apoptosis [18], facilitate the hyperphosphorylation of tau [16], protect blood–brain barrier integrity [21], and facilitate the uptake of glutamate by astrocytes [22].
The Cognitive Function and Ageing Neuropathology Study (CFANS) is a large scale, population representative neuropathology cohort (>65 y) which contains the spectrum of age-associated neurodegenerative pathologies. The study combined longitudinal assessment of cognitive function, including Mini-Mental State Examination (MMSE), with a brain collection program to enable neuropathological associations with cognitive impairment to be determined without preselection into clinical groups [23]. The current study aimed to characterise the expression of NDRG2, investigate the relationship between NDRG2 and a panel of astrocyte markers (GFAP, EAAT2, GS), and relate the expression of NDRG2 to age-associated neurodegenerative pathology in the CFAS cohort.
2. Results
2.1. Comparison of the Immunoreactive Profile of NDRG2 with a Panel of Astrocyte Markers
NDRG2 exclusively immunolabelled the astrocyte cell body and radiating processes throughout the cortex (Figure 1A). Both NDRG2 and EAAT2 (Figure 1B) extensively immunolabelled the distal delicate processes of astrocytes, in comparison to antibodies against GFAP (Figure 1C) and GS (Figure 1D), which predominantly stained astrocyte cell bodies and proximal cell processes. Quantitative assessment of the immunoreactive profile of NDRG2, GFAP, GS, and EAAT2 across the cohort is shown in Table 1 and Figure 3A. Significant differences between these measures of astrocytes were detected (Friedman’s p < 0.001). NDRG2 immunoreactivity did not correlate with either brain pH (rs = −0.067, p = 0.562) or PMD (rs = 0.03, p = 0.79).
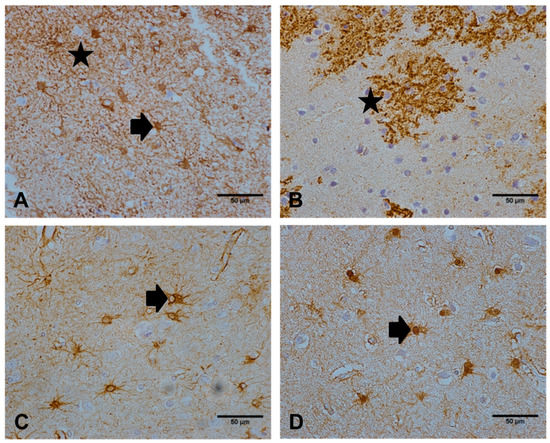
Figure 1.
Immunoreactive profiles observed with a panel of astrocyte markers. (A) N-myc downstream regulated gene 2 (NDRG2 immunolabelled the astrocyte cell body (as indicated by the arrow) and radiating processes (star). (B) excitatory amino acid transporter 2 (EAAT2) extensively immunolabelled the extending processes (star) of astrocytes. Both (C) glial fibrillary acidic protein (GFAP) and (D) glutamine synthetase (GS) predominantly stained astrocyte cell bodies and immediate cell processes (arrow). Images are representative of the astrocyte immunolabelling profile observed in the temporal cortex of 97 cases. Scale bar represents 50 μm.

Table 1.
Comparison of the immunoreactive profile (% area immunoreactivity) of the astrocyte markers NDRG2, GFAP, GS, and EAAT2 in the TCx of an ageing cohort.
2.2. NDRG2 Does Not Correlate with the Expression of Other Astrocyte Markers
Expression of NDRG2 did not correlate with levels of GFAP (rs = −0.155, p = 0.134), EAAT2 (rs = −0.178, p = 0.091) or GS (rs = 0.085, p = 0.429). In contrast, GFAP negatively correlated with EAAT2 expression (rs = −0.249, p = 0.015) and EAAT2 expression correlated with GS expression (rs = 0.209, p = 0.046) in the ageing brain. GFAP expression did not correlate with GS (rs = 0.147, p = 0.163).
2.3. Not All Astrocytes are NDRG2+
To assess if NDRG2 immunolabelled a greater proportion of astrocytes than the current standard astrocyte marker GFAP, dual labelling was performed. While co-localisation of NDRG2 with GFAP was frequently observed, both NDRG2+/GFAP− and NDRG2−/GFAP+ astrocytes were detected (Figure 2).
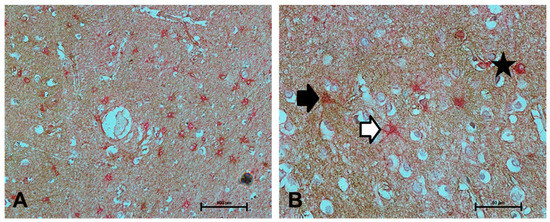
Figure 2.
Colocalisation of NDRG2 with GFAP. (A–B) Colocalisation of NDRG2 (brown) with GFAP (red) was frequently observed (as indicated by the star), however both NDRG2+/GFAP- (black arrow) and NDRG2-/GFAP+ astrocytes (white arrow) were detected in the ageing temporal cortex. Scale bar represents 100 μm in A and 50 μm in B.
2.4. NDRG2 Expression Correlates with Local Tau Pathology
NDRG2 expression did not correlate with Braak NFT stage, a measure of global AD pathology in the ageing brain (JT p = 0.786) (Figure 3B), in contrast to both GFAP (JT p = 0.032) and GS (JT p = 0.017) which showed a positive correlation.
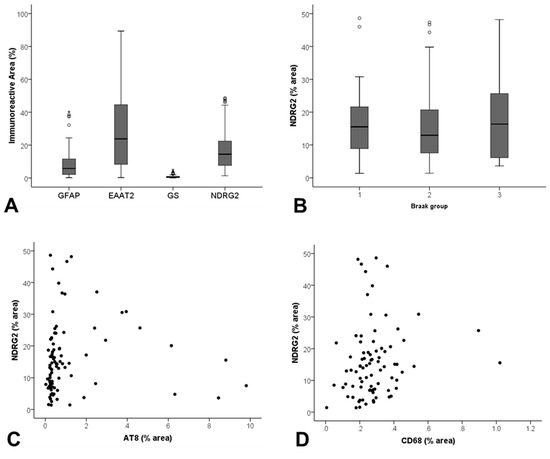
Figure 3.
Assessment of NDRG2 expression in the ageing temporal cortex. (A) Significant differences in the immunoreactive profile of a panel of astrocyte markers were detected (p < 0.001). (B) NDRG2 expression did not correlate with Braak stage, but did show a relationship to (C) local levels of tau pathology (AT8) (rs = 0.335, p = 0.001), and (D) CD68+ microglia (rs = 0.219, p = 0.047).
As astrocytes respond to pathology in their immediate vicinity, we investigated the relationship of NDRG2 to local measures of AD pathology, assessed by Aβ and AT8+% area immunoreactivity. NDRG2 expression did not correlate with Aβ (rs = 0.154, p = 0.146), but showed a relationship to AT8 (rs = 0.335, p = 0.001) (Figure 3C). Dual immunolabelling confirmed increased expression of NDRG2 by astrocytes in the vicinity of AT8+ neurones (Figure 4).
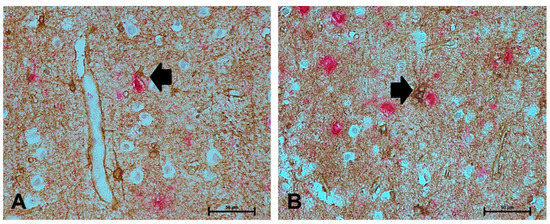
Figure 4.
NDRG2+ astrocytes associate with neurofibrillary tangles (NFT) in the ageing brain. (A–B) NDRG2+ astrocytes (brown) associated with AT8 immunoreactivity (red), as indicated by the black arrow. Scale bar represents 50 μm.
GS expression correlated with both Aβ (rs = 0.244, p = 0.02) and AT8 (rs = 0.242, p = 0.023). Neither GFAP nor EAAT2 expression correlated with local levels of Aβ (GFAP rs = 0.128, p = 0.221; EAAT2 rs0.071, p = 0.503) or AT8 (GFAP rs = 0.163, p = 0.124; EAAT2 rs = −0.18, p = 0.087).
2.5. NDRG2 Expression Correlates with CD68 but not with Oxidative DNA Damage
To determine if NDRG2 associated with microglial reactivity we investigated the relationship with CD68 and MHC II. NDRG2 correlated with CD68 (rs = 0.219, p = 0.047) (Figure 3D), but not MHC II expression (rs = 0.164, p = 0.146) in the ageing brain. No other astrocyte marker correlated with either CD68 (GFAP rs = 0.09, p = 0.409; EAAT2 rs = −0.051, p = 0.638; GS rs = 0.193, p = 0.078) or MHC II (GFAP rs = −0.177, p = 0.109; EAAT2 rs = −0.013, p = 0.904; GS rs = 0.109, p = 0.331).
To determine if NDRG2 associated with oxidative stress we investigated their relationship with γH2AX, a marker of oxidative DNA damage. NDRG2 did not correlate with astrocyte nuclear γH2AX immunoreactivity (rs = 0.179, p = 0.089). Similarly, neither GFAP (rs = 0.025, p = 0.813), nor GS (rs = 0.14, p = 0.183) correlated with γH2AX, however EAAT2 correlated with increased levels of astrocyte nuclear γH2AX immunoreactivity (rs = 0.222, p = 0.033).
3. Discussion
NDRG2 is expressed at high levels in the brain, and plays a role in growth, differentiation and development, and the stress response [24]. NDRG2 is upregulated in the hippocampus in AD in post-mortem studies [15] and animal models [16]. However, in contrast to these studies which report that NDRG2 expression is associated with neurons, the current study demonstrates NDRG2 is exclusively associated with astrocytes in the ageing brain and correlates with tau and microglial pathology.
NDRG2 was first identified in a normal human brain cDNA library in 2001 [25]. While some studies report NDRG2 expression in pyramidal neurons, senile plaques and dystrophic neurons in the hippocampus in AD [15], more recent studies investigating the expression of NDRG2 in a variety of mammalian brains, including human, demonstrate that NDRG2 is a specific marker for astrocytes and co-localises with the majority of known astrocyte markers [13,14]. In the current study, significant differences in the immunoreactive profiles of the panel of astrocyte markers were detected and the patterns of staining varied greatly across the cohort, supporting the findings of a recent study comparing NDRG2 expression with other astrocyte markers in different mouse cerebral regions [26]. Both GFAP and GS predominantly labelled astrocyte cell bodies and immediate cell processes. However, as only approximately 15% of the total astrocyte volume is GFAP+, the entire astrocyte domain is not fully visualised [27]. In contrast, antibodies to NDRG2 and EAAT2 immunolabel the extending radiating processes of astrocytes clearly demonstrating the extent of the astrocyte domain.
Levels of NDRG2 increase in AD transgenic mice (APP/PS1), and suppression of NDRG2 has been shown to improve cognitive function in this model [28]. As the NDRG2+ distal processes of astrocytes are in close proximity to synapses, this suggests a role in impairing synaptic transduction [13]. In contrast, NDRG2 deficiency has recently been shown to exacerbate long-term memory impairment in this model of AD, suggesting a neuroprotective role for NDRG2 [29]. Recent studies have shown that knockdown of NDRG2 significantly reduces tau phosphorylation in a human neuroblastoma cell line overexpressing wild type APP695 [16]. Our data demonstrate that expression of NDRG2 positively correlates with AT8+ tau pathology in the ageing brain; however, whether this is a mechanism actively contributing to neuronal pathology or a response to neurodegeneration is currently unknown.
NDRG2 is an injury response gene that positively regulates early astrocyte activation and the inflammatory response in a mouse model of cortical stab injury [30]. The reciprocal interactions between microglia and astrocytes play a key role in the progression of AD [31]. In the current study NDRG2 did not correlate with expression levels of the other astrocyte markers but did correlate with CD68, a lysosomal protein present in phagocytic microglia in the ageing brain. CD68+ microglia positively associate with dementia, neuritic plaques and tangles in the ageing population [32]. Whether NDRG2+ astrocytes are linked to microglial pathology, or vice-versa, is unknown and whether these NDRG2+ glia are adopting a neuroprotective or a neurotoxic phenotype needs to be determined. Additional studies to interrogate the association with other microglial markers, including TREM−2, are also required.
NDRG2 is mainly expressed in the cytoplasm, however it has been shown to translocate to the nucleus under stress conditions [33,34,35], and control astrocyte morphology via Rho-GTPase signalling [36]. Oxidative stress and the associated DNA damage may directly impact astrocytes, consequently modifying their normal function [37]. We previously reported a reduction in expression of γH2AX by astrocytes associated with increasing levels of Alzheimer’s type pathology [38]. Here our results demonstrate that the astrocyte DNA damage response does not correlate with NDRG2 but does correlate with expression of EAAT2. Both γH2AX and EAAT2 were highest in cases with low levels of AD pathology, and which may constitute a stress-response aimed at increasing glutamate uptake and reducing neuronal excitotoxicity at the earliest stages of AD pathology.
Ageing population-representative neuropathology cohorts, such as CFAS, are a valuable resource for investigating correlates of cellular pathology. A comprehensive panel of astrocyte markers, including NDRG2, is required to fully elucidate the complexity of the astrocyte response, providing a snapshot of different aspects of astrocyte pathobiology and how they relate to age-related neurodegenerative pathology. In the current study we demonstrate that NDRG2 is exclusively expressed by astrocytes, and that levels of NDRG2 positively correlate with neurofibrillary tangles and CD68+ microglial pathology in the temporal cortex of the CFAS cohort. Future studies are required to characterise the interaction between NDRG2+ astrocytes, microglia and tangle pathobiology, and may identify potential therapeutic targets to modify the astrocyte phenotype.
4. Materials and Methods
4.1. Human CNS Cases
Human autopsy brain tissue was obtained from one centre (Cambridge, 97 cases) of the Cognitive Function and Ageing Study (CFAS) [23,39], following multi-centre research ethics committee (REC) approval (REC Reference number 15/SW/0246, 10 August 2015). Formalin-fixed lateral temporal cortex samples (superior/middle temporal gyrus, Brodmann areas 22/21) of all cases in the subcohort were used to maintain the population-representative basis of the study. Neuropathological lesions were previously assessed as part of the core CFAS neuropathology study using a modified protocol from the Consortium to Establish a Registry of Alzheimer’s Disease (CERAD) [40] (wwws.cfas.ac.uk) and Braak neurofibrillary tangle (NFT) staging [41]. The mean age of death was 85.6 years (SEM 7.4). The median post-mortem delay was 17 h (interquartile range [IQR] 10–32 h) and brain pH 6.49 (IQR 6.25–6.75).
4.2. Immunohistochemistry
Immunohistochemistry was performed using a standard avidin-biotin complex (ABC) method. Sections were deparaffinised, rehydrated to water and endogenous peroxidase activity quenched by placing the sections in 0.3% H2O2/methanol for 20 min at room temperature (RT). Following pressure cooker antigen retrieval in Access Super pH9.5 (Menarini Diagnostics, Wokingham, UK) and incubation with 1.5% normal serum for 30 min at RT, the sections were incubated with primary antibody (Table 2). To visualise antibody binding, the horse-radish peroxidase avidin biotin complex was used (Vectastain Elite goat IgG kit, Vector Laboratories, Peterborough, UK) with 3,3′-diaminobenzidine (DAB) as the chromagen (Vector Laboratories, UK; brown). Negative controls, either omission of the primary antibody or goat isotype control, were included in every run.
Table 2.
Antibody source and specificity.
Dual labelling experiments to visualise colocalisation of NDRG2 with neurofibrillary tangles or GFAP were performed using a combined colour product method. Briefly, NDRG2 expression was first visualised as described above. Sections were then incubated with the avidin-biotin blocking kit (Vector Laboratories, UK), according to the manufacturer’s instructions. The tissue was incubated overnight at 4 °C with either anti-AT8 (1:400) or anti-GFAP (1:1000), followed by biotinylated secondary antibody, and visualised with the alkaline-phosphatase-conjugated avidin-biotin complex and alkaline phosphatase substrate 1 (Vector Laboratories, UK, red). Every immuno-run included a single-labelled section, which showed the same pattern and intensity of immunoreactivity as seen in the double-labelling experiments.
4.3. Quantitative Analysis of NDRG2
Assessment of NDRG2-specific immunoreactivity was performed by capturing bright-field microscopic images in 3 adjacent 350 μm-wide cortical ribbons, consisting of contiguous fields to cover the total cortical thickness through the apex of the gyrus, using a ×20 objective (Nikon Eclipse Ni-U microscope, Nikon Instruments Europe BV, Amsterdam, Netherlands) and analysed using the Analysis ^D software (Olympus Biosystems, Watford, UK). The image was thresholded and the immunoreactive area (%) of the field determined per total area examined.
4.4. Previously Assessed Astrocyte Markers and Age-Associated Pathology
Previous studies of the temporal cortex in the CFAS cohort using an identical approach have determined the immunoreactive profile (% area immunoreactivity) of astrocytes (GFAP, EAAT2, GS), microglia (CD68, MHC II) and AD pathology (Aβ, AT8) in 3 adjacent cortical ribbons [10,38,42]. To assess the DNA damage response, the total number of γH2AX positive astrocyte nuclei was quantitated using Analysis˄D software [38].
4.5. Statistical Analysis
As the data from all of the markers were positively skewed, non-parametric methods were used for statistical analysis. Differences between astrocyte markers were compared using Friedman’s test. For assessment in relation to Braak NFT stage, the cases were grouped into entorhinal stages (Braak stages 0-II; 30 cases), limbic stages (Braak stages III-IV; 50 cases) and isocortical stages of tangle pathology (Braak stages V-VI; 17 cases). Differences according to Braak group and to local plaque and tangle scores were assessed by Kruskal–Wallis test, and significance of trend assessed using the Jonckheere–Terpstra test (JT). Correlation analysis was performed using Spearman’s rank test. Statistical analyses were performed using the statistical package SPSS (version 23, IBM, Armonk, NY, USA).
Author Contributions
Conceptualization, J.E.S. and S.B.W.; methodology, M.M.F., J.E.S., N.G., R.W.; formal analysis, M.M.F., S.B.W., J.E.S.; writing—original draft preparation, M.M.F., J.E.S.; writing—review and editing, M.M.F., R.W., N.G., C.J.G., P.R.H., F.E.M., C.B., S.B.W., J.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
MMF is supported by a scholarship from King Abdulaziz University (KAU), funded by both KAU and the Saudi Arabian Ministry of Education. CFAS study is supported by the Department of Health and the Medical Research Council (grants MRC/G9901400 and MRC U.1052.00.0013); the UKNIHR Biomedical Research Centre for Ageing and Age-related Disease Award to the Newcastle upon Tyne Hospitals Foundation Trust; the Cambridge Brain Bank is supported by the NIHR Cambridge Biomedical Research Centre; The Cambridgeshire and Peterborough NIHR CLAHRC; Nottingham University Hospitals NHS Trust; University of Sheffield and the Sheffield Teaching Hospitals NHS Foundation Trust; The Thomas Willis Oxford Brain Collection, supported by the Oxford Biomedical Research Centre; The Walton Centre NHS Foundation Trust, Liverpool. The imaging equipment used in this study was funded by Sheffield charity Neurocare.
Acknowledgments
We would like to acknowledge the essential contribution of the liaison officers, the general practitioners, their staff, and nursing and residential home staff. We are grateful to our respondents and their families for their generous gift to medical research, which has made this study possible.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| Aβ | β-amyloid |
| AD | Alzheimer’s disease |
| CFANS | Cognitive Function and Ageing Study |
| CFAS | Cognitive Function and Ageing Neuropathology Study |
| CNS | Central nervous system |
| EAAT2 | Excitatory amino acid transporter 2 |
| GFAP | Glial fibrillary acidic protein |
| GS | Glutamine synthetase |
| NDRG2 | N-myc downstream regulated gene 2 |
| NFT | Neurofibrillary tangle |
References
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef]
- Garwood, C.J.; Ratcliffe, L.E.; Simpson, J.E.; Heath, P.R.; Ince, P.G.; Wharton, S.B. Review: Astrocytes in Alzheimer’s disease and other age-associated dementias: A supporting player with a central role. Neuropathol. Appl. Neurobiol. 2017, 43, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Wang, X.; Goldman, S.; Nedergaard, M. Astrocytic complexity distinguishes the human brain. Trends Neurosci. 2006, 29, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely hominid features of adult human astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Parpura, V.; Rodriguez-Arellano, J.J.; Zorec, R. Astroglia in Alzheimer’s Disease. Adv. Exp. Med. Biol. 2019, 1175, 273–324. [Google Scholar] [CrossRef]
- Masliah, E.; Alford, M.; DeTeresa, R.; Mallory, M.; Hansen, L. Deficient glutamate transport is associated with neurodegeneration in Alzheimer’s disease. Ann. Neurol. 1996, 40, 759–766. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef]
- Broer, S.; Brookes, N. Transfer of glutamine between astrocytes and neurons. J. Neurochem. 2001, 77, 705–719. [Google Scholar] [CrossRef]
- Newington, J.T.; Harris, R.A.; Cumming, R.C. Reevaluating Metabolism in Alzheimer’s Disease from the Perspective of the Astrocyte-Neuron Lactate Shuttle Model. J. Neurodegener. Dis. 2013, 2013, 234572. [Google Scholar] [CrossRef]
- Simpson, J.E.; Ince, P.G.; Lace, G.; Forster, G.; Shaw, P.J.; Matthews, F.; Savva, G.; Brayne, C.; Wharton, S.B. Astrocyte phenotype in relation to Alzheimer-type pathology in the ageing brain. Neurobiol. Aging 2010, 31, 578–590. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Olabarria, M.; Noristani, H.N.; Yeh, C.Y.; Rodriguez, J.J. Astrocytes in Alzheimer’s disease. Neurotherapeutics 2010, 7, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Zhai, Y.; Wei, H.; Zhang, C.; Xing, G.; Yu, Y.; He, F. Characterization and expression of three novel differentiation-related genes belong to the human NDRG gene family. Mol. Cell Biochem. 2002, 229, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Flugge, G.; Araya-Callis, C.; Garea-Rodriguez, E.; Stadelmann-Nessler, C.; Fuchs, E. NDRG2 as a marker protein for brain astrocytes. Cell Tissue Res. 2014, 357, 31–41. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Okuda, T.; Kokame, K.; Miyata, T. Differential expression patterns of NDRG family proteins in the central nervous system. J. Histochem. Cytochem. 2008, 56, 175–182. [Google Scholar] [CrossRef]
- Mitchelmore, C.; Buchmann-Moller, S.; Rask, L.; West, M.J.; Troncoso, J.C.; Jensen, N.A. NDRG2: A novel Alzheimer’s disease associated protein. Neurobiol. Dis 2004, 16, 48–58. [Google Scholar] [CrossRef]
- Rong, X.F.; Sun, Y.N.; Liu, D.M.; Yin, H.J.; Peng, Y.; Xu, S.F.; Wang, L.; Wang, X.L. The pathological roles of NDRG2 in Alzheimer’s disease, a study using animal models and APPwt-overexpressed cells. CNS Neurosci. 2017, 23, 667–679. [Google Scholar] [CrossRef]
- Herskowitz, J.H.; Seyfried, N.T.; Duong, D.M.; Xia, Q.; Rees, H.D.; Gearing, M.; Peng, J.; Lah, J.J.; Levey, A.I. Phosphoproteomic analysis reveals site-specific changes in GFAP and NDRG2 phosphorylation in frontotemporal lobar degeneration. J. Proteome. Res. 2010, 9, 6368–6379. [Google Scholar] [CrossRef]
- Ma, Y.L.; Zhang, L.X.; Liu, G.L.; Fan, Y.; Peng, Y.; Hou, W.G. N-Myc Downstream-Regulated Gene 2 (Ndrg2) Is Involved in Ischemia-Hypoxia-Induced Astrocyte Apoptosis: A Novel Target for Stroke Therapy. Mol. Neurobiol. 2017, 54, 3286–3299. [Google Scholar] [CrossRef]
- Deng, Y.; Yao, L.; Chau, L.; Ng, S.S.; Peng, Y.; Liu, X.; Au, W.S.; Wang, J.; Li, F.; Ji, S.; et al. N-Myc downstream-regulated gene 2 (NDRG2) inhibits glioblastoma cell proliferation. Int. J. Cancer 2003, 106, 342–347. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, Y.; Kang, H.B.; Jaroszewski, L.; Deacon, A.M.; Lee, H.; Choi, W.C.; Kim, K.J.; Kim, C.H.; Kang, B.S.; et al. Crystal structure of the human N-Myc downstream-regulated gene 2 protein provides insight into its role as a tumor suppressor. J. Biol. Chem. 2011, 286, 12450–12460. [Google Scholar] [CrossRef]
- Takarada-Iemata, M.; Yoshikawa, A.; Ta, H.M.; Okitani, N.; Nishiuchi, T.; Aida, Y.; Kamide, T.; Hattori, T.; Ishii, H.; Tamatani, T.; et al. N-myc downstream-regulated gene 2 protects blood–brain barrier integrity following cerebral ischemia. Glia 2018, 66, 1432–1446. [Google Scholar] [CrossRef] [PubMed]
- Yin, A.; Guo, H.; Tao, L.; Cai, G.; Wang, Y.; Yao, L.; Xiong, L.; Zhang, J.; Li, Y. NDRG2 Protects the Brain from Excitotoxicity by Facilitating Interstitial Glutamate Uptake. Transl. Stroke Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wharton, S.B.; Brayne, C.; Savva, G.M.; Matthews, F.E.; Forster, G.; Simpson, J.; Lace, G.; Ince, P.G. Epidemiological neuropathology: The MRC Cognitive Function and Aging Study experience. J. Alzheimers Dis. 2011, 25, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Yin, A.; Yao, L.; Li, Y. N-myc downstream-regulated gene 2 in the nervous system: From expression pattern to function. Acta Biochim. Biophys. Sin. 2015, 47, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.C.; Yao, L.B.; Liu, X.P.; Nie, X.Y.; Wang, X.G.; Su, C.Z. Exploring a new gene containing ACP like domain in human brain and expression in E. coli. Prog. Biochem. Biophys. 2001, 28, 72–76. [Google Scholar]
- Zhang, Z.; Ma, Z.; Zou, W.; Guo, H.; Liu, M.; Ma, Y.; Zhang, L. The Appropriate Marker for Astrocytes: Comparing the Distribution and Expression of Three Astrocytic Markers in Different Mouse Cerebral Regions. Biomed. Res. Int. 2019, 2019, 9605265. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Wang, F.; Zhong, H.; Li, X.; Peng, Y.; Kinden, R.; Liang, W.; Li, X.; Shi, M.; Liu, L.; Wang, Q.; et al. Electroacupuncture attenuates reference memory impairment associated with astrocytic NDRG2 suppression in APP/PS1 transgenic mice. Mol. Neurobiol. 2014, 50, 305–313. [Google Scholar] [CrossRef]
- Tao, L.; Zhu, Y.; Wang, R.; Han, J.; Ma, Y.; Guo, H.; Tang, W.; Zhuo, L.; Fan, Z.; Yin, A.; et al. N-myc downstream-regulated gene 2 deficiency aggravates memory impairment in Alzheimer’s disease. Behav. Brain Res. 2019, 112384. [Google Scholar] [CrossRef]
- Takarada-Iemata, M.; Kezuka, D.; Takeichi, T.; Ikawa, M.; Hattori, T.; Kitao, Y.; Hori, O. Deletion of N-myc downstream-regulated gene 2 attenuates reactive astrogliosis and inflammatory response in a mouse model of cortical stab injury. J. Neurochem. 2014, 130, 374–387. [Google Scholar] [CrossRef]
- Bouvier, D.S.; Murai, K.K. Synergistic actions of microglia and astrocytes in the progression of Alzheimer’s disease. J. Alzheimers Dis. 2015, 45, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Minett, T.; Classey, J.; Matthews, F.E.; Fahrenhold, M.; Taga, M.; Brayne, C.; Ince, P.G.; Nicoll, J.A.; Boche, D. Microglial immunophenotype in dementia with Alzheimer’s pathology. J. Neuroinflammation 2016, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhao, Z.Y.; Wang, Y.Z.; Ji, S.P.; Liu, X.P.; Liu, X.W.; Che, H.L.; Lin, W.; Li, X.; Zhang, J.; et al. Immunohistochemical detection of Ndrg2 in the mouse nervous system. Neuroreport 2008, 19, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Takeichi, T.; Takarada-Iemata, M.; Hashida, K.; Sudo, H.; Okuda, T.; Kokame, K.; Hatano, T.; Takanashi, M.; Funabe, S.; Hattori, N.; et al. The effect of Ndrg2 expression on astroglial activation. Neurochem. Int. 2011, 59, 21–27. [Google Scholar] [CrossRef]
- Li, Y.; Xu, N.; Cai, L.; Gao, Z.; Shen, L.; Zhang, Q.; Hou, W.; Zhong, H.; Wang, Q.; Xiong, L. NDRG2 is a novel p53-associated regulator of apoptosis in C6-originated astrocytes exposed to oxygen-glucose deprivation. PLoS ONE 2013, 8, e57130. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, Z.; Zou, W.; Zhang, L.; Li, Y.; Zhang, J.; Liu, M.; Hou, W.; Ma, Y. N-myc downstream-regulated gene 2 controls astrocyte morphology via Rho-GTPase signaling. J. Cell Physiol. 2019, 234, 20847–20858. [Google Scholar] [CrossRef]
- Iida, T.; Furuta, A.; Nishioka, K.; Nakabeppu, Y.; Iwaki, T. Expression of 8-oxoguanine DNA glycosylase is reduced and associated with neurofibrillary tangles in Alzheimer’s disease brain. Acta Neuropathol. 2002, 103, 20–25. [Google Scholar] [CrossRef]
- Simpson, J.E.; Ince, P.G.; Haynes, L.J.; Theaker, R.; Gelsthorpe, C.; Baxter, L.; Forster, G.; Lace, G.L.; Shaw, P.J.; Matthews, F.E.; et al. Population variation in oxidative stress and astrocyte DNA damage in relation to Alzheimer-type pathology in the ageing brain. Neuropathol. Appl Neurobiol. 2010, 36, 25–40. [Google Scholar] [CrossRef]
- Cognitive function and dementia in six areas of England and Wales: The distribution of MMSE and prevalence of GMS organicity level in the MRC CFA Study. The Medical Research Council Cognitive Function and Ageing Study (MRC CFAS). Psychol. Med. 1998, 28, 319–335.
- Mirra, S.S. The CERAD neuropathology protocol and consensus recommendations for the postmortem diagnosis of Alzheimer’s disease: A commentary. Neurobiol Aging 1997, 18, S91–S94. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Fluteau, A.; Ince, P.G.; Minett, T.; Matthews, F.E.; Brayne, C.; Garwood, C.J.; Ratcliffe, L.E.; Morgan, S.; Heath, P.R.; Shaw, P.J.; et al. The nuclear retention of transcription factor FOXO3a correlates with a DNA damage response and increased glutamine synthetase expression by astrocytes suggesting a neuroprotective role in the ageing brain. Neurosci. Lett. 2015, 609, 11–17. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).