Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
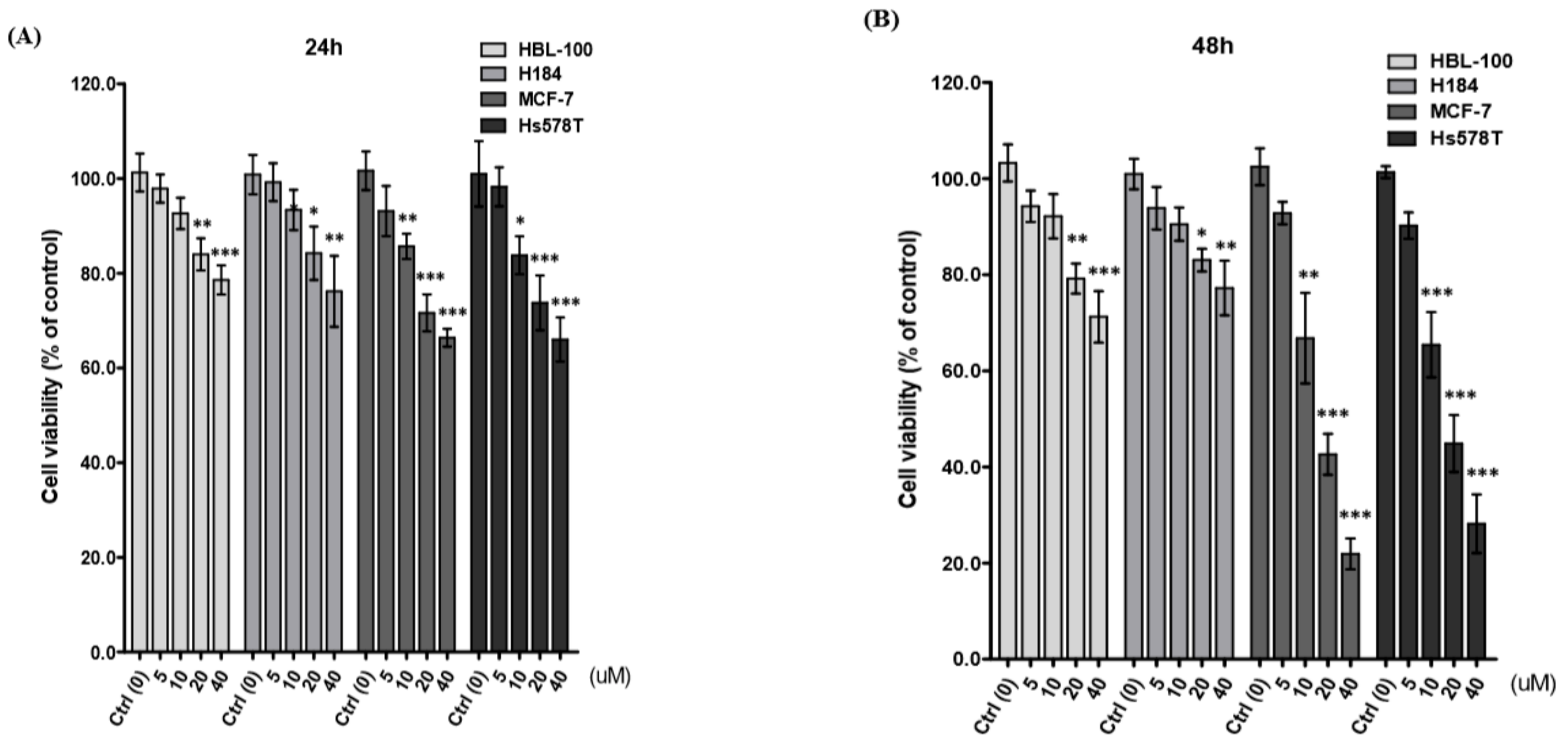
2.1. Diosgenin Inhibits Breast Cancer Cell Proliferation
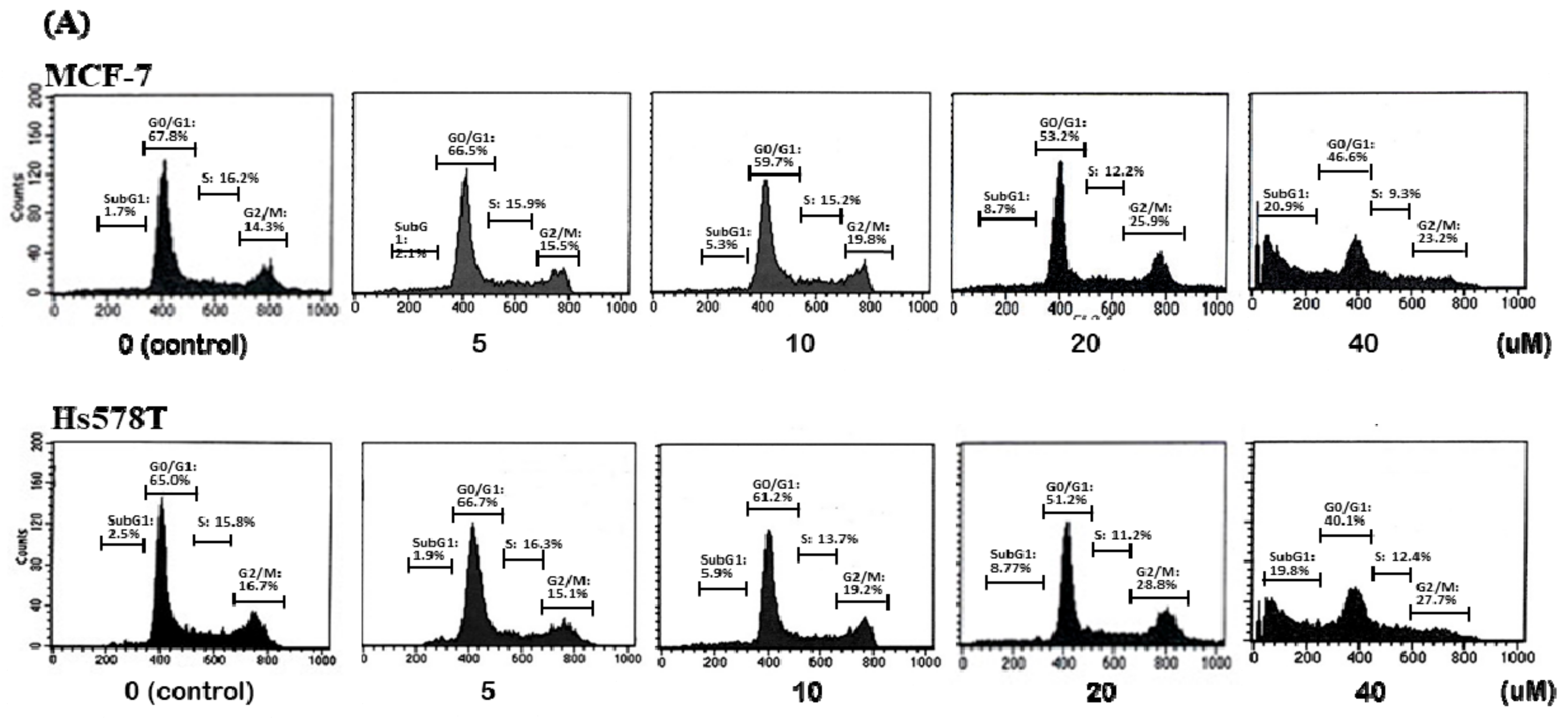
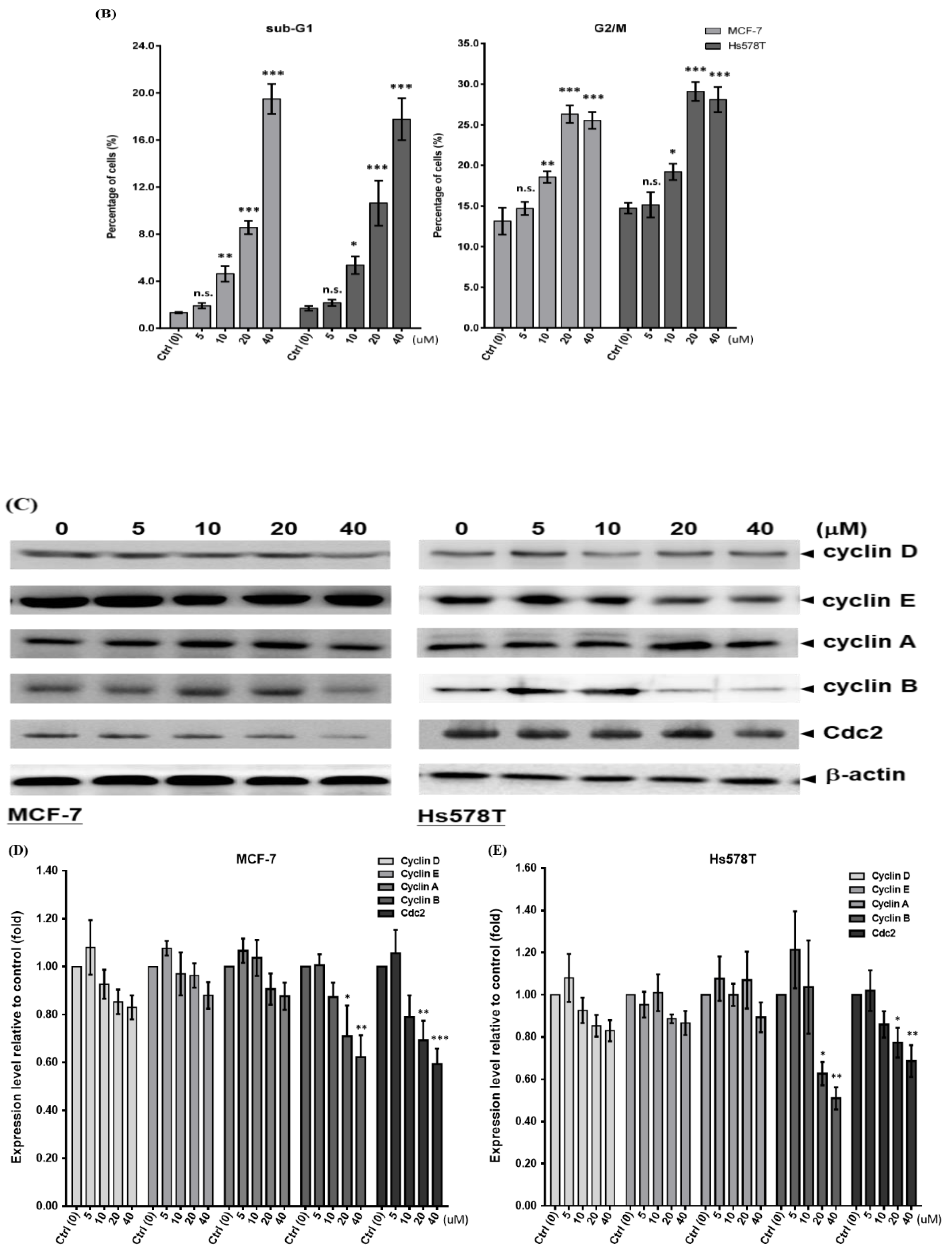
2.2. Diosgenin Induces G2/M Arrest in Breast Cancer Cell Lines
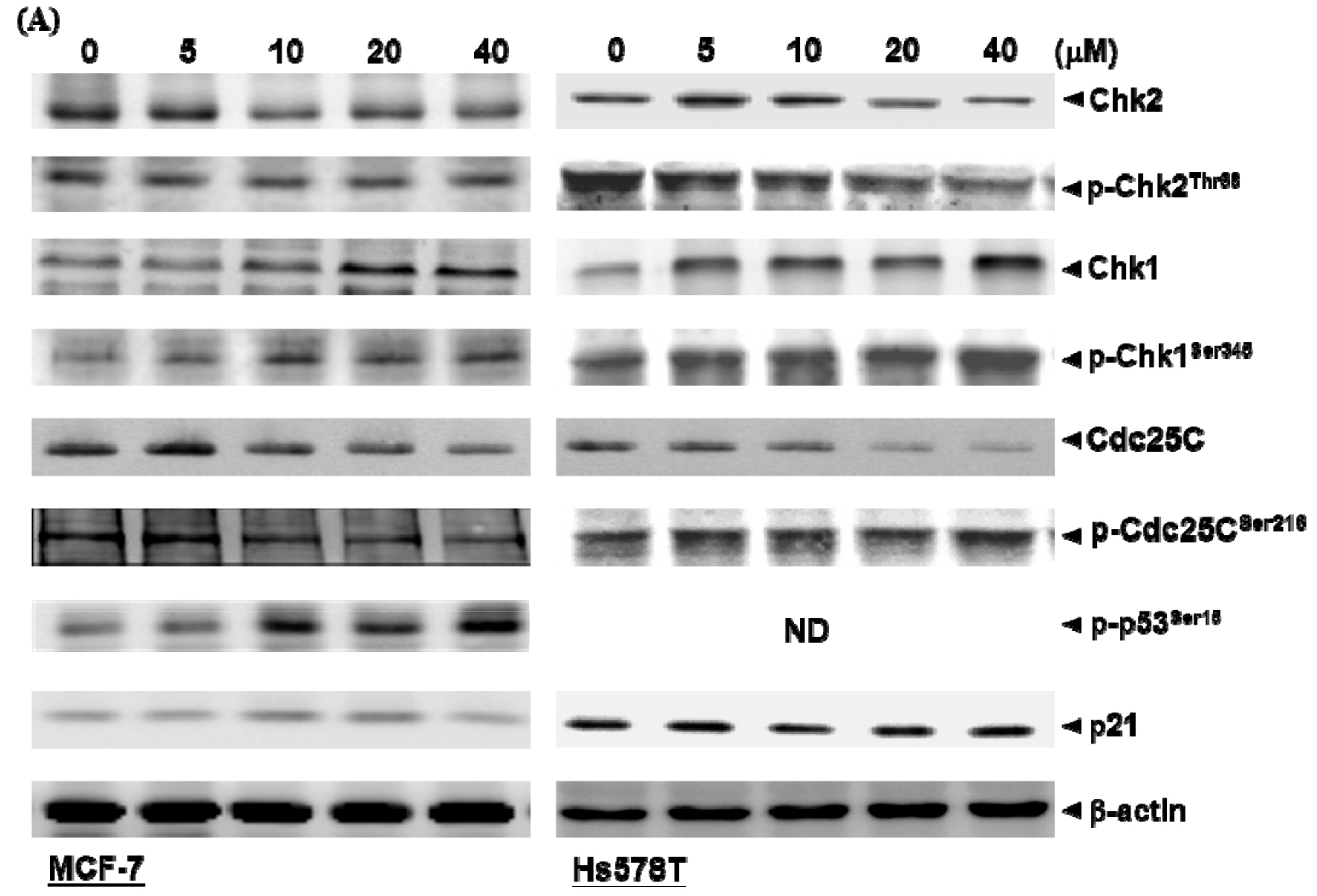
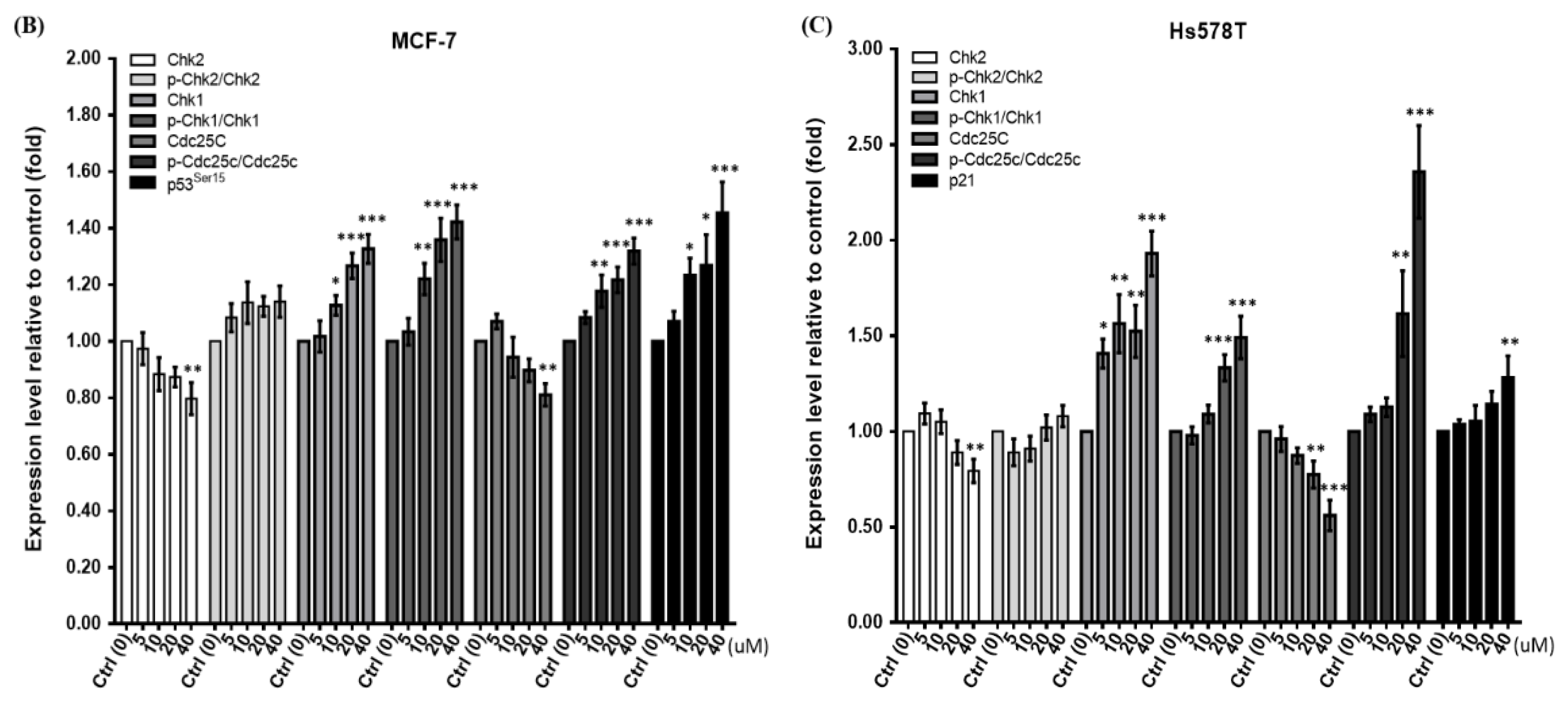
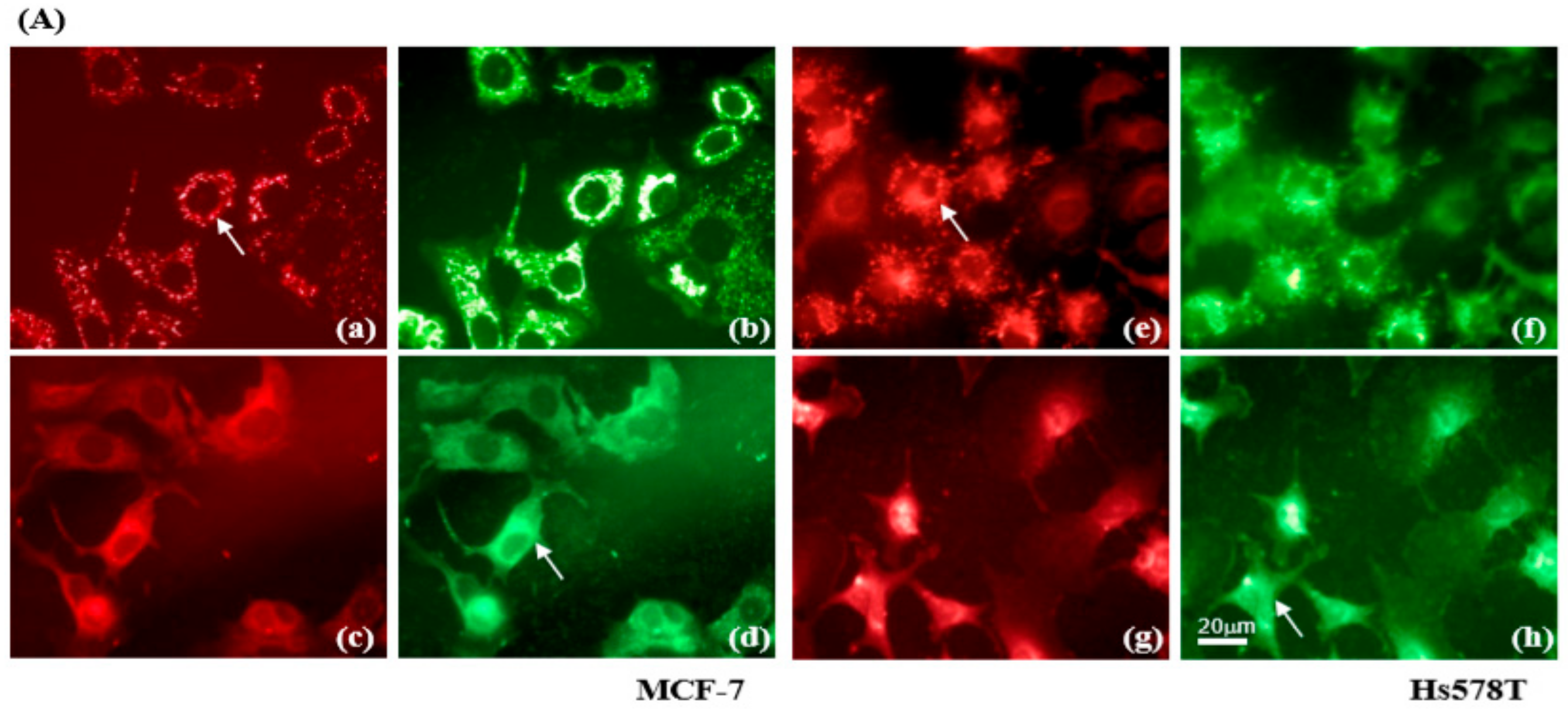
2.3. Diosgenin Primarily Activates Chk1-Mediated Growth Arrest of Breast Cancer Cells
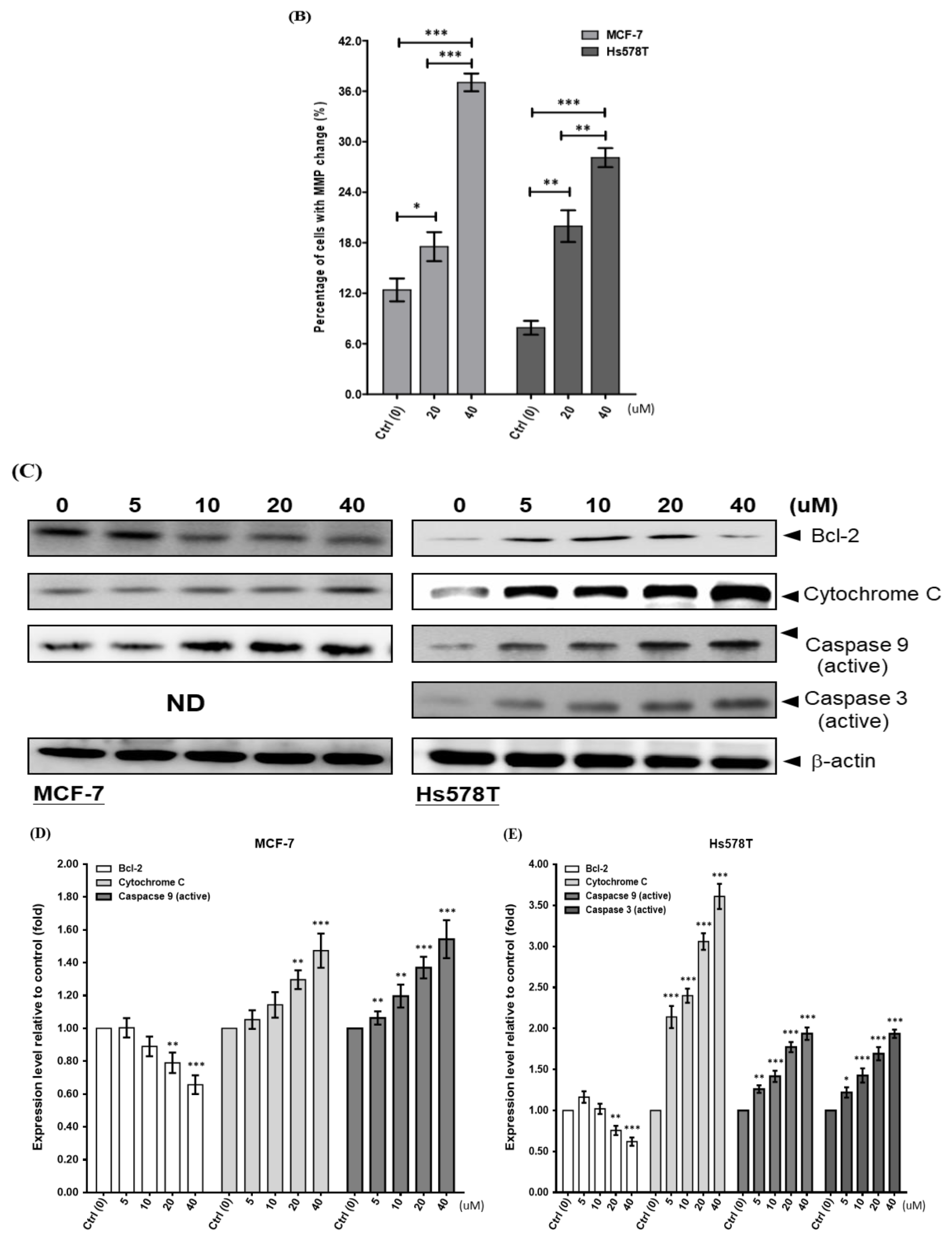
2.4. Diosgenin Induces Mitochondrial Dysfunction in Breast Cancer Lines
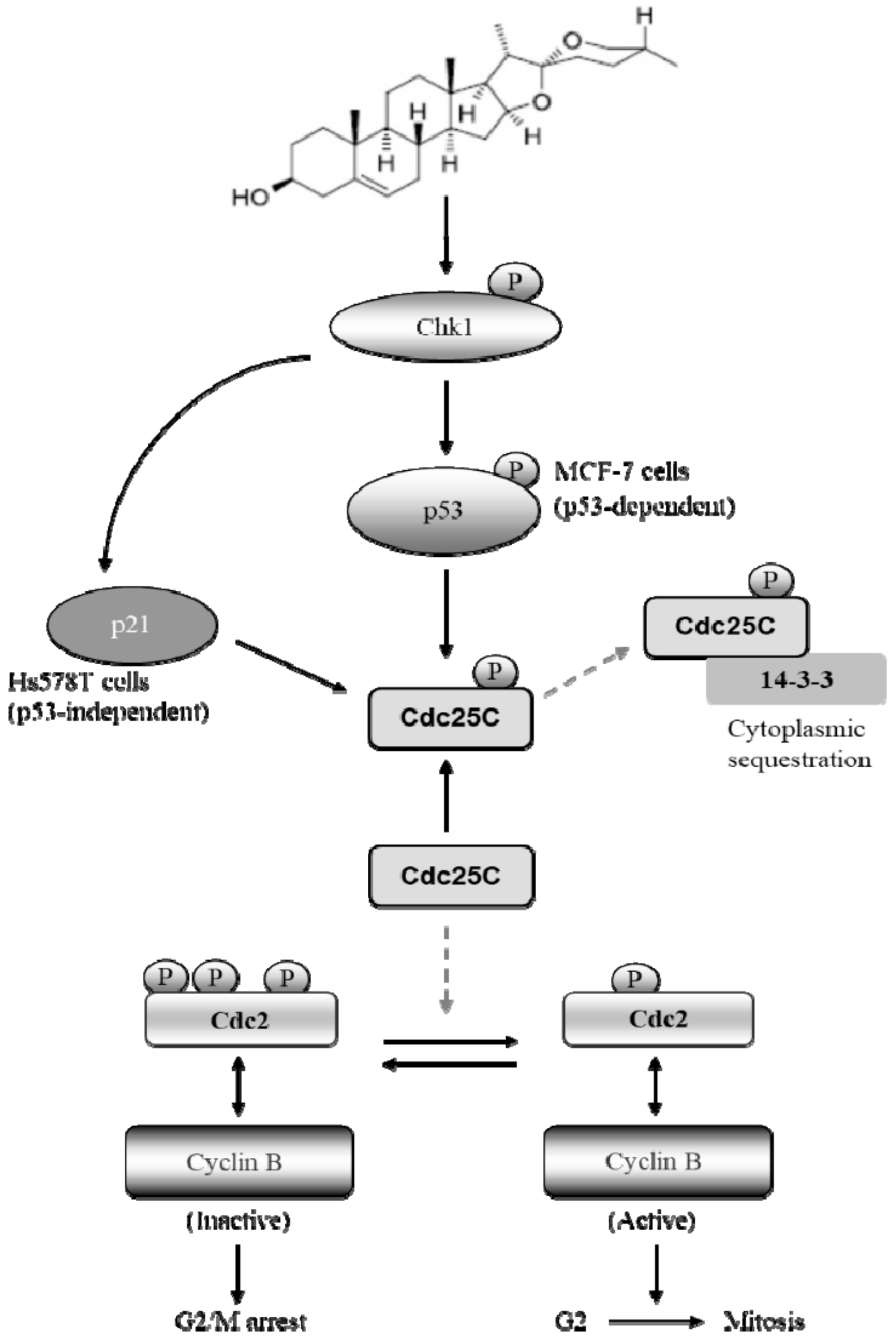
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Cycle Analysis
4.5. Western Blot Analyses
4.6. Mitochondrial Membrane Potential Assay
4.7. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Chk1 | checkpoint kinase 1 |
| Chk2 | checkpoint kinase 2 |
| CDK | cyclin-dependent kinase |
| CDKI | cyclin-dependent kinase inhibitor |
| Cdc25C | cell division cycle 25C |
| Cdc2 | cell division cycle 2 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| ∆Ψm | mitochondrial membrane potential |
References
- Ministry of Health and Welfare of Taiwan. Statistical Results on Causes of Death in Taiwan. 2017. Available online: http://www.mohw.gov.tw/mp-2.html (accessed on 12 October 2017).
- Gokduman, K. Strategies Targeting DNA Topoisomerase I in Cancer Chemotherapy: Camptothecins, Nanocarriers for Camptothecins, Organic Non-Camptothecin Compounds and Metal Complexes. Curr. Drug Targets 2016, 17, 1928–1939. [Google Scholar] [CrossRef]
- Bae, S.K.; Gwak, J.; Song, I.S.; Park, H.S.; Oh, S. Induction of apoptosis in colon cancer cells by a novel topoisomerase I inhibitor TopIn. Biochem. Biophys. Res. Commun. 2011, 409, 75–81. [Google Scholar] [CrossRef]
- Sanchez-Moreno, P.; Boulaiz, H.; Ortega-Vinuesa, J.L.; Peula-Garcia, J.M.; Aranega, A. Novel drug delivery system based on docetaxel-loaded nanocapsules as a therapeutic strategy against breast cancer cells. Int. J. Mol. Sci. 2012, 13, 4906–4919. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.R.; Huuse, E.M.; Bathen, T.F.; Goa, P.E.; Bofin, A.M.; Pedersen, T.B.; Lundgren, S.; Gribbestad, I.S. Assessment of early docetaxel response in an experimental model of human breast cancer using DCE-MRI, ex vivo HR MAS, and in vivo 1H MRS. NMR Biomed. 2010, 23, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.M.; Rosen, R.T.; Vassil, A.; Ho, C.T.; Zhang, H.; Ghai, G.; Lambert, G.; DiPaola, R.S. Modulation of bcl-2 and cytotoxicity by licochalcone-A, a novel estrogenic flavonoid. Anticancer Res. 2000, 20, 2653–2658. [Google Scholar] [PubMed]
- Yu, L.; Ma, J.; Han, J.; Wang, B.; Chen, X.; Gao, C.; Li, D.; Zheng, Q. Licochalcone B Arrests Cell Cycle Progression and Induces Apoptosis in Human Breast Cancer MCF-7 Cells. Recent Pat. Anti-Cancer Drug Discov. 2016, 11, 444–452. [Google Scholar] [CrossRef]
- He, H.; Zheng, L.; Sun, Y.P.; Zhang, G.W.; Yue, Z.G. Steroidal saponins from Paris polyphylla suppress adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMP-2 and MMP-9. Asian Pac. J. Cancer Prev. 2014, 15, 10911–10916. [Google Scholar] [CrossRef]
- Tao, X.; Yin, L.; Xu, L.; Peng, J. Dioscin: A diverse acting natural compound with therapeutic potential in metabolic diseases, cancer, inflammation and infections. Pharmacol. Res. 2018, 137, 259–269. [Google Scholar] [CrossRef]
- Morgia, G.; Voce, S.; Palmieri, F.; Gentile, M.; Iapicca, G.; Giannantoni, A.; Blefari, F.; Carini, M.; Vespasiani, G.; Santelli, G.; et al. Association between selenium and lycopene supplementation and incidence of prostate cancer: Results from the post-hoc analysis of the procomb trial. Phytomedicine 2017, 34, 1–5. [Google Scholar] [CrossRef]
- Rampogu, S.; Parameswaran, S.; Lemuel, M.R.; Lee, K.W. Exploring the Therapeutic Ability of Fenugreek against Type 2 Diabetes and Breast Cancer Employing Molecular Docking and Molecular Dynamics Simulations. Evid. Based Complement. Altern. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Selim, S.; Al Jaouni, S. Anti-inflammatory, antioxidant and antiangiogenic activities of diosgenin isolated from traditional medicinal plant, Costus speciosus (Koen ex.Retz.) Sm. Nat. Prod. Res. 2016, 30, 1830–1833. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Shanmugam, M.K.; Warrier, S.; Merarchi, M.; Arfuso, F.; Kumar, A.P.; Bishayee, A. Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients 2018, 10, 645. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.; Kumar, P. Fenugreek: A review on its nutraceutical properties and utilization in various food products. J. Saudi Soc. Agric. Sci. 2016, 17, 97–106. [Google Scholar] [CrossRef]
- Nappez, C.; Liagre, B.; Beneytout, J.L. Changes in lipoxygenase activities in human erythroleukemia (HEL) cells during diosgenin-induced differentiation. Cancer Lett. 1995, 96, 133–140. [Google Scholar] [CrossRef]
- Corbiere, C.; Liagre, B.; Bianchi, A.; Bordji, K.; Dauca, M.; Netter, P.; Beneytout, J.L. Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, on human 1547 osteosarcoma cells. Int. J. Oncol. 2003, 22, 899–905. [Google Scholar] [CrossRef]
- Corbiere, C.; Liagre, B.; Terro, F.; Beneytout, J.L. Induction of antiproliferative effect by diosgenin through activation of p53, release of apoptosis-inducing factor (AIF) and modulation of caspase-3 activity in different human cancer cells. Cell Res. 2004, 14, 188–196. [Google Scholar] [CrossRef]
- Leger, D.Y.; Liagre, B.; Beneytout, J.L. Role of MAPKs and NF-kappaB in diosgenin-induced megakaryocytic differentiation and subsequent apoptosis in HEL cells. Int. J. Oncol. 2006, 28, 201–207. [Google Scholar]
- Wang, S.L.; Cai, B.; Cui, C.B.; Liu, H.W.; Wu, C.F.; Yao, X.S. Apoptosis of human chronic myeloid leukemia k562 cell induced by prosapogenin B of dioscin (P.B) in vitro. Ai Zheng 2003, 22, 795–800. [Google Scholar]
- Hou, R.; Zhou, Q.L.; Wang, B.X.; Tashiro, S.; Onodera, S.; Ikejima, T. Diosgenin induces apoptosis in HeLa cells via activation of caspase pathway. Acta Pharmacol. Sin. 2004, 25, 1077–1082. [Google Scholar]
- Leger, D.Y.; Liagre, B.; Corbiere, C.; Cook-Moreau, J.; Beneytout, J.L. Diosgenin induces cell cycle arrest and apoptosis in HEL cells with increase in intracellular calcium level, activation of cPLA2 and COX-2 overexpression. Int. J. Oncol. 2004, 25, 555–562. [Google Scholar] [CrossRef]
- Liu, M.J.; Wang, Z.; Ju, Y.; Wong, R.N.; Wu, Q.Y. Diosgenin induces cell cycle arrest and apoptosis in human leukemia K562 cells with the disruption of Ca2+ homeostasis. Cancer Chemother. Pharmacol. 2005, 55, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liu, Y.; Niu, C.; Cheng, Y. Diosgenin increased DDX3 expression in hepatocellular carcinoma. Am. J. Transl. Res. 2018, 10, 3590–3599. [Google Scholar] [PubMed]
- Eliyahu, D.; Evans, S.; Rosen, N.; Eliyahu, S.; Zwiebel, J.; Paik, S.; Lippman, M. p53Val135 temperature sensitive mutant suppresses growth of human breast cancer cells. Breast Cancer Res. Treat. 1994, 30, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Rabi, T.; Wang, L.; Banerjee, S. Novel triterpenoid 25-hydroxy-3-oxoolean-12-en-28-oic acid induces growth arrest and apoptosis in breast cancer cells. Breast Cancer Res. Treat. 2007, 101, 27–36. [Google Scholar] [CrossRef]
- Janicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef]
- Chen, J.S.; Lin, S.Y.; Tso, W.L.; Yeh, G.C.; Lee, W.S.; Tseng, H.; Chen, L.C.; Ho, Y.S. Checkpoint kinase 1-mediated phosphorylation of Cdc25C and bad proteins are involved in antitumor effects of loratadine-induced G2/M phase cell-cycle arrest and apoptosis. Mol. Carcinog. 2006, 45, 461–478. [Google Scholar] [CrossRef]
- Aumsuwan, P.; Khan, S.I.; Khan, I.A.; Ali, Z.; Avula, B.; Walker, L.A.; Shariat-Madar, Z.; Helferich, W.G.; Katzenellenbogen, B.S.; Dasmahapatra, A.K. The anticancer potential of steroidal saponin, dioscin, isolated from wild yam (Dioscorea villosa) root extract in invasive human breast cancer cell line MDA-MB-231 in vitro. Arch. Biochem. Biophys. 2016, 591, 98–110. [Google Scholar] [CrossRef]
- Mao, Z.; Shen, X.; Dong, P.; Liu, G.; Pan, S.; Sun, X.; Hu, H.; Pan, L.; Huang, J. Fucosterol exerts antiproliferative effects on human lung cancer cells by inducing apoptosis, cell cycle arrest and targeting of Raf/MEK/ERK signalling pathway. Phytomedicine 2018, 61, 152809. [Google Scholar] [CrossRef]
- Riccardi, C.; Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef]
- Nie, C.; Zhou, J.; Qin, X.; Shi, X.; Zeng, Q.; Liu, J.; Yan, S.; Zhang, L. Diosgenininduced autophagy and apoptosis in a human prostate cancer cell line. Mol. Med. Rep. 2016, 14, 4349–4359. [Google Scholar] [CrossRef]
- Dong, M.; Meng, Z.; Kuerban, K.; Qi, F.; Liu, J.; Wei, Y.; Wang, Q.; Jiang, S.; Feng, M.; Ye, L. Diosgenin promotes antitumor immunity and PD-1 antibody efficacy against melanoma by regulating intestinal microbiota. Cell Death Dis. 2018, 9, 1039. [Google Scholar] [CrossRef]
- Tsai, Y.S.; Lee, K.W.; Huang, J.L.; Liu, Y.S.; Juo, S.H.; Kuo, W.R.; Chang, G.J.; Lin, C.S.; Jong, Y.J. Arecoline, a major alkaloid of areca nut, inhibits p53, represses DNA repair, and triggers DNA damage response in human epithelial cells. Toxicology 2008, 249, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Vafeiadou, K.; Rice-Evans, C.; Cadenas, E.; Spencer, J.P. Inhibition of cellular proliferation by the genistein metabolite 5,7,3′,4′-tetrahydroxyisoflavone is mediated by DNA damage and activation of the ATR signalling pathway. Arch. Biochem. Biophys. 2007, 468, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Calcabrini, A.; Garcia-Martinez, J.M.; Gonzalez, L.; Tendero, M.J.; Ortuno, M.T.; Crateri, P.; Lopez-Rivas, A.; Arancia, G.; González-Porqué, P.; Martín-Pérez, J. Inhibition of proliferation and induction of apoptosis in human breast cancer cells by lauryl gallate. Carcinogenesis 2006, 27, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Liao, C.H.; Chien, M.H.; Tsai, T.Y.; Lin, J.K.; Weng, M.S. Induction of p21(Waf1/Cip1) by garcinol via downregulation of p38-MAPK signaling in p53-independent H1299 lung cancer. J. Agric. Food Chem. 2014, 62, 2085–2095. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Matsumoto, M.; Kitagawa, K.; Kotake, Y.; Suzuki, S.; Shirasawa, S.; Nakayama, K.I.; Nakanishi, M.; Niida, H.; Kitagawa, M. Chk1 phosphorylates the tumour suppressor Mig-6, regulating the activation of EGF signalling. EMBO J. 2012, 31, 2365–2377. [Google Scholar] [CrossRef]
- Liu, M.J.; Yue, P.Y.; Wang, Z.; Wong, R.N. Methyl protodioscin induces G2/M arrest and apoptosis in K562 cells with the hyperpolarization of mitochondria. Cancer Lett. 2005, 224, 229–241. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H.; Huang, M.; Wang, N.; Zhang, J.; Zhang, Y.; Bai, G.; Fong, W.-F.; Yang, M.; Yao, X. Methyl protodioscin induces G2/M cell cycle arrest and apoptosis in HepG2 liver cancer cells. Cancer Lett. 2006, 241, 102–109. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef]
- Hibasami, H.; Moteki, H.; Ishikawa, K.; Katsuzaki, H.; Imai, K.; Yoshioka, K.; Ishii, Y.; Komiya, T. Protodioscin isolated from fenugreek (Trigonella foenumgraecum L.) induces cell death and morphological change indicative of apoptosis in leukemic cell line H-60, but not in gastric cancer cell line KATO III. Int. J. Mol. Med. 2003, 11, 23–26. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef] [PubMed]
- Lim do, Y.; Jeong, Y.; Tyner, A.L.; Park, J.H. Induction of cell cycle arrest and apoptosis in HT-29 human colon cancer cells by the dietary compound luteolin. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G66–G75. [Google Scholar] [CrossRef] [PubMed]
- Hirose, Y.; Berger, M.S.; Pieper, R.O. Abrogation of the Chk1-mediated G(2) checkpoint pathway potentiates temozolomide-induced toxicity in a p53-independent manner in human glioblastoma cells. Cancer Res. 2001, 61, 5843–5849. [Google Scholar] [PubMed]
- Reers, M.; Smith, T.W.; Chen, L.B. J-aggregate formation of a carbocyanine as a quantitative fluorescent indicator of membrane potential. Biochemistry 1991, 30, 4480–4486. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, W.-L.; Lin, J.-Y.; Shieh, J.-C.; Yeh, H.-F.; Hsieh, Y.-H.; Cheng, Y.-C.; Lee, H.-J.; Shen, C.-Y.; Cheng, C.-W. Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells. Int. J. Mol. Sci. 2020, 21, 172. https://doi.org/10.3390/ijms21010172
Liao W-L, Lin J-Y, Shieh J-C, Yeh H-F, Hsieh Y-H, Cheng Y-C, Lee H-J, Shen C-Y, Cheng C-W. Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells. International Journal of Molecular Sciences. 2020; 21(1):172. https://doi.org/10.3390/ijms21010172
Chicago/Turabian StyleLiao, Wen-Ling, Jing-Yi Lin, Jia-Ching Shieh, Hsiao-Fong Yeh, Yi-Hsien Hsieh, Yu-Chun Cheng, Huei-Jane Lee, Chen-Yang Shen, and Chun-Wen Cheng. 2020. "Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells" International Journal of Molecular Sciences 21, no. 1: 172. https://doi.org/10.3390/ijms21010172
APA StyleLiao, W.-L., Lin, J.-Y., Shieh, J.-C., Yeh, H.-F., Hsieh, Y.-H., Cheng, Y.-C., Lee, H.-J., Shen, C.-Y., & Cheng, C.-W. (2020). Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells. International Journal of Molecular Sciences, 21(1), 172. https://doi.org/10.3390/ijms21010172