The Role of Macrophage in the Pathogenesis of Osteoporosis



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
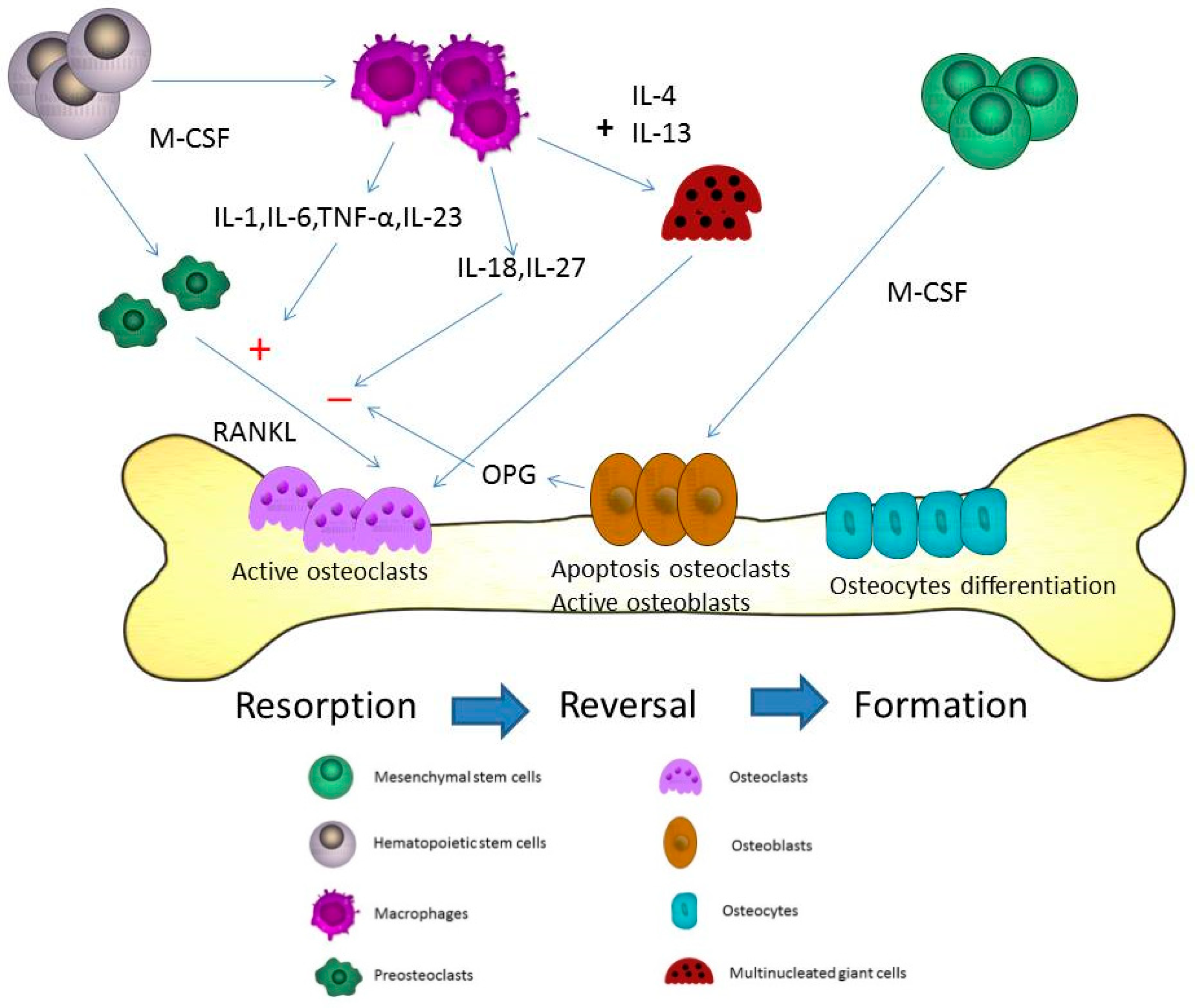
2. Osteoblast, Osteoclast and Osteocyte in Bone Formation and Homeostasis
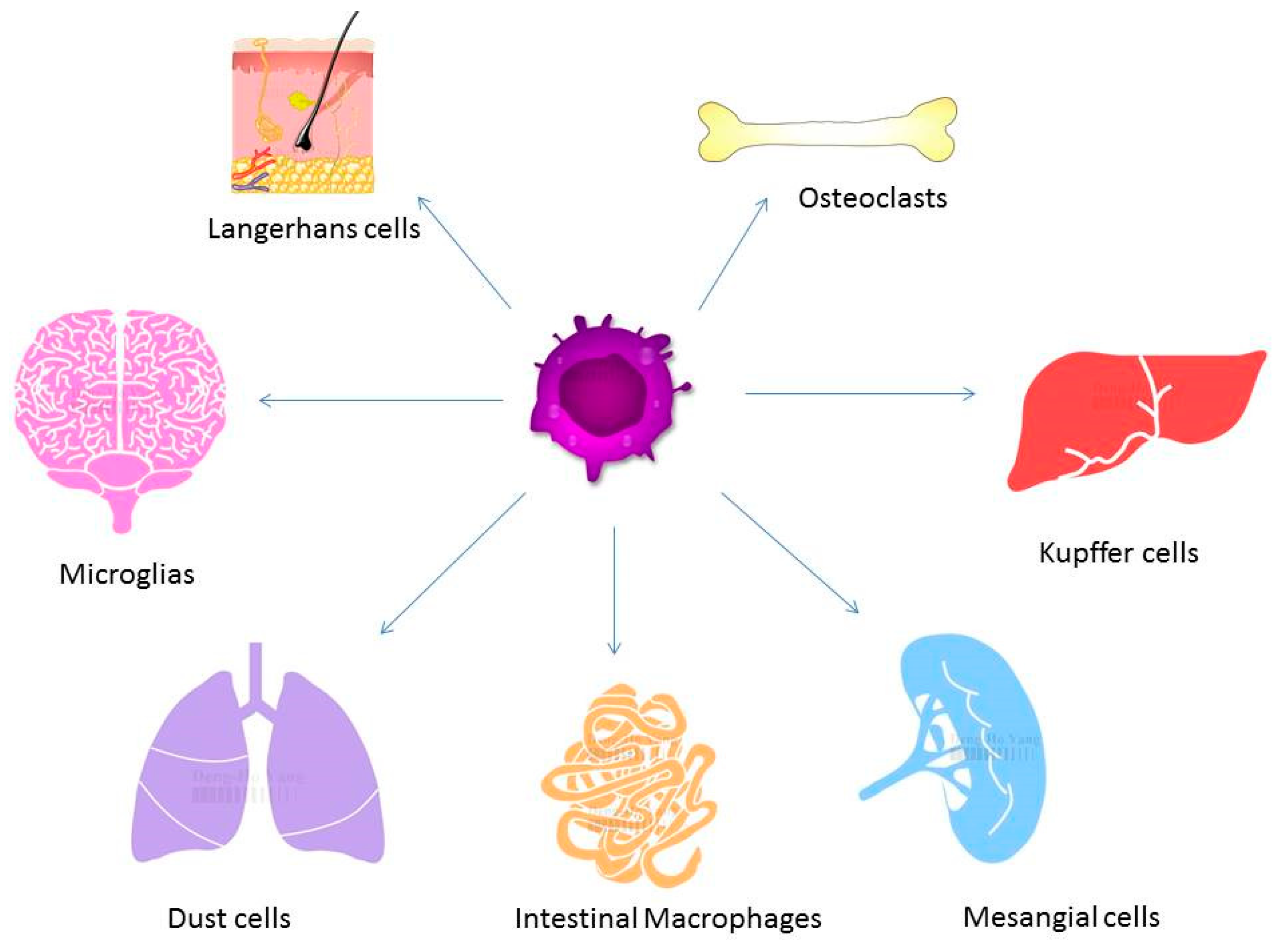
3. Macrophages in the Pathogenesis of Osteoporosis
4. The Cytokines from Macrophages Contribute to the Process of Osteoporosis
5. The Fusion of Macrophages/Monocytes to Form Multinucleated Cells—Osteoclasts
6. Different Cytokines for the Pathogenesis of Osteoporosis
7. Estrogen Deficiency Induced the Expression of Different Cytokines in Osteoporosis
8. The Activation and Differentiation of Macrophages to Osteoclasts in the Development of Osteoporosis
9. The Effect of Macrophages in Osteoblasts
10. The Activation and Regulation of Chemokines in Macrophages-Associated Osteoporosis
11. The Effect of Macrophages in Osteocytes
12. The Modulation of Insulin-Like Growth Factor-1 (IGF-1) in Macrophages
13. The Possible Therapy of Osteoporosis by Targeting Macrophages
14. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RANK | Receptor activator of NF-κB |
| RANKL | Receptor activator of NF-κB ligand |
| M-CSF | Macrophage-colony stimulating factor |
| TGF-β | Transforming growth factor-β |
| IFN-γ | Interferon-γ |
| TNF-α | Tumor necrosis factor-α |
| OPG | Osteoprotegerin |
| PTH | Parathyroid hormone |
| RA | Rheumatoid arthritis |
| TRAF | Tumor necrosis receptor-associated factor |
References
- Hadji, P.; Klein, S.; Gothe, H.; Haussler, B.; Kless, T.; Schmidt, T.; Steinle, T.; Verheyen, F.; Linder, R. The epidemiology of osteoporosis—bone evaluation study (best): An analysis of routine health insurance data. Dtsch. Arztebl. Int. 2013, 110, 52–57. [Google Scholar] [PubMed]
- Cunha-Henriques, S.; Costa-Paiva, L.; Pinto-Neto, A.M.; Fonsechi-Carvesan, G.; Nanni, L.; Morais, S.S. Postmenopausal women with osteoporosis and musculoskeletal status: A comparative cross-sectional study. J. Clin. Med. Res. 2011, 3, 168–176. [Google Scholar] [CrossRef][Green Version]
- Johnell, O.; Kanis, J. Epidemiology of osteoporotic fractures. Osteoporos. Int. 2005, 16, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef]
- Kassem, M.; Kveiborg, M.; Eriksen, E.F. Production and action of transforming growth factor-beta in human osteoblast cultures: Dependence on cell differentiation and modulation by calcitriol. Eur. J. Clin. Investig. 2000, 30, 429–437. [Google Scholar] [CrossRef]
- Jilka, R.L.; Weinstein, R.S.; Bellido, T.; Parfitt, A.M.; Manolagas, S.C. Osteoblast programmed cell death (apoptosis): Modulation by growth factors and cytokines. J. Bone Miner. Res. 1998, 13, 793–802. [Google Scholar] [CrossRef]
- Lucas, P.A. Chemotactic response of osteoblast-like cells to transforming growth factor beta. Bone 1989, 10, 459–463. [Google Scholar] [CrossRef]
- Kasagi, S.; Chen, W. Tgf-beta1 on osteoimmunology and the bone component cells. Cell Biosci. 2013, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Zupan, J.; Jeras, M.; Marc, J. Osteoimmunology and the influence of pro-inflammatory cytokines on osteoclasts. Biochem. Med. (Zagreb) 2013, 23, 43–63. [Google Scholar] [CrossRef]
- Baron, R.; Rawadi, G. Targeting the wnt/beta-catenin pathway to regulate bone formation in the adult skeleton. Endocrinology 2007, 148, 2635–2643. [Google Scholar] [CrossRef]
- Canalis, E.; Giustina, A.; Bilezikian, J.P. Mechanisms of anabolic therapies for osteoporosis. N. Engl. J. Med. 2007, 357, 905–916. [Google Scholar] [CrossRef]
- Poole, K.E.; van Bezooijen, R.L.; Loveridge, N.; Hamersma, H.; Papapoulos, S.E.; Lowik, C.W.; Reeve, J. Sclerostin is a delayed secreted product of osteocytes that inhibits bone formation. FASEB J. 2005, 19, 1842–1844. [Google Scholar] [CrossRef]
- Long, C.L.; Humphrey, M.B. Osteoimmunology: The expanding role of immunoreceptors in osteoclasts and bone remodeling. Bonekey Rep. 2012, 1. [Google Scholar] [CrossRef]
- Kylmaoja, E.; Nakamura, M.; Tuukkanen, J. Osteoclasts and remodeling based bone formation. Curr. Stem. Cell Res. Ther. 2016, 11, 626–633. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem Biophys 2008, 473, 201–209. [Google Scholar] [CrossRef]
- Horwood, N.J. Macrophage polarization and bone formation: A review. Clin. Rev. Allergy Immunol. 2016, 51, 79–86. [Google Scholar] [CrossRef]
- Dou, C.; Ding, N.; Zhao, C.; Hou, T.; Kang, F.; Cao, Z.; Liu, C.; Bai, Y.; Dai, Q.; Ma, Q.; et al. Estrogen deficiency-mediated m2 macrophage osteoclastogenesis contributes to m1/m2 ratio alteration in ovariectomized osteoporotic mice. J. Bone Miner. Res. 2018, 33, 899–908. [Google Scholar] [CrossRef]
- Pereira, M.; Petretto, E.; Gordon, S.; Bassett, J.H.D.; Williams, G.R.; Behmoaras, J. Common signalling pathways in macrophage and osteoclast multinucleation. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef]
- Murray, P.J. Macrophage polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Dey, A.; Allen, J.; Hankey-Giblin, P.A. Ontogeny and polarization of macrophages in inflammation: Blood monocytes versus tissue macrophages. Front. Immunol. 2014, 5, 683. [Google Scholar] [CrossRef]
- Silva, M.T. When two is better than one: Macrophages and neutrophils work in concert in innate immunity as complementary and cooperative partners of a myeloid phagocyte system. J. Leukoc. Biol. 2010, 87, 93–106. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Dar, H.Y.; Mishra, P.K. Immunoporosis: Immunology of osteoporosis-role of t cells. Front. Immunol. 2018, 9, 657. [Google Scholar] [CrossRef]
- Yun, T.J.; Chaudhary, P.M.; Shu, G.L.; Frazer, J.K.; Ewings, M.K.; Schwartz, S.M.; Pascual, V.; Hood, L.E.; Clark, E.A. Opg/fdcr-1, a tnf receptor family member, is expressed in lymphoid cells and is up-regulated by ligating cd40. J. Immunol. 1998, 161, 6113–6121. [Google Scholar]
- Jung, Y.K.; Kang, Y.M.; Han, S. Osteoclasts in the inflammatory arthritis: Implications for pathologic osteolysis. Immune Netw. 2019, 19, e2. [Google Scholar] [CrossRef]
- Sims, N.A.; Green, J.R.; Glatt, M.; Schlict, S.; Martin, T.J.; Gillespie, M.T.; Romas, E. Targeting osteoclasts with zoledronic acid prevents bone destruction in collagen-induced arthritis. Arthritis Rheum. 2004, 50, 2338–2346. [Google Scholar] [CrossRef]
- Mansoori, M.N.; Shukla, P.; Kakaji, M.; Tyagi, A.M.; Srivastava, K.; Shukla, M.; Dixit, M.; Kureel, J.; Gupta, S.; Singh, D. Il-18bp is decreased in osteoporotic women: Prevents inflammasome mediated il-18 activation and reduces th17 differentiation. Sci. Rep. 2016, 6, 33680. [Google Scholar] [CrossRef]
- Maugeri, D.; Mamazza, C.; Lo Giudice, F.; Puglisi, N.; Muscoso, E.G.; Rizzotto, M.; Testai, M.; Bennati, E.; Lentini, A.; Panebianco, P. Interleukin-18 (il-18) and matrix metalloproteinase-9 (mmp-9) in post-menopausal osteoporosis. Arch. Gerontol. Geriatr. 2005, 40, 299–305. [Google Scholar] [CrossRef]
- Tang, M.; Tian, L.; Luo, G.; Yu, X. Interferon-gamma-mediated osteoimmunology. Front. Immunol 2018, 9, 1508. [Google Scholar] [CrossRef]
- Sato, K.; Suematsu, A.; Okamoto, K.; Yamaguchi, A.; Morishita, Y.; Kadono, Y.; Tanaka, S.; Kodama, T.; Akira, S.; Iwakura, Y.; et al. Th17 functions as an osteoclastogenic helper t cell subset that links t cell activation and bone destruction. J. Exp. Med. 2006, 203, 2673–2682. [Google Scholar] [CrossRef]
- Shukla, P.; Mansoori, M.N.; Singh, D. Efficacy of anti-il-23 monotherapy versus combination therapy with anti-il-17 in estrogen deficiency induced bone loss conditions. Bone 2018, 110, 84–95. [Google Scholar] [CrossRef]
- Khera, A.; Kanta, P.; Kalra, J.; Dumir, D.; M, T. Resveratrol restores the level of key inflammatory cytokines and rankl/opg ratio in the femur of rat osteoporosis model. J. Women Aging 2018, 1–13. [Google Scholar] [CrossRef]
- Woodward, J. Regulation of haematopoietic progenitor cell proliferation and survival: The involvement of the osteoblast. Cell Adh. Migr. 2010, 4, 4–6. [Google Scholar] [CrossRef][Green Version]
- Shukla, P.; Mansoori, M.N.; Kakaji, M.; Shukla, M.; Gupta, S.K.; Singh, D. Interleukin 27 (il-27) alleviates bone loss in estrogen-deficient conditions by induction of early growth response-2 gene. J. Biol. Chem. 2017, 292, 4686–4699. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of rankl/rank/opg in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef]
- Udagawa, N.; Takahashi, N.; Akatsu, T.; Tanaka, H.; Sasaki, T.; Nishihara, T.; Koga, T.; Martin, T.J.; Suda, T. Origin of osteoclasts: Mature monocytes and macrophages are capable of differentiating into osteoclasts under a suitable microenvironment prepared by bone marrow-derived stromal cells. Proc. Natl. Acad. Sci. USA 1990, 87, 7260–7264. [Google Scholar] [CrossRef]
- Kao, W.J.; McNally, A.K.; Hiltner, A.; Anderson, J.M. Role for interleukin-4 in foreign-body giant cell formation on a poly(etherurethane urea) in vivo. J. Biomed. Mater. Res. 1995, 29, 1267–1275. [Google Scholar] [CrossRef]
- DeFife, K.M.; Jenney, C.R.; McNally, A.K.; Colton, E.; Anderson, J.M. Interleukin-13 induces human monocyte/macrophage fusion and macrophage mannose receptor expression. J. Immunol. 1997, 158, 3385–3390. [Google Scholar]
- DeFife, K.M.; Jenney, C.R.; Colton, E.; Anderson, J.M. Cytoskeletal and adhesive structural polarizations accompany il-13-induced human macrophage fusion. J. Histochem. Cytochem. 1999, 47, 65–74. [Google Scholar] [CrossRef]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. Opgl is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef]
- Khosla, S. Minireview: The opg/rankl/rank system. Endocrinology 2001, 142, 5050–5055. [Google Scholar] [CrossRef]
- Garnero, P.; Darte, C.; Delmas, P.D. A model to monitor the efficacy of alendronate treatment in women with osteoporosis using a biochemical marker of bone turnover. Bone 1999, 24, 603–609. [Google Scholar] [CrossRef]
- Greenblatt, M.B.; Shim, J.H. Osteoimmunology: A brief introduction. Immune Netw. 2013, 13, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Mann, G.N.; Jacobs, T.W.; Buchinsky, F.J.; Armstrong, E.C.; Li, M.; Ke, H.Z.; Ma, Y.F.; Jee, W.S.; Epstein, S. Interferon-gamma causes loss of bone volume in vivo and fails to ameliorate cyclosporin a-induced osteopenia. Endocrinology 1994, 135, 1077–1083. [Google Scholar] [CrossRef]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of osteoclast differentiation by cytokine networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef]
- Kong, Y.Y.; Feige, U.; Sarosi, I.; Bolon, B.; Tafuri, A.; Morony, S.; Capparelli, C.; Li, J.; Elliott, R.; McCabe, S.; et al. Activated t cells regulate bone loss and joint destruction in adjuvant arthritis through osteoprotegerin ligand. Nature 1999, 402, 304–309. [Google Scholar] [CrossRef]
- Geusens, P.; Lems, W.F. Osteoimmunology and osteoporosis. Arthritis Res. Ther. 2011, 13, 242. [Google Scholar] [CrossRef]
- Manabe, N.; Kawaguchi, H.; Chikuda, H.; Miyaura, C.; Inada, M.; Nagai, R.; Nabeshima, Y.; Nakamura, K.; Sinclair, A.M.; Scheuermann, R.H.; et al. Connection between b lymphocyte and osteoclast differentiation pathways. J. Immunol. 2001, 167, 2625–2631. [Google Scholar] [CrossRef]
- Nemeth, K.; Schoppet, M.; Al-Fakhri, N.; Helas, S.; Jessberger, R.; Hofbauer, L.C.; Goettsch, C. The role of osteoclast-associated receptor in osteoimmunology. J. Immunol. 2011, 186, 13–18. [Google Scholar] [CrossRef]
- Kameda, T.; Mano, H.; Yuasa, T.; Mori, Y.; Miyazawa, K.; Shiokawa, M.; Nakamaru, Y.; Hiroi, E.; Hiura, K.; Kameda, A.; et al. Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts. J. Exp. Med. 1997, 186, 489–495. [Google Scholar] [CrossRef]
- Kim, H.; Tabata, A.; Tomoyasu, T.; Ueno, T.; Uchiyama, S.; Yuasa, K.; Tsuji, A.; Nagamune, H. Estrogen stimuli promote osteoblastic differentiation via the subtilisin-like proprotein convertase pace4 in mc3t3-e1 cells. J. Bone Miner. Metab. 2014. [Google Scholar] [CrossRef]
- Stepan, J.J.; Alenfeld, F.; Boivin, G.; Feyen, J.H.; Lakatos, P. Mechanisms of action of antiresorptive therapies of postmenopausal osteoporosis. Endocr. Regul. 2003, 37, 225–238. [Google Scholar]
- Gao, J.; Liu, Q.; Liu, X.; Ji, C.; Qu, S.; Wang, S.; Luo, Y. Cyclin g2 suppresses estrogen-mediated osteogenesis through inhibition of wnt/beta-catenin signaling. PLoS ONE 2014, 9, e89884. [Google Scholar]
- Cenci, S.; Toraldo, G.; Weitzmann, M.N.; Roggia, C.; Gao, Y.; Qian, W.P.; Sierra, O.; Pacifici, R. Estrogen deficiency induces bone loss by increasing t cell proliferation and lifespan through ifn-gamma-induced class ii transactivator. Proc. Natl. Acad. Sci. USA 2003, 100, 10405–10410. [Google Scholar] [CrossRef]
- D’Amelio, P.; Isaia, G.C. Immune system and postmenopausal bone loss. Clinic Rev. Bone Miner. Metab 2009, 7, 262–268. [Google Scholar] [CrossRef]
- Zhao, R. Immune regulation of osteoclast function in postmenopausal osteoporosis: A critical interdisciplinary perspective. Int. J. Med. Sci. 2012, 9, 825–832. [Google Scholar] [CrossRef]
- Chin, K.Y. The relationship between follicle-stimulating hormone and bone health: Alternative explanation for bone loss beyond oestrogen? Int. J. Med. Sci. 2018, 15, 1373–1383. [Google Scholar] [CrossRef]
- Suzuki, R.; Fukami, S.; Tomomura, M.; Tomomura, A.; Shirataki, Y. Screening for natural medicines effective for the treatment of osteoporosis. J. Nat. Med. 2019, 73, 331–337. [Google Scholar] [CrossRef]
- Jin, H.; Yao, L.; Chen, K.; Liu, Y.; Wang, Q.; Wang, Z.; Liu, Q.; Cao, Z.; Kenny, J.; Tickner, J.; et al. Evodiamine inhibits rankl-induced osteoclastogenesis and prevents ovariectomy-induced bone loss in mice. J. Cell Mol. Med. 2019, 23, 522–534. [Google Scholar] [CrossRef]
- Kim, B.; Lee, K.Y.; Park, B. Icariin abrogates osteoclast formation through the regulation of the rankl-mediated traf6/nf-kappab/erk signaling pathway in raw264.7 cells. Phytomedicine 2018, 51, 181–190. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Ha, H.; Kim, R.; Cho, C.W.; Song, Y.R.; Hong, H.D.; Kim, T. Protective effects of a polysaccharide ble0 isolated from barley leaf on bone loss in ovariectomized mice. Int. J. Biol. Macromol. 2019, 123, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Li, J.; Xing, L.; Yao, Z. Bone remodeling and the role of traf3 in osteoclastic bone resorption. Front. Immunol. 2018, 9, 2263. [Google Scholar] [CrossRef]
- Bishop, G.A.; Stunz, L.L.; Hostager, B.S. Traf3 as a multifaceted regulator of b lymphocyte survival and activation. Front. Immunol. 2018, 9, 2161. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Okinaga, T.; Hikiji, H.; Ariyoshi, W.; Yoshiga, D.; Habu, M.; Tominaga, K.; Nishihara, T. Zoledronic acid exacerbates inflammation through m1 macrophage polarization. Inflamm. Regen. 2018, 38, 16. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Lee, Y.; Hwang, H.G.; Sung, S.H.; Lee, M.; Son, Y.J. Betulin suppresses osteoclast formation via down-regulating nfatc1. J. Clin. Med. 2018, 7, 154. [Google Scholar] [CrossRef]
- Vi, L.; Baht, G.S.; Soderblom, E.J.; Whetstone, H.; Wei, Q.; Furman, B.; Puviindran, V.; Nadesan, P.; Foster, M.; Poon, R.; et al. Macrophage cells secrete factors including lrp1 that orchestrate the rejuvenation of bone repair in mice. Nat. Commun. 2018, 9, 5191. [Google Scholar] [CrossRef]
- Lind, M.; Deleuran, B.; Yssel, H.; Fink-Eriksen, E.; Thestrup-Pedersen, K. Il-4 and il-13, but not il-10, are chemotactic factors for human osteoblasts. Cytokine 1995, 7, 78–82. [Google Scholar] [CrossRef]
- Silfversward, C.J.; Frost, A.; Brandstrom, H.; Nilsson, O.; Ljunggren, O. Interleukin-4 and interleukin-13 potentiate interleukin-1 induced secretion of interleukin-6 in human osteoblast-like cells. J. Orthop. Res. 2004, 22, 1058–1062. [Google Scholar] [CrossRef]
- Palmqvist, P.; Lundberg, P.; Persson, E.; Johansson, A.; Lundgren, I.; Lie, A.; Conaway, H.H.; Lerner, U.H. Inhibition of hormone and cytokine-stimulated osteoclastogenesis and bone resorption by interleukin-4 and interleukin-13 is associated with increased osteoprotegerin and decreased rankl and rank in a stat6-dependent pathway. J. Biol. Chem. 2006, 281, 2414–2429. [Google Scholar] [CrossRef]
- Panagakos, F.S. Transforming growth factor--alpha stimulates chemotaxis of osteoblasts and osteoblast-like cells in vitro. Biochem. Mol. Biol. Int. 1994, 33, 643–650. [Google Scholar] [PubMed]
- Harmer, D.; Falank, C.; Reagan, M.R. Interleukin-6 interweaves the bone marrow microenvironment, bone loss, and multiple myeloma. Front. Endocrinol. (Lausanne) 2018, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Meednu, N.; Rosenberg, A.; Rangel-Moreno, J.; Wang, V.; Glanzman, J.; Owen, T.; Zhou, X.; Zhang, H.; Boyce, B.F.; et al. B cells inhibit bone formation in rheumatoid arthritis by suppressing osteoblast differentiation. Nat. Commun. 2018, 9, 5127. [Google Scholar] [CrossRef] [PubMed]
- Pathak, J.L.; Bakker, A.D.; Verschueren, P.; Lems, W.F.; Luyten, F.P.; Klein-Nulend, J.; Bravenboer, N. Cxcl8 and ccl20 enhance osteoclastogenesis via modulation of cytokine production by human primary osteoblasts. PLoS ONE 2015, 10, e0131041. [Google Scholar] [CrossRef]
- Collins, F.L.; Williams, J.O.; Bloom, A.C.; Singh, R.K.; Jordan, L.; Stone, M.D.; McCabe, L.R.; Wang, E.C.Y.; Williams, A.S. Ccl3 and mmp-9 are induced by tl1a during death receptor 3 (tnfrsf25)-dependent osteoclast function and systemic bone loss. Bone 2017, 97, 94–104. [Google Scholar] [CrossRef]
- Xuan, W.; Feng, X.; Qian, C.; Peng, L.; Shi, Y.; Xu, L.; Wang, F.; Tan, W. Osteoclast differentiation gene expression profiling reveals chemokine ccl4 mediates rankl-induced osteoclast migration and invasion via pi3k pathway. Cell Biochem. Funct. 2017, 35, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Song, C.; Wang, Y.; Lei, Z.; Xu, F.; Guan, H.; Chen, A.; Li, F. Inhibition of prmt5 suppresses osteoclast differentiation and partially protects against ovariectomy-induced bone loss through downregulation of cxcl10 and rsad2. Cell Signal. 2017, 34, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Lee, S.; Knoll, J.; Rauch, A.; Ostermay, S.; Luther, J.; Malkusch, N.; Lerner, U.H.; Zaiss, M.M.; Neven, M.; et al. Loss of menin in osteoblast lineage affects osteocyte-osteoclast crosstalk causing osteoporosis. Cell Death Differ. 2017, 24, 672–682. [Google Scholar] [CrossRef]
- Koizumi, K.; Saitoh, Y.; Minami, T.; Takeno, N.; Tsuneyama, K.; Miyahara, T.; Nakayama, T.; Sakurai, H.; Takano, Y.; Nishimura, M.; et al. Role of cx3cl1/fractalkine in osteoclast differentiation and bone resorption. J. Immunol. 2009, 183, 7825–7831. [Google Scholar] [CrossRef]
- Zimmerman, S.M.; Heard-Lipsmeyer, M.E.; Dimori, M.; Thostenson, J.D.; Mannen, E.M.; O’Brien, C.A.; Morello, R. Loss of rankl in osteocytes dramatically increases cancellous bone mass in the osteogenesis imperfecta mouse (oim). Bone Rep. 2018, 9, 61–73. [Google Scholar] [CrossRef]
- Komori, T. Mouse models for the evaluation of osteocyte functions. J. Bone Metab. 2014, 21, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Wijenayaka, A.R.; Prideaux, M.; Kogawa, M.; Ormsby, R.T.; Evdokiou, A.; Bonewald, L.F.; Findlay, D.M.; Atkins, G.J. Regulation of fgf23 expression in idg-sw3 osteocytes and human bone by pro-inflammatory stimuli. Mol. Cell Endocrinol. 2015, 399, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Mei, J.; Yu, Z.; Zhai, Z.; Qiao, H.; Dai, K. Lenalidomide regulates osteocytes fate and related osteoclastogenesis via il-1beta/nf-kappab/rankl signaling. Biochem. Biophys. Res. Commun. 2018, 501, 547–555. [Google Scholar] [CrossRef]
- McGregor, N.E.; Murat, M.; Elango, J.; Poulton, I.J.; Walker, E.C.; Crimeen-Irwin, B.; Ho, P.W.M.; Gooi, J.H.; Martin, T.J.; Sims, N.A. Il-6 exhibits both cis and trans signaling in osteocytes and osteoblasts, but only trans signaling promotes bone formation and osteoclastogenesis. J. Biol. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Choi, Y.; Bae, M.K.; Hwang, D.S.; Shin, S.H.; Lee, J.Y. Zoledronate enhances osteocyte-mediated osteoclast differentiation by il-6/rankl axis. Int. J. Mol. Sci. 2019, 20, 1467. [Google Scholar] [CrossRef]
- Metzger, C.E.; Gong, S.; Aceves, M.; Bloomfield, S.A.; Hook, M.A. Osteocytes reflect a pro-inflammatory state following spinal cord injury in a rodent model. Bone 2019, 120, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Chhana, A.; Pool, B.; Callon, K.E.; Tay, M.L.; Musson, D.; Naot, D.; McCarthy, G.; McGlashan, S.; Cornish, J.; Dalbeth, N. Monosodium urate crystals reduce osteocyte viability and indirectly promote a shift in osteocyte function towards a proinflammatory and proresorptive state. Arthritis Res. Ther. 2018, 20, 208. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Lee, J.; Byeon, J.S.; Jeong, D.U.; Gu, N.Y.; Cho, I.S.; Cha, S.H. Effects of three-dimensional spheroid culture on equine mesenchymal stem cell plasticity. Vet. Res. Commun. 2018, 42, 171–181. [Google Scholar] [CrossRef] [PubMed]
- de Barros Silva, P.G.; de Oliveira, C.C.; Brizeno, L.; Wong, D.; Lima Junior, R.; Goncalves, R.P.; Sousa, F.B.; Mota, M.; de Albuquerque Ribeiro, R.; Alves, A. Immune cellular profile of bisphosphonate-related osteonecrosis of the jaw. Oral Dis. 2016, 22, 649–657. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, C.; Yang, Y.; Yang, H.; Chen, J. Osteocyte-intrinsic mtorc1 signaling restrains trabecular bone accrual in mice. J. Cell Biochem. 2018, 119, 8743–8749. [Google Scholar] [CrossRef]
- Siegenthaler, B.; Ghayor, C.; Gjoksi-Cosandey, B.; Ruangsawasdi, N.; Weber, F.E. The bromodomain inhibitor n-methyl pyrrolidone prevents osteoporosis and bmp-triggered sclerostin expression in osteocytes. Int. J. Mol. Sci. 2018, 19, 3332. [Google Scholar] [CrossRef]
- Taylor, S.; Hu, R.; Pacheco, E.; Locher, K.; Pyrah, I.; Ominsky, M.S.; Boyce, R.W. Differential time-dependent transcriptional changes in the osteoblast lineage in cortical bone associated with sclerostin antibody treatment in ovariectomized rats. Bone Rep. 2018, 8, 95–103. [Google Scholar] [CrossRef]
- Boyce, R.W.; Brown, D.; Felx, M.; Mellal, N.; Locher, K.; Pyrah, I.; Ominsky, M.S.; Taylor, S. Decreased osteoprogenitor proliferation precedes attenuation of cancellous bone formation in ovariectomized rats treated with sclerostin antibody. Bone Rep. 2018, 8, 90–94. [Google Scholar] [CrossRef]
- Toscani, D.; Bolzoni, M.; Ferretti, M.; Palumbo, C.; Giuliani, N. Role of osteocytes in myeloma bone disease: Anti-sclerostin antibody as new therapeutic strategy. Front. Immunol. 2018, 9, 2467. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, X.; Li, B.; Jiang, S.; Jiang, L. Increased activity of osteocyte autophagy in ovariectomized rats and its correlation with oxidative stress status and bone loss. Biochem. Biophys. Res. Commun. 2014, 451, 86–92. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; Sasso, G.R.; Simoes, M.J.; Simoes, R.S.; Baracat, M.C.; Sasso-Cerri, E.; Cerri, P.S. Osteoporosis and autophagy: What is the relationship? Rev. Assoc. Med. Bras. 2017, 63, 173–179. [Google Scholar] [CrossRef]
- Ye, T.; Cao, P.; Qi, J.; Zhou, Q.; Rao, D.S.; Qiu, S. Protective effect of low-dose risedronate against osteocyte apoptosis and bone loss in ovariectomized rats. PLoS ONE 2017, 12, e0186012. [Google Scholar] [CrossRef]
- Reis, L.C.; Ramos-Sanchez, E.M.; Petitto-Assis, F.; Nerland, A.H.; Hernandez-Valladares, M.; Selheim, F.; Floeter-Winter, L.M.; Goto, H. Insulin-like growth factor-i as an effector element of the cytokine il-4 in the development of a leishmania major infection. Mediators Inflamm. 2018, 2018, 9787128. [Google Scholar] [CrossRef]
- Sun, K.T.; Cheung, K.K.; Au, S.W.N.; Yeung, S.S.; Yeung, E.W. Overexpression of mechano-growth factor modulates inflammatory cytokine expression and macrophage resolution in skeletal muscle injury. Front. Physiol. 2018, 9, 999. [Google Scholar] [CrossRef]
- Wang, J.; Xie, L.; Wang, S.; Lin, J.; Liang, J.; Xu, J. Azithromycin promotes alternatively activated macrophage phenotype in systematic lupus erythematosus via pi3k/akt signaling pathway. Cell Death Dis. 2018, 9, 1080. [Google Scholar] [CrossRef]
- Tonkin, J.; Temmerman, L.; Sampson, R.D.; Gallego-Colon, E.; Barberi, L.; Bilbao, D.; Schneider, M.D.; Musaro, A.; Rosenthal, N. Monocyte/macrophage-derived igf-1 orchestrates murine skeletal muscle regeneration and modulates autocrine polarization. Mol. Ther. 2015, 23, 1189–1200. [Google Scholar] [CrossRef]
- Zhao, G.; Monier-Faugere, M.C.; Langub, M.C.; Geng, Z.; Nakayama, T.; Pike, J.W.; Chernausek, S.D.; Rosen, C.J.; Donahue, L.R.; Malluche, H.H.; et al. Targeted overexpression of insulin-like growth factor i to osteoblasts of transgenic mice: Increased trabecular bone volume without increased osteoblast proliferation. Endocrinology 2000, 141, 2674–2682. [Google Scholar] [CrossRef]
- Grinspoon, S.; Baum, H.; Lee, K.; Anderson, E.; Herzog, D.; Klibanski, A. Effects of short-term recombinant human insulin-like growth factor i administration on bone turnover in osteopenic women with anorexia nervosa. J. Clin. Endocrinol. Metab. 1996, 81, 3864–3870. [Google Scholar] [PubMed]
- Gao, Y.; Wu, X.; Terauchi, M.; Li, J.Y.; Grassi, F.; Galley, S.; Yang, X.; Weitzmann, M.N.; Pacifici, R. T cells potentiate pth-induced cortical bone loss through cd40l signaling. Cell Metab. 2008, 8, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Milstrey, A.; Wieskoetter, B.; Hinze, D.; Grueneweller, N.; Stange, R.; Pap, T.; Raschke, M.; Garcia, P. Dose-dependent effect of parathyroid hormone on fracture healing and bone formation in mice. J. Surg. Res. 2017, 220, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Lei, W.; Duan, R.; Li, Y.; Luo, L.; Boyce, B.F. Rankl cytokine enhances tnf-induced osteoclastogenesis independently of tnf receptor associated factor (traf) 6 by degrading traf3 in osteoclast precursors. J. Biol. Chem. 2017, 292, 10169–10179. [Google Scholar] [CrossRef]
- Suzuki, T.; Nakamura, Y.; Kato, H. Effects of denosumab on bone metabolism and bone mineral density with anti-tnf inhibitors, tocilizumab, or abatacept in osteoporosis with rheumatoid arthritis. Ther. Clin. Risk Manag. 2018, 14, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Tada, M.; Inui, K.; Sugioka, Y.; Mamoto, K.; Okano, T.; Koike, T. Abatacept might increase bone mineral density at femoral neck for patients with rheumatoid arthritis in clinical practice: Airtight study. Rheumatol. Int. 2018, 38, 777–784. [Google Scholar] [CrossRef]
- Zerbini, C.A.F.; Clark, P.; Mendez-Sanchez, L.; Pereira, R.M.R.; Messina, O.D.; Una, C.R.; Adachi, J.D.; Lems, W.F.; Cooper, C.; Lane, N.E. Biologic therapies and bone loss in rheumatoid arthritis. Osteoporos. Int. 2017, 28, 429–446. [Google Scholar] [CrossRef]
- Moon, S.J.; Ahn, I.E.; Jung, H.; Yi, H.; Kim, J.; Kim, Y.; Kwok, S.K.; Park, K.S.; Min, J.K.; Park, S.H.; et al. Temporal differential effects of proinflammatory cytokines on osteoclastogenesis. Int. J. Mol. Med. 2013, 31, 769–777. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.-H.; Yang, M.-Y. The Role of Macrophage in the Pathogenesis of Osteoporosis. Int. J. Mol. Sci. 2019, 20, 2093. https://doi.org/10.3390/ijms20092093
Yang D-H, Yang M-Y. The Role of Macrophage in the Pathogenesis of Osteoporosis. International Journal of Molecular Sciences. 2019; 20(9):2093. https://doi.org/10.3390/ijms20092093
Chicago/Turabian StyleYang, Deng-Ho, and Meng-Yin Yang. 2019. "The Role of Macrophage in the Pathogenesis of Osteoporosis" International Journal of Molecular Sciences 20, no. 9: 2093. https://doi.org/10.3390/ijms20092093
APA StyleYang, D.-H., & Yang, M.-Y. (2019). The Role of Macrophage in the Pathogenesis of Osteoporosis. International Journal of Molecular Sciences, 20(9), 2093. https://doi.org/10.3390/ijms20092093