Xenobiotics Formed during Food Processing: Their Relation with the Intestinal Microbiota and Colorectal Cancer

 , and
, and
Abstract
1. Introduction
1.1. Impact of Diet on Colorectal Cancer
1.2. Intestinal Microbiota and Human Health
2. Food Processing and Xenobiotics
3. Effect of Food Processing-Borne Xenobiotics on the Gut Microbiota
3.1. Impact of Xenobiotics on Gut Microbiota
3.2. Impact of the Gut Microbiota on the Toxicity of Xenobiotics
4. Creating a Balance between Xenobiotics and a Healthy Gut
5. Future Perspectives
5.1. Probiotics and Prebiotics to Counteract the Effect of Pro-Carcinogenic Compounds
5.2. Longitudinal Studies on Long Term Impact of Xenobiotics Derived from Food Processing
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACs | Aminocarbolines |
| AIAs | Aminoimidazoazarenes |
| ATP | Adenosine triphosphate |
| B(a)P | Benzo(a) pyrene |
| CI | Confidence interval |
| CRC | Colorectal cancer |
| CYP1A1 | Aryl-4 monooxigenase |
| CYP450 | Cytochrome P450 |
| DiMeIQx | 2-Amino-3,4,8-trimethylimidazo[4,5-f] quinoxaline |
| EPIC | European Prospective Investigation into Cancer |
| HCAs | Heterocyclic amines |
| IARC | The International Agency for Research on Cancer |
| LAB | Lactic acid bacteria |
| MD | Mediterranean Diet |
| MeIQ | 2-Amino-3,4-dimethylimidazo[4,5-f] quinoline |
| MeIQx | 2-Amino-3,8-dimethylimidazo[4,5-f] quinoxaline |
| MN | Micronucleous assay |
| MTT | (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) |
| NAs | Nitrosamines |
| NOCs | N-Nitroso compounds |
| OECD | Organization for Economic Cooperation and Development |
| PAHs | Polycyclic aromatic hydrocarbons |
| PhIP | 2-Amino-1-methyl-6-phenylimidazo[4,5-b] pyridine |
| ROS | Reactive oxygen species |
| RR | Relative risk |
| SCE | Sister Chromatid Exchange assay |
| SCFA | Short chain fatty acids |
| WD | Western Diet |
References
- Viera, C.; Rodrigues, M.; Russo, E.; Amedei, A. Role of diet and gut microbiota on colorectal cancer immunomodulation. World J. Gastroenterol. 2019, 25, 151–162. [Google Scholar] [CrossRef]
- Cunnane, S.C. Origins and evolution of the Western diet: Implications of iodine and seafood intakes for the human brain. Am. J. Clin. Nutr. 2005, 82, 483. [Google Scholar] [CrossRef][Green Version]
- Naja, F.; Hwalla, N.; Itani, L.; Karam, S.; Mehio Sibai, A.; Nasreddine, L. A Western dietary pattern is associated with overweight and obesity in a national sample of Lebanese adolescents (13–19 years): A cross-sectional study. Br. J. Nutr. 2015, 114, 1909–1919. [Google Scholar] [CrossRef]
- Di Daniele, N.; Noce, A.; Vidiri, M.F.; Moriconi, E.; Marrone, G.; Annicchiarico-Petruzzelli, M.; D’Urso, G.; Tesauro, M.; Rovella, V.; De Lorenzo, A. Impact of Mediterranean diet on metabolic syndrome, cancer and longevity. Oncotarget 2017, 8, 8947–8979. [Google Scholar] [CrossRef] [PubMed]
- Farinetti, A.; Zurlo, V.; Manenti, A.; Coppi, F.; Mattioli, A. Mediterranean diet and colorectal cancer: A systematic review. Nutrition 2017, 43–44, 82–88. [Google Scholar] [CrossRef]
- Liu, L.; Zhuang, W.; Wang, R.-Q.; Mukherjee, R.; Xiao, S.-M.; Chen, Z.; Wu, X.-T.; Zhou, Y.; Zhang, H.-Y. Is dietary fat associated with the risk of colorectal cancer? A meta-analysis of 13 prospective cohort studies. Eur. J. Nutr. 2011, 50, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Beresford, S.A.A.; Johnson, K.C.; Ritenbaugh, C.; Lasser, N.L.; Snetselaar, L.G.; Black, H.R.; Anderson, G.L.; Assaf, A.R.; Bassford, T.; Bowen, D.; et al. Low-fat dietary pattern and risk of colorectal cancer. JAMA 2006, 295, 643–654. [Google Scholar] [CrossRef]
- Vieira, A.R.; Abar, L.; Chan, D.S.M.; Vingeliene, S.; Polemiti, E.; Stevens, C.; Greenwood, D.; Norat, T. Foods and beverages and colorectal cancer risk: A systematic review and meta-analysis of cohort studies, an update of the evidence of the WCRF-AICR Continuous Update Project. Ann. Oncol. 2017, 28, 1788–1802. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Barupal, D.K.; Rothwell, J.A.; Jenab, M.; Fedirko, V.; Romieu, I.; Aleksandrova, K.; Overvad, K.; Kyrø, C.; Tjønneland, A.; et al. Dietary flavonoid intake and colorectal cancer risk in the European prospective investigation into cancer and nutrition (EPIC) cohort. Int. J. Cancer 2017, 140, 1836–1844. [Google Scholar] [CrossRef]
- De Stefani, E.; Boffetta, P.; Deneo-Pellegrini, H.; Ronco, A.L.; Aune, D.; Acosta, G.; Brennan, P.; Mendilaharsu, M.; Ferro, G. Meat intake, meat mutagens and risk of lung cancer in Uruguayan men. Cancer Causes Control. 2009, 20, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Meat and cancer. Meat Sci. 2010, 84, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Wie, G.-A.; Cho, Y.-A.; Kang, H.; Ryu, K.-A.; Yoo, M.-K.; Kim, Y.-A.; Jung, K.-W.; Kim, J.; Lee, J.-H.; Joung, H. Red meat consumption is associated with an increased overall cancer risk: A prospective cohort study in Korea. Br. J. Nutr. 2014, 112, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K.; Stewart, B.W.; et al. Carcinogenicity of consumption of red and processed meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef]
- Aune, D.; Chan, D.S.M.; Vieira, A.R.; Navarro Rosenblatt, D.A.; Vieira, R.; Greenwood, D.C.; Kampman, E.; Norat, T. Red and processed meat intake and risk of colorectal adenomas: A systematic review and meta-analysis of epidemiological studies. Cancer Causes Control. 2013, 24, 611–627. [Google Scholar] [CrossRef]
- Chiang, V.S.C.; Quek, S.Y. The relationship of red meat with cancer: Effects of thermal processing and related physiological mechanisms. Crit. Rev. Food Sci. Nutr. 2017, 57, 1153–1173. [Google Scholar] [CrossRef] [PubMed]
- Catsburg, C.E.; Gago-Dominguez, M.; Yuan, J.M.; Castelao, J.E.; Cortessis, V.K.; Pike, M.C.; Stern, M.C. Dietary sources of N-nitroso compounds and bladder cancer risk: Findings from the Los Angeles bladder cancer study. Int. J. Cancer 2014, 134, 125–135. [Google Scholar] [CrossRef]
- Ministerio de Agricultura y Pesca Alimentación y Medio Ambiente (MAPAMA). Informe del Consumo de Alimentación en España; MAPAMA: Madrid, Spain, 2016; pp. 1–225.
- Hirose, M.; Takahashi, S.; Ogawa, K.; Futakuchi, M.; Shirai, T. Phenolics: Blocking agents for heterocyclic amine-induced carcinogenesis. Food Chem. Toxicol. 1999, 37, 985–992. [Google Scholar] [CrossRef]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLOS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Knights, D.; Ward, T.L.; McKinlay, C.E.; Miller, H.; Gonzalez, A.; McDonald, D.; Knight, R. Rethinking “Enterotypes” . Cell Host Microbe 2014, 16, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Mahurkar, A.; Rahnavard, G.; Crabtree, J.; Orvis, J.; Hall, A.B.; Brady, A.; Creasy, H.H.; McCracken, C.; Giglio, M.G.; et al. Strains, functions and dynamics in the expanded Human Microbiome Project. Nature 2017, 50, 61–66. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]
- Integrative HMP (iHMP) Research Network Consortium. The integrative human microbiome project: Dynamic analysis of microbiome-host omics profiles during periods of human health and disease. Cell Host Microbe 2014, 16, 276–289. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 6150–6172. [Google Scholar] [CrossRef]
- De Palma, G.; Lynch, M.D.J.; Lu, J.; Dang, V.T.; Deng, Y.; Jury, J.; Umeh, G.; Miranda, P.M.; Pigrau Pastor, M.; Sidani, S.; et al. Transplantation of fecal microbiota from patients with irritable bowel syndrome alters gut function and behavior in recipient mice. Sci. Transl. Med. 2017, 9, eaaf6397. [Google Scholar] [CrossRef]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Qin, J.; Balzola, F.; Bernstein, C.; Ho, G.T.; Lees, C.; Bork, P.; Dusko, E.; Wang, J. A human gut microbial gene catalogue established by metagenomic sequencing: Commentary. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Krajmalnik-Brown, R.; Lozupone, C.; Kang, D.-W.; Adams, J.B. Gut bacteria in children with autism spectrum disorders: Challenges and promise of studying how a complex community influences a complex disease. Microb. Ecol. Heal. Dis. 2015, 26, 26914. [Google Scholar] [CrossRef]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D. Thermal processing of food reduces gut microbiota diversity of the host and triggers adaptation of the microbiota: Evidence from two vertebrates. Microbiome 2018, 6, 99–113. [Google Scholar] [CrossRef]
- Katsidzira, L.; Laubscher, R.; Gangaidzo, I.T.; Swart, R.; Makunike-Mutasa, R.; Manyanga, T.; Thomson, S.; Ramesar, R.; Matenga, J.A.; Rusakaniko, S. Dietary patterns and colorectal cancer risk in Zimbabwe: A population based case-control study. Cancer Epidemiol. 2018, 57, 33–38. [Google Scholar] [CrossRef]
- Shirai, T.; Tamano, S.; Sano, M.; Masui, T.; Hasegawa, R.; Ito, N. Carcinogenicity of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) in rats: Dose-response studies. Princess Takamatsu Symp. 1995, 23, 232–239. [Google Scholar]
- Kim, E.; Coelho, D.; Blachier, F. Review of the association between meat consumption and risk of colorectal cancer. Nutr. Res. 2013, 33, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Bastide, N.; Pierre, F.; Corpet, D. Heme iron from meat and risk of colorectal cancer: A meta-analysis and a review of the mechanisms involved. Cancer Prev. Res. 2011, 4, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Viegas, O.; Novo, P.; Pinto, E.; Pinho, O.; Ferreira, I. Effect of charcoal types and grilling conditions on formation of heterocyclic aromatic amines (HAs) and polycyclic aromatic hydrocarbons (PAHs) in grilled muscle foods. Food Chem. Toxicol. 2012, 50, 2128–2134. [Google Scholar] [CrossRef] [PubMed]
- Chiavarini, M.; Bertarelli, G.; Minelli, L.; Fabiani, R. Dietary intake of meat cooking-related mutagens(HCAs) and risk of colorectal adenoma and cancer: A systematic review and meta-analysis. Nutrients 2017, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Martín-Calero, A.; Tejral, G.; Ayala, J.; González, V.; Afonso, A. Suitability of ionic liquids as mobile-phase additives in HPLC with fluorescence and UV detection for the determination of heterocyclic aromatic amines. J. Sep. Sci. 2010, 33, 182–190. [Google Scholar] [CrossRef]
- Etemadi, A.; Abnet, C.C.; Graubard, B.I.; Beane-Freeman, L.; Freedman, N.D.; Liao, L.; Dawsey, S.M.; Sinha, R. Anatomical subsite can modify the association between meat and meat compounds and risk of colorectal adenocarcinoma: Findings from three large US cohorts. Int. J. Cancer 2018, 143, 2261–2270. [Google Scholar] [CrossRef]
- Martínez Góngora, V.; Matthes, K.L.; Castaño, P.R.; Linseisen, J.; Rohrmann, S. Dietary heterocyclic amine intake and colorectal adenoma risk: A systematic review and meta-analysis. Cancer Epidemiol. Biomark. Prev. 2018, 28, 99–109. [Google Scholar] [CrossRef]
- Helmus, D.S.; Thompson, C.L.; Zelenskiy, S.; Tucker, T.C.; Li, L. Red Meat-derived heterocyclic amines increase risk of colon cancer: A population-based case-control study. Nutr. Cancer 2013, 65, 1141–1150. [Google Scholar] [CrossRef]
- Khan, M.; Busquets, R.; Santos, F.; Puignou, L. New method for the analysis of heterocyclic amines in meat extracts using pressurised liquid extraction and liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2008, 1194, 155–160. [Google Scholar] [CrossRef]
- Kondjoyan, A.; Chevolleau, S.; Portanguen, S.; Molina, J.; Ikonic, P.; Clerjon, S.; Debrauwer, L. Realation between crust development and heterocyclic aromatic amine formation when air-roasting a meat cylinder. Food Chem. 2016, 213, 641–646. [Google Scholar] [CrossRef]
- Buła, M.; Przybylski, W.; Jaworska, D.; Kajak-Siemaszko, K. Formation of heterocyclic aromatic amines in relation to pork quality and heat treatment parameters. Food Chem. 2019, 276, 511–516. [Google Scholar] [CrossRef]
- Rohrmann, S.; Nimptsch, K.; Sinha, R.; Willett, W.C.; Giovannucci, E.L.; Platz, E.A.; Wu, K. Intake of Meat mutagens and risk of prostate cancer in a cohort of U.S. health professionals. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Larsson, B.; Sahlberg, P.; Eriksson, A.; Busk, L. Polycyclic Aromatic Hydrocarbons in Grilled Food. J. Agric. Food Chem. 1983, 31, 867–873. [Google Scholar] [CrossRef]
- Alomirah, H.; Al-Zenki, S.; Al-Hooti, S.; Zaghloul, S.; Sawaya, W.; Ahmed, N.; Kannan, K. Concentrations and dietary exposure to polycyclic aromatic hydrocarbons (PAHs) from grilled and smoked foods. Food Control 2011, 22, 2028–2035. [Google Scholar] [CrossRef]
- Flores-Balcázar, C.; Rosales-Pérez, S.; Galván-Salazar, G.; López-Navarro, O. Nutrientes de la dieta y apoptosis como mecanismos reguladores del cáncer. iMedPub J. 2015, 11. [Google Scholar] [CrossRef]
- Pérez-Morales, G.; Morales, P.; Haza, A. Polycyclic aromatic hidrocarbons (PAHS) I: Toxicity, population exposure and involved foods. RCCV 2016, 10, 1–15. [Google Scholar] [CrossRef]
- Tamaki, A.; Hayashi, H.; Nakajima, H.; Takii, T.; Katagiri, D.; Miyazawa, K.; Hirose, K.; Onozaki, K. Polycyclic aromatic hydrocarbon increases mRNA level for interleukin 1 beta in human fibroblast-like synoviocyte line via aryl hydrocarbon receptor. Biol. Pharm. Bull. 2004, 27, 407–410. [Google Scholar] [CrossRef]
- Joshi, A.D.; Kim, A.; Lewinger, J.P.; Ulrich, C.M.; Potter, J.D.; Cotterchio, M.; Le Marchand, L.; Stern, M.C. Meat intake, cooking methods, dietary carcinogens, and colorectal cancer risk: Findings from the Colorectal Cancer Family Registry. Cancer Med. 2015, 4, 936–952. [Google Scholar] [CrossRef] [PubMed]
- Busquets, R.; Bordas, M.; Toribio, F.; Puignou, L.; Galceran, M.T. Occurrence of heterocyclic amines in several home-cooked meat dishes of the Spanish diet. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 802, 79–86. [Google Scholar] [CrossRef]
- Trichopoulou, A.; Martínez-González, M.A.; Tong, T.Y.N.; Forouhi, N.G.; Khandelwal, S.; Prabhakaran, D.; Mozaffarian, D.; de Lorgeril, M. Definitions and potential health benefits of the Mediterranean diet: Views from experts around the world. BMC Med. 2014, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Druzhinin, V.G.; Matskova, L.V.; Fucic, A. Induction and modulation of genotoxicity by the bacteriome in mammals. Mutat. Res. 2018, 776, 70–77. [Google Scholar] [CrossRef] [PubMed]
- De Kok, T. Evaluation of fecal mutagenicity and colorectal cancer risk. Mutat. Res. Mutat. Res. 2000, 463, 53–101. [Google Scholar] [CrossRef]
- Grasso, F.; Frisan, T. Bacterial genotoxins: Merging the DNA damage response into infection biology. Biomolecules 2015, 5, 1762–1782. [Google Scholar] [CrossRef] [PubMed]
- Jinadasa, R.N.; Bloom, S.E.; Weiss, R.S.; Duhamel, G.E. Cytolethal distending toxin: A conserved bacterial genotoxin that blocks cell cycle progression, leading to apoptosis of a broad range of mammalian cell lineages. Microbiology 2011, 157, 1851–1875. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, A.; Nadel, B.; Raoult, D.; Neefjes, J.; Gorvel, J.-P. Collateral damage: Insights into bacterial mechanisms that predispose host cells to cancer. Nat. Rev. Microbiol. 2017, 15, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Schauer, D.B.; Fox, J.G. In vivo virulence properties of bacterial cytolethal-distending toxin. Cell. Microbiol. 2008, 10, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Nougayrede, J.-P. Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science 2006, 313, 848–851. [Google Scholar] [CrossRef]
- Cougnoux, A.; Dalmasso, G.; Martinez, R.; Buc, E.; Delmas, J.; Gibold, L.; Sauvanet, P.; Darcha, C.; Déchelotte, P.; Bonnet, M.; et al. Bacterial genotoxin colibactin promotes colon tumour growth by inducing a senescence-associated secretory phenotype. Gut 2014, 63, 1932–1942. [Google Scholar] [CrossRef]
- Buc, E.; Dubois, D.; Sauvanet, P.; Raisch, J.; Delmas, J.; Darfeuille-Michaud, A.; Pezet, D.; Bonnet, R. High prevalence of mucosa-associated E. coli producing cyclomodulin and genotoxin in colon cancer. PLoS ONE 2013, 8, e56964. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.; Hogan, A.M.; Winter, D.C. Microbial and viral pathogens in colorectal cancer. Lancet Oncol. 2011, 12, 504–512. [Google Scholar] [CrossRef]
- Wu, S.; Morin, P.J.; Maouyo, D.; Sears, C.L. Bacteroides fragilis enterotoxin induces c-Myc expression and cellular proliferation. Gastroenterology 2003, 124, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its fadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, Y. Escherichia coli cytotoxic necrotizing factors and Bordetella dermonecrotic toxin: The dermonecrosis-inducing toxins activating Rho small GTPases. Toxicon 2001, 39, 1619–1627. [Google Scholar] [CrossRef]
- Nougayrède, J.-P.; Taieb, F.; De Rycke, J.; Oswald, E. Cyclomodulins: Bacterial effectors that modulate the eukaryotic cell cycle. Trends Microbiol. 2005, 13, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Barrasa, J.I.; Olmo, N.; Lizarbe, M.A.; Turnay, J. Bile acids in the colon, from healthy to cytotoxic molecules. Toxicol. In Vitro 2013, 27, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, H.; Chu, E.S.H.; Zhang, X.; Sheng, J.; Nakatsu, G.; Ng, S.C.; Chan, A.W.H.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. Peptostreptococcus anaerobius induces intracellular cholesterol biosynthesis in colon cells to induce proliferation and causes dysplasia in mice. Gastroenterology 2017, 152, 1419–1433. [Google Scholar] [CrossRef]
- Huycke, M.M.; Abrams, V.; Moore, D.R. Enterococcus faecalis produces extracellular superoxide and hydrogen peroxide that damages colonic epithelial cell DNA. Carcinogenesis 2002, 23, 529–536. [Google Scholar] [CrossRef]
- Owen, R.W. Generation of reactive oxygen species by the faecal matrix. Gut 2000, 46, 225–232. [Google Scholar] [CrossRef]
- Attene-Ramos, M.S. Evidence that hydrogen sulfide is a genotoxic agent. Mol. Cancer Res. 2006, 4, 9–14. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Lan, A.; Beaumont, M.; Audebert, M.; Wong, X.; Yamada, K.; Yin, Y.; Tomé, D.; Carrasco-Pozo, C.; Gotteland, M.; et al. The deleterious metabolic and genotoxic effects of the bacterial metabolite p-cresol on colonic epithelial cells. Free Radic. Biol. Med. 2015, 85, 219–227. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Andriamihaja, M.; Davila, A.-M.; Lan, A.; Grauso, M.; Armand, L.; Benamouzig, R.; Tomé, D. Changes in the luminal environment of the colonic epithelial cells and physiopathological consequences. Am. J. Pathol. 2017, 187, 476–486. [Google Scholar] [CrossRef]
- Szekely, J.; Gates, K.S. Noncovalent DNA binding and the mechanism of oxidative DNA damage by fecapentaene-12. Chem. Res. Toxicol. 2006, 19, 117–121. [Google Scholar] [CrossRef]
- Mouillé, B.; Delpal, S.; Mayeur, C.; Blachier, F. Inhibition of human colon carcinoma cell growth by ammonia: A non-cytotoxic process associated with polyamine synthesis reduction. Biochim. Biophys. Acta Gen. Subj. 2003, 1624, 88–97. [Google Scholar] [CrossRef]
- Federici, E.; Prete, R.; Lazzi, C.; Pellegrini, N.; Moretti, M.; Corsetti, A.; Cenci, G. Bacterial composition, genotoxicity, and cytotoxicity of fecal samples from individuals consuming omnivorous or vegetarian diets. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Daniela, E.; Sara, S.; Marcella, M.; Giovanni, A.; Meynier, A.; Sophie, V.; Cristina, C.M. Fecal water genotoxicity in healthy free-living young Italian people. Food Chem. Toxicol. 2014, 64, 104–109. [Google Scholar] [CrossRef]
- Vanhaecke, L.; Knize, M.G.; Noppe, H.; De Brabander, H.; Verstraete, W.; Van de Wiele, T. Intestinal bacteria metabolize the dietary carcinogen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine following consumption of a single cooked chicken meal in humans. Food Chem. Toxicol. 2008, 46, 140–148. [Google Scholar] [CrossRef]
- Kim, J.; Park, S.-H.; Do, K.H.; Kim, D.; Moon, Y. Interference with mutagenic aflatoxin B1-induced checkpoints through antagonistic action of ochratoxin A in intestinal cancer cells: A molecular explanation on potential risk of crosstalk between carcinogens. Oncotarget 2016, 7, 39627–39639. [Google Scholar] [CrossRef]
- Prete, R.; Tofalo, R.; Federici, E.; Ciarrocchi, A.; Cenci, G.; Corsetti, A. Food-associated Lactobacillus plantarum and yeasts inhibit the genotoxic effect of 4-nitroquinoline-1-oxide. Front. Microbiol. 2017, 8, 2349. [Google Scholar] [CrossRef]
- Ribière, C.; Peyret, P.; Parisot, N.; Darcha, C.; Déchelotte, P.J.; Barnich, N.; Peyretaillade, E.; Boucher, D. Oral exposure to environmental pollutant benzo[a]pyrene impacts the intestinal epithelium and induces gut microbial shifts in murine model. Sci. Rep. 2016, 6, 31027. [Google Scholar] [CrossRef]
- Defois, C.; Ratel, J.; Denis, S.; Batut, B.; Beugnot, R.; Peyretaillade, E.; Engel, E.; Peyret, P. Environmental pollutant benzo[a]pyrene impacts the volatile metabolome and transcriptome of the human gut microbiota. Front. Microbiol. 2017, 8, 1562. [Google Scholar] [CrossRef]
- Defois, C.; Ratel, J.; Garrait, G.; Denis, S.; Le Goff, O.; Talvas, J.; Mosoni, P.; Engel, E.; Peyret, P. Food chemicals disrupt human gut microbiota activity and impact intestinal homeostasis as revealed by in vitro systems. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Morotomi, M.; Mutai, M. In vitro binding of potent mutagenic pyrolysates to intestinal bacteria. J. Natl. Cancer Inst. 1986, 77, 195–201. [Google Scholar] [PubMed]
- Orrhage, K.; Sillerström, E.; Gustafsson, J.-Å.; Nord, C.E.; Rafter, J. Binding of mutagenic heterocyclic amines by intestinal and lactic acid bacteria. Mutat. Res. Mol. Mech. Mutagen. 1994, 311, 239–248. [Google Scholar] [CrossRef]
- Vanhaecke, L.; Vercruysse, F.; Boon, N.; Verstraete, W.; Cleenwerck, I.; De Wachter, M.; De Vos, P.; van de Wiele, T. Isolation and characterization of human intestinal bacteria capable of transforming the dietary carcinogen 2-Amino-1-Methyl-6-Phenylimidazo[4,5-b]Pyridine. Appl. Environ. Microbiol. 2008, 74, 1469–1477. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, J.; Empl, M.T.; Schwab, C.; Fekry, M.I.; Engels, C.; Schneider, M.; Lacroix, C.; Steinberg, P.; Sturla, S.J. Gut microbial transformation of the dietary imidazoquinoxaline mutagen MelQx reduces its cytotoxic and mutagenic potency. Toxicol. Sci. 2017, 159, 266–276. [Google Scholar] [CrossRef]
- Carmody, R.N.; Turnbaugh, P.J. Host-microbial interactions in the metabolism of therapeutic and diet-derived xenobiotics. J. Clin. Investig. 2014, 124, 4173–4181. [Google Scholar] [CrossRef] [PubMed]
- Kassie, F. Intestinal microflora plays a crucial role in the genotoxicity of the cooked food mutagen 2-amino-3-methylimidazo[4,5-f]quinoline (IQ). Carcinogenesis 2001, 22, 1721–1725. [Google Scholar] [CrossRef]
- Humblot, C.; Murkovic, M.; Rigottier-Gois, L.; Bensaada, M.; Bouclet, A.; Andrieux, C.; Anba, J.; Rabot, S. Glucuronidase in human intestinal microbiota is necessary for the colonic genotoxicity of the food-borne carcinogen 2-amino-3-methylimidazo[4,5-f]quinoline in rats. Carcinogenesis 2007, 28, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Kaderlik, K.R.; Mulder, G.J.; Turesky, R.J.; Lang, N.P.; Teitel, C.H.; Chiarelli, M.P.; Kadlubar, F.F. Glucuronidation of N -hydroxy heterocyclic amines by human and rat liver microsomes. Carcinogenesis 1994, 15, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Do, K.N.; Fink, L.N.; Jensen, T.E.; Gautier, L.; Parlesak, A. TLR2 Controls intestinal carcinogen detoxication by CYP1A1. PLoS ONE 2012, 7, e32309. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development. Genetic Toxicology Guidance Document; OECD: Paris, France, 2015; pp. 1–58. [Google Scholar]
- Thybaud, V.; Lorge, E.; Levy, D.D.; van Benthem, J.; Douglas, G.R.; Marchetti, F.; Moore, M.M.; Schoeny, R. Main issues addressed in the 2014-2015 revisions to the OECD Genetic Toxicology Test Guidelines. Environ. Mol. Mutagen. 2017, 58, 284–295. [Google Scholar] [CrossRef]
- Turkez, H.; Arslan, M.E.; Ozdemir, O. Genotoxicity testing: Progress and prospects for the next decade. Expert Opin. Drug Metab. Toxicol. 2017, 13, 1089–1098. [Google Scholar] [CrossRef]
- Iglesias, T.; López de Cerain, A.; Irache, J.; Martín-Arbella, N.; Wilcox, M.; Pearson, J.; Azqueta, A. Evaluation of the cytotoxicity, genotoxicity and mucus permeation capacity of several surface modified poly(anhydride) nanoparticles designed for oral drug delivery. Int. J. Pharm. 2017, 517, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Róka, E.; Ujhelyi, Z.; Deli, M.; Bocsik, A.; Fenyvesi, É.; Szente, L.; Fenyvesi, F.; Vecsernyés, M.; Váradi, J.; Fehér, P.; et al. Evaluation of the cytotoxicity of α-cyclodextrin derivatives on the Caco-2 Cell line and human erythrocytes. Molecules 2015, 20, 20269–20285. [Google Scholar] [CrossRef]
- Valdés, L.; Gueimonde, M.; Ruas-Madiedo, P. Monitoring in real time the cytotoxic effect of Clostridium difficile upon the intestinal epithelial cell line HT29. J. Microbiol. Methods 2015, 119, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Fair, K.L.; Colquhoun, J.; Hannan, N.R.F. Intestinal organoids for modelling intestinal development and disease. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170217. [Google Scholar] [CrossRef]
- Kaushik, G.; Ponnusamy, M.P.; Batra, S.K. Concise review: Current status of three-dimensional organoids as preclinical models. Stem Cells 2018, 36, 1329–1340. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Cai, Y.-Z.; Zhang, Y. Natural phenolic compounds from medicinal herbs and dietary plants: Potential use for cancer prevention. Nutr. Cancer 2009, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Anderson, D.; de Pascual-Teresa, S.; Santos-Buelga, C.; Clifford, M.N.; Ioannides, C. Evaluation of the antigenotoxic potential of monomeric and dimeric flavanols, and black tea polyphenols against heterocyclic amine-induced DNA damage in human lymphocytes using the Comet assay. Mutat. Res. 2002, 515, 39–56. [Google Scholar] [CrossRef]
- Quelhas, I.; Petisca, C.; Viegas, O.; Melo, A.; Pinho, O.; Ferreira, I.M.P.L.V.O. Effect of green tea marinades on the formation of heterocyclic aromatic amines and sensory quality of pan-fried beef. Food Chem. 2010, 122, 98–104. [Google Scholar] [CrossRef]
- Bear, W.L.; Teel, R.W. Effects of citrus flavonoids on the mutagenicity of heterocyclic amines and on cytochrome P450 1A2 activity. Anticancer Res. 2000, 20, 3609–3614. [Google Scholar] [PubMed]
- Edenharder, R.; Sager, J.W.; Glatt, H.; Muckel, E.; Platt, K.L. Protection by beverages, fruits, vegetables, herbs, and flavonoids against genotoxicity of 2-acetylaminofluorene and 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) in metabolically competent V79 cells. Mutat. Res. 2002, 521, 57–72. [Google Scholar] [CrossRef]
- Cheng, K.-W.; Wu, Q.; Zheng, Z.P.; Peng, X.; Simon, J.E.; Chen, F.; Wang, M. Inhibitory effect of fruit extracts on the formation of heterocyclic amines. J. Agric. Food Chem. 2007, 55, 10359–10365. [Google Scholar] [CrossRef] [PubMed]
- Sepahpour, S.; Selamat, J.; Khatib, A.; Manap, M.Y.A.; Abdull Razis, A.F.; Hajeb, P. Inhibitory effect of mixture herbs/spices on formation of heterocyclic amines and mutagenic activity of grilled beef. Food Addit. Contam. Part A 2018, 35, 1911–1927. [Google Scholar] [CrossRef] [PubMed]
- Balogh, Z.; Gray, J.I.; Gomaa, E.A.; Booren, A.M. Formation and inhibition of heterocyclic aromatic amines in fried ground beef patties. Food Chem. Toxicol. 2000, 38, 395–401. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I. Effect of Bifidobacterium longum and inulin on gut bacterial metabolism and carcinogen-induced aberrant crypt foci in rats. Carcinogenesis 1998, 19, 281–285. [Google Scholar] [CrossRef]
- Chen, C.-C.; Lin, W.-C.; Kong, M.-S.; Shi, H.N.; Walker, W.A.; Lin, C.-Y.; Huang, C.-T.; Lin, Y.-C.; Jung, S.-M.; Lin, T.-Y. Oral inoculation of probiotics Lactobacillus acidophilus NCFM suppresses tumour growth both in segmental orthotopic colon cancer and extra-intestinal tissue. Br. J. Nutr. 2012, 107, 1623–1634. [Google Scholar] [CrossRef]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Hago, A.M.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; et al. Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef]
- Lili, Z.; Junyan, W.; Hongfei, Z.; Baoqing, Z.; Bolin, Z. Detoxification of cancerogenic compounds by lactic acid bacteria strains. Crit. Rev. Food Sci. Nutr. 2017, 1–16. [Google Scholar] [CrossRef]
- Tsuda, H.; Hara, K.; Miyamoto, T. Binding of mutagens to exopolysaccharide produced by Lactobacillus plantarum mutant strain 301102S. J. Dairy Sci. 2008, 91, 2960–2966. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, A.; Nagpal, R.; Mohania, D.; Behare, P.; Verma, V.; Kumar, P.; Poddar, D.; Aggarwal, P.K.; Henry, C.J.K.; et al. Cancer-preventing attributes of probiotics: An update. Int. J. Food Sci. Nutr. 2010, 61, 473–496. [Google Scholar] [CrossRef]
- Nowak, A.; Libudzisz, Z. Ability of probiotic Lactobacillus casei DN 114001 to bind or/and metabolise heterocyclic aromatic amines in vitro. Eur. J. Nutr. 2009, 48, 419–427. [Google Scholar] [CrossRef]
- Nowak, A.; Śliżewska, K.; Otlewska, A. Antigenotoxic activity of lactic acid bacteria, prebiotics, and products of their fermentation against selected mutagens. Regul. Toxicol. Pharmacol. 2015, 73, 938–946. [Google Scholar] [CrossRef]
- Beer, F.; Urbat, F.; Steck, J.; Huch, M.; Bunzel, D.; Bunzel, M.; Kulling, S.E. Metabolism of foodborne heterocyclic aromatic amines by Lactobacillus reuteri DSM 20016. J. Agric. Food Chem. 2017, 65, 6797–6811. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Zhang, X.; Covasa, M. Emerging roles of lactic acid bacteria in protection against colorectal cancer. World J. Gastroenterol. 2014, 20, 7878–7886. [Google Scholar] [CrossRef]
- Nowak, A.; Śliżewska, K.; Błasiak, J.; Libudzisz, Z. The influence of Lactobacillus casei DN 114 001 on the activity of faecal enzymes and genotoxicity of faecal water in the presence of heterocyclic aromatic amines. Anaerobe 2014, 30, 129–136. [Google Scholar] [CrossRef]
- Gianfredi, V.; Salvatori, T.; Villarini, M.; Moretti, M.; Nucci, D.; Realdon, S. Is dietary fibre truly protective against colon cancer? A systematic review and meta-analysis. Int. J. Food Sci. Nutr. 2018, 69, 904–915. [Google Scholar] [CrossRef]
- Lim, C.C.; Ferguson, L.R.; Tannock, G.W. Dietary fibres as “prebiotics”: Implications for colorectal cancer. Mol. Nutr. Food Res. 2005, 49, 609–619. [Google Scholar] [CrossRef]
- Mahowald, M.A.; Rey, F.E.; Seedorf, H.; Turnbaugh, P.J.; Fulton, R.S.; Wollam, A.; Shah, N.; Wang, C.; Magrini, V.; Wilson, R.K.; et al. Characterizing a model human gut microbiota composed of members of its two dominant bacterial phyla. Proc. Natl. Acad. Sci. USA 2009, 106, 5859–5864. [Google Scholar] [CrossRef]
- Jakszyn, P.; Agudo, A.; Ibáñez, R.; García-Closas, R.; Pera, G.; Amiano, P.; González, C.A. Development of a food database of nitrosamines, heterocyclic amines, and polycyclic aromatic hydrocarbons. J. Nutr. 2004, 134, 2011–2014. [Google Scholar] [CrossRef]


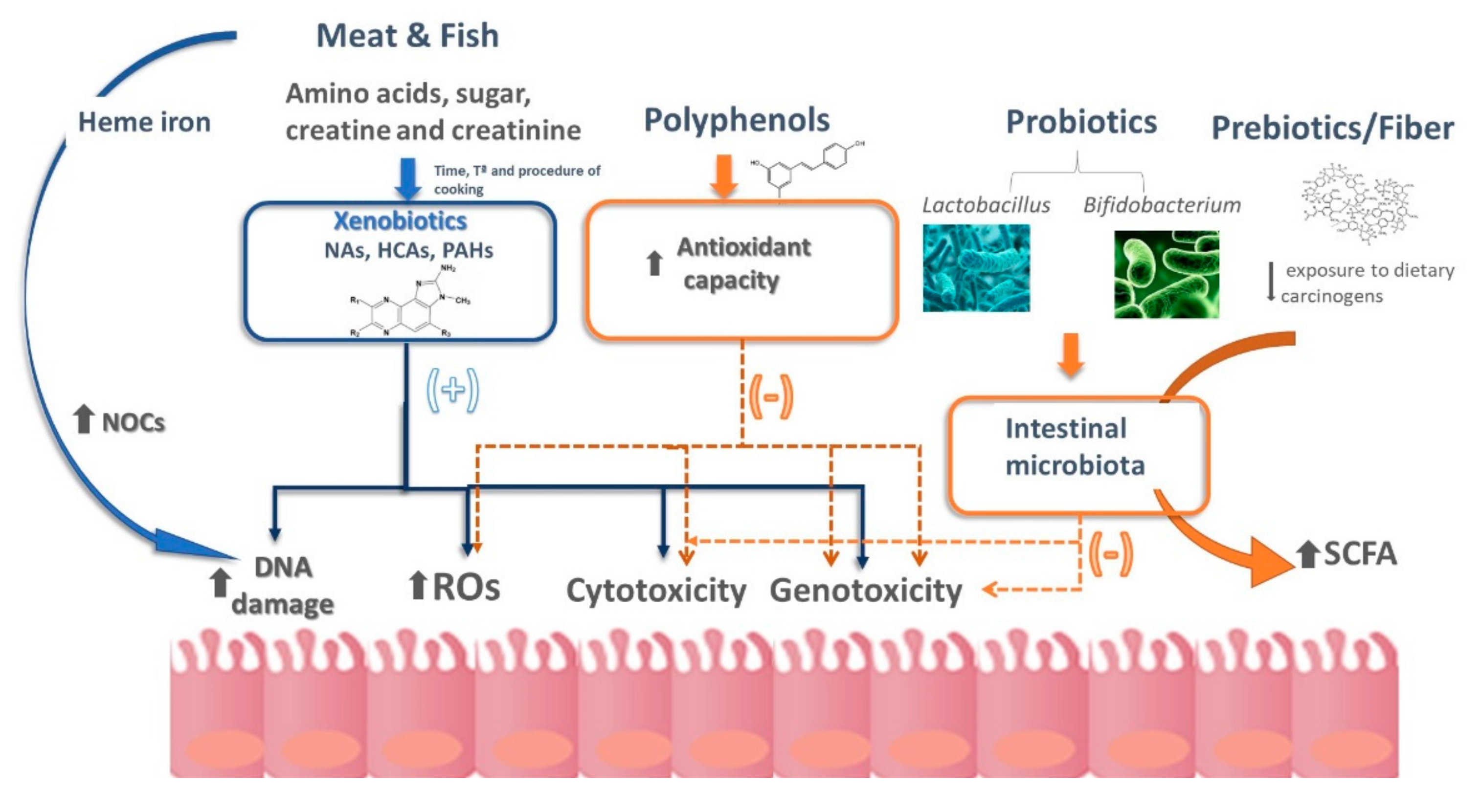{kind=link}
| Year | No. Subjects | Analytical Category | Source | Dose | Pathology | Ref. |
|---|---|---|---|---|---|---|
| 2018 | 407,270 | HCAs | Red meat | n.a a | MeIQx and DiMeIQx association with all anatomical subsites of colorectal cancer. PhIP associations with total colorectal and colon cancers. Not evidenced an association between ingested B(a)P and CRC | [46] |
| MeIQx | ||||||
| DiMeIQx | ||||||
| PhIP | ||||||
| PAHs | ||||||
| B(a)P | ||||||
| 2018 | 76,657 | HCAs | Red meat | 50 ng/day | Association of HCAs, B(a)P, and mutagenicity index with the risk of colorectal adenomas | [47] |
| MeIQx | n.a a | |||||
| DiMeIQx | 40 ng/day | |||||
| PhIP | n.a a | |||||
| PAHs | ||||||
| B(a)P | ||||||
| 2013 | total 3707: 1062 cases and 1645 controls | HCAs | Red meat | n.a a | Colon cancer | [48] |
| MeIQx | ||||||
| DiMeIQ | ||||||
| PhIP |
| Main Mechanism | Molecules/Compounds Involved | Microbial Group | Experimental Approach Used for Study | Mode of Action | Ref. | |
|---|---|---|---|---|---|---|
| Direct mechanisms | Genotoxins | Typhoid toxin | Salmonella enterica serovar Typhi | In vitro and animal models | DNAse activity; induction of symptoms characteristic of typhoid fever | [64] |
| Cytolethal distending toxin | Proteobacteria | Cell lines and primary cell and mouse models of chronic infections | DNase activity; Proinflamation and carcinogenic potential | [65,66,67] | ||
| Colibactin | Escherichia coli group B | Eukaryotic cells | DNA double-strand breaks | [68] | ||
| Epidemiological and animal model | DNA double-strand breaks in vitro and in vivo; enhanced tumour growth by senescence | [69,70] | ||||
| Alteration of host cellular cycle | Cytotoxin-associated gene A Vacuolating cytotoxin A | Helicobacter pylori | Molecular, experimental and epidemiological | DNA damage; Increases IL-8; produces reactive oxygen species (ROS) and nitric oxide; increases concentrations of cyclo-oxygenase 2; decreases apoptosis; and increases cell proliferation | [66,71] | |
| Enterotoxin | Bacteroides fragilis | In vitro and epidemiological | DNA damage; high levels of ROS; Diarrheal disease, associated with colorectal cancer | [62,72] | ||
| Adhesin A | Fusobacterium nucleatum | In vitro and epidemiological | Activation of β catenin pathway | [66,73] | ||
| ExoS exotoxin | Pseudomonas aeruginosa | In vitro, experimental and epidemiological | Activation of pathways with final mechanism leading to DNA damage; unknown mechanisms in cancer generation | [62,66] | ||
| Cysteine protease-like | Shigella flexneri | In vitro and epidemiological | Potassium outflow conducting to ROS production; induce degradation of p53; DNA damage; dysentery | [62,66] | ||
| Avirulence protein A | Salmonella enterica | In vitro and mouse model of inflammation-associated cancer | Target β-catenin pathway; colonic tumorigenesis and tumour progression | [66] | ||
| Cytotoxic necrotising factor | Escherichia coli | In vitro and animal models | Activates Rho GTPase; modifies cytoskeleton; triggers G1-S transition; downregulate mismatch repair genes; the role of CNF in infections in not clear | [71,74] | ||
| Cycle-inhibiting factor | In vitro | Inhibition of mitosis | [75] | |||
| Secondary bile acids | Anaerobic bacteria with 7-α dehydroxylation activity of primary bile acids | In vitro colon cells and animal models | Changes in physicochemical membrane properties; Apoptosis and genomic damage by ROS; Deoxycholic acid is carcinogenic at high doses and long-term treatment in animal models | [76] | ||
| Indirect mechanisms | Oxidative stress | Reactive oxygen species | Peptostreptococcus anaerobius | In vivo, in vitro and epidemiological | Increase of human colon tumour tissues and adenomas; these bacteria increase colon dysplasia in a mouse model of CRC by induction of ROS levels, which promotes cholesterol synthesis and cell proliferation. | [77] |
| Enterococcus faecalis | In vitro and in vivo models, epidemiological | Induction of ROS, activation of macrophages; promotion of tumorigenesis | [66,78] | |||
| Faecal matrix | In vitro | Unknown reducing agent | [79] | |||
| Formation of H2S | H2S | Sulfate-reducing bacteria | Epidemiological and in vitro models | Promotes instability or cumulative mutations in a predisposed genetic background | [80] | |
| Inflammation | Wall-extracted antigen | Streptococcus bovis | Epidemiological and molecular | Activation of cyclo-oxygenase 2, interleukin 8 production, and cell proliferation | [71] | |
| Disabling cellular DNA repair process | Listeriolysin O | Listeria monocytogenes | In vitro and epidemiological | Pore formation in intestinal host cells; Prevention of recruitment of repair complex to DNA breaks; listeriosis | [66] | |
| Secreted effector protein EspF | Escherichia coli | In vitro | Down-regulation DNA mismatch repair | [66] | ||
| Protein metabolism | Phenol/indol/p-cresol/ | Intestinal bacteria | Colonic cells | Increased anion superoxide production and genotoxic effects | [81,82] | |
| Fecapentanes | Bacteroides sp. | In vitro; In vivo | Cytotoxic and mutagenic effects via ROS production; Controversial in vivo effect | [63,83] | ||
| Ammonium | Intestinal bacteria | In vitro | Antiproliferative effect without decrease of cell viability | [84] | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogacka, A.M.; Gómez-Martín, M.; Suárez, A.; González-Bernardo, O.; de los Reyes-Gavilán, C.G.; González, S. Xenobiotics Formed during Food Processing: Their Relation with the Intestinal Microbiota and Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2051. https://doi.org/10.3390/ijms20082051
Nogacka AM, Gómez-Martín M, Suárez A, González-Bernardo O, de los Reyes-Gavilán CG, González S. Xenobiotics Formed during Food Processing: Their Relation with the Intestinal Microbiota and Colorectal Cancer. International Journal of Molecular Sciences. 2019; 20(8):2051. https://doi.org/10.3390/ijms20082051
Chicago/Turabian StyleNogacka, Alicja M., María Gómez-Martín, Adolfo Suárez, Oscar González-Bernardo, Clara G. de los Reyes-Gavilán, and Sonia González. 2019. "Xenobiotics Formed during Food Processing: Their Relation with the Intestinal Microbiota and Colorectal Cancer" International Journal of Molecular Sciences 20, no. 8: 2051. https://doi.org/10.3390/ijms20082051
APA StyleNogacka, A. M., Gómez-Martín, M., Suárez, A., González-Bernardo, O., de los Reyes-Gavilán, C. G., & González, S. (2019). Xenobiotics Formed during Food Processing: Their Relation with the Intestinal Microbiota and Colorectal Cancer. International Journal of Molecular Sciences, 20(8), 2051. https://doi.org/10.3390/ijms20082051