High Body Mass Index is Associated with Elevated Blood Levels of Progerin mRNA


Abstract
1. Introduction
2. Results
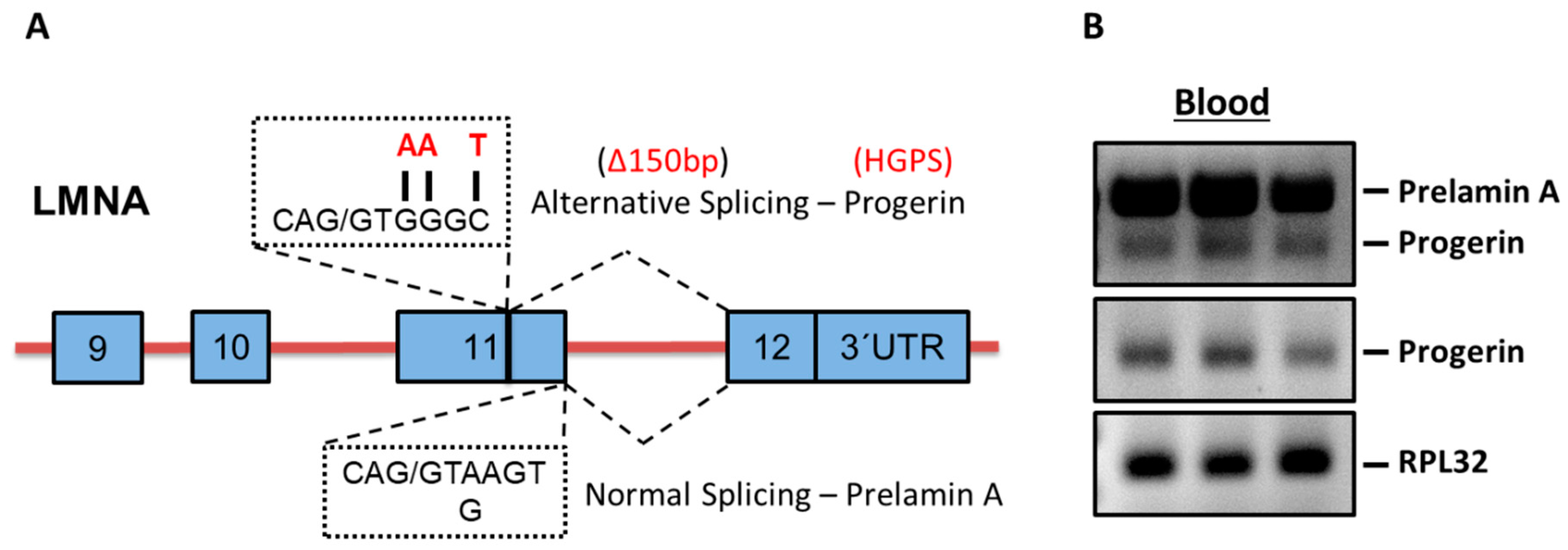
2.1. Progerin mRNA is Detectable in Human Blood Samples
2.2. Characteristics of Study Groups
2.3. Progerin mRNA and CRP Were Upregulated in the Overweight Group
2.4. BMI Correlates Positively with Progerin mRNA, CRP, and HbA1c
3. Discussion
Limitations
4. Materials and Methods
4.1. Study Population
4.2. RT-PCR of mRNA
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | Body-Mass-Index |
| CVS | Cardiovascular system |
| DCMP | Dilative cardiomyopathy |
| HGPS | Hutchinson–Gilford-Progeria syndrome |
| Hs-CRP | High-Sensitive C-Reactive Protein |
| HCMP | Hypertrophic cardiomyopathy |
| ICMP | Ischemic cardiomyopathy |
| NYHA | New York Heart Association |
| PCR | Polymerase Chain Reaction |
Appendix


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DCMP | ICMP | HCMP | Others | Total | |
|---|---|---|---|---|---|
| BMI < 25 kg/m2, n (%) | 22 (49) | 13 (28) | 5 (11) | 6 (13) | 46 |
| BMI ≥ 25 kg/m2, n (%) | 37 (57) | 18 (28) | 7 (11) | 3 (5) | 65 |
| Total | 59 | 31 | 12 | 9 | 111 |
References
- Yosipovitch, G.; DeVore, A.; Dawn, A. Obesity and the skin: Skin physiology and skin manifestations of obesity. J. Am. Acad. Dermatol. 2007, 56, 901–916, quiz 917–920. [Google Scholar] [CrossRef] [PubMed]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Sacks, G.; Swinburn, B.; Lawrence, M. Obesity Policy Action framework and analysis grids for a comprehensive policy approach to reducing obesity. Obes. Rev. 2009, 10, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef]
- Berrington de Gonzalez, A.; Hartge, P.; Cerhan, J.R.; Flint, A.J.; Hannan, L.; MacInnis, R.J.; Moore, S.C.; Tobias, G.S.; Anton-Culver, H.; Freeman, L.B.; et al. Body-mass index and mortality among 1.46 million white adults. N. Engl. J. Med. 2010, 363, 2211–2219. [Google Scholar] [CrossRef]
- Miard, S.; Picard, F. Obesity and aging have divergent genomic fingerprints. Int. J. Obes. (Lond.) 2008, 32, 1873–1874. [Google Scholar] [CrossRef] [PubMed]
- Barzilai, N.; Huffman, D.M.; Muzumdar, R.H.; Bartke, A. The critical role of metabolic pathways in aging. Diabetes 2012, 61, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Slawik, M.; Vidal-Puig, A.J. Lipotoxicity, overnutrition and energy metabolism in aging. Ageing Res. Rev. 2006, 5, 144–164. [Google Scholar] [CrossRef]
- Vidak, S.; Foisner, R. Molecular insights into the premature aging disease progeria. Histochem. Cell Biol. 2016, 145, 401–417. [Google Scholar] [CrossRef]
- Niccoli, T.; Partridge, L. Ageing as a risk factor for disease. Curr. Biol. 2012, 22, R741–R752. [Google Scholar] [CrossRef] [PubMed]
- De Sandre-Giovannoli, A.; Bernard, R.; Cau, P.; Navarro, C.; Amiel, J.; Boccaccio, I.; Lyonnet, S.; Stewart, C.L.; Munnich, A.; Le Merrer, M.; et al. Lamin a truncation in Hutchinson-Gilford progeria. Science 2003, 300, 2055. [Google Scholar] [CrossRef]
- Ułas, J.; Gradkowska, M.G.; Jezierska, M.; Skup, M.G.; Skangiel-Kramska, J.; Odefeld-Nowak, B. Bilateral changes in glutamate uptake, muscarinic receptor binding and acetylcholinesterase level in the rat hippocampus after unilateral entorhinal cortex lesions. Neurochem. Int. 1986, 9, 255–263. [Google Scholar]
- Shumaker, D.K.; Kuczmarski, E.R.; Goldman, R.D. The nucleoskeleton: Lamins and actin are major players in essential nuclear functions. Curr. Opin. Cell Biol. 2003, 15, 358–366. [Google Scholar] [CrossRef]
- Amendola, M.; van Steensel, B. Mechanisms and dynamics of nuclear lamina-genome interactions. Curr. Opin. Cell Biol 2014, 28, 61–68. [Google Scholar] [CrossRef]
- Ahmed, M.S.; Ikram, S.; Bibi, N.; Mir, A. Hutchinson-Gilford Progeria Syndrome: A Premature Aging Disease. Mol. Neurobiol. 2018, 55, 4417–4427. [Google Scholar] [CrossRef]
- Olive, M.; Harten, I.; Mitchell, R.; Beers, J.K.; Djabali, K.; Cao, K.; Erdos, M.R.; Blair, C.; Funke, B.; Smoot, L.; et al. Cardiovascular pathology in Hutchinson-Gilford progeria: Correlation with the vascular pathology of aging. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2301–2309. [Google Scholar] [CrossRef]
- Hilton, B.A.; Liu, J.; Cartwright, B.M.; Liu, Y.; Breitman, M.; Wang, Y.; Jones, R.; Tang, H.; Rusinol, A.; Musich, P.R.; et al. Progerin sequestration of PCNA promotes replication fork collapse and mislocalization of XPA in laminopathy-related progeroid syndromes. FASEB J. 2017. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T. Lamin A-dependent nuclear defects in human aging. Science 2006, 312, 1059–1063. [Google Scholar] [CrossRef]
- Rodriguez, S.; Coppede, F.; Sagelius, H.; Eriksson, M. Increased expression of the Hutchinson-Gilford progeria syndrome truncated lamin A transcript during cell aging. Eur. J. Hum. Genet. 2009, 17, 928–937. [Google Scholar] [CrossRef]
- Deschenes, M.; Chabot, B. The emerging role of alternative splicing in senescence and aging. Aging Cell 2017, 16, 918–933. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; Garcia-Cardena, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Hamczyk, M.R.; Villa-Bellosta, R.; Quesada, V.; Gonzalo, P.; Vidak, S.; Nevado, R.M.; Andres-Manzano, M.J.; Misteli, T.; Lopez-Otin, C.; Andres, V. Progerin accelerates atherosclerosis by inducing endoplasmic reticulum stress in vascular smooth muscle cells. EMBO Mol. Med. 2019. [Google Scholar] [CrossRef]
- Gielen, M.; Hageman, G.J.; Antoniou, E.E.; Nordfjall, K.; Mangino, M.; Balasubramanyam, M.; de Meyer, T.; Hendricks, A.E.; Giltay, E.J.; Hunt, S.C.; et al. Body mass index is negatively associated with telomere length: A collaborative cross-sectional meta-analysis of 87 observational studies. Am. J. Clin. Nutr. 2018, 108, 453–475. [Google Scholar] [CrossRef]
- Osborne, T.B.; Mendel, L.B.; Ferry, E.L. The Effect of Retardation of Growth Upon the Breeding Period and Duration of Life of Rats. Science 1917, 45, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Progeria, rapamycin and normal aging: Recent breakthrough. Aging (Albany NY) 2011, 3, 685–691. [Google Scholar] [CrossRef]
- Messner, M.; Ghadge, S.K.; Goetsch, V.; Wimmer, A.; Dorler, J.; Polzl, G.; Zaruba, M.M. Upregulation of the aging related LMNA splice variant progerin in dilated cardiomyopathy. PLoS ONE 2018, 13, e0196739. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.A.; Grochova, D.; McKenna, T.; Borate, B.; Trivedi, N.S.; Erdos, M.R.; Eriksson, M. Global genome splicing analysis reveals an increased number of alternatively spliced genes with aging. Aging Cell 2016, 15, 267–278. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Blomberg, B.B. Ageing and obesity similarly impair antibody responses. Clin. Exp. Immunol. 2017, 187, 64–70. [Google Scholar] [CrossRef]
- Abdelsamed, H.A.; Zebley, C.C.; Youngblood, B. Epigenetic Maintenance of Acquired Gene Expression Programs during Memory CD8 T Cell Homeostasis. Front Immunol. 2018, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Franceschi, C.; Monti, D.; Ginaldi, L. Inflamm-ageing and lifelong antigenic load as major determinants of ageing rate and longevity. FEBS Lett. 2005, 579, 2035–2039. [Google Scholar] [CrossRef]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Deleidi, M.; Jaggle, M.; Rubino, G. Immune aging, dysmetabolism, and inflammation in neurological diseases. Front Neurosci. 2015, 9, 172. [Google Scholar] [CrossRef]
- Spaulding, C.C.; Walford, R.L.; Effros, R.B. Calorie restriction inhibits the age-related dysregulation of the cytokines TNF-alpha and IL-6 in C3B10RF1 mice. Mech Ageing Dev. 1997, 93, 87–94. [Google Scholar] [CrossRef]
- Starr, M.E.; Evers, B.M.; Saito, H. Age-associated increase in cytokine production during systemic inflammation: Adipose tissue as a major source of IL-6. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 723–730. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Pedersen, M.; Pedersen, B.K. Aging and proinflammatory cytokines. Curr. Opin. Hematol. 2001, 8, 131–136. [Google Scholar] [CrossRef]
- Revechon, G.; Viceconte, N.; McKenna, T.; Sola Carvajal, A.; Vrtacnik, P.; Stenvinkel, P.; Lundgren, T.; Hultenby, K.; Franco, I.; Eriksson, M. Rare progerin-expressing preadipocytes and adipocytes contribute to tissue depletion over time. Sci. Rep. 2017, 7, 4405. [Google Scholar] [CrossRef]
- Pacheco, L.M.; Gomez, L.A.; Dias, J.; Ziebarth, N.M.; Howard, G.A.; Schiller, P.C. Progerin expression disrupts critical adult stem cell functions involved in tissue repair. Aging (Albany NY) 2014, 6, 1049–1063. [Google Scholar] [CrossRef]
- Expert Panel on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults. Executive summary of the clinical guidelines on the identification, evaluation, and treatment of overweight and obesity in adults. Arch. Intern. Med. 1998, 158, 1855–1867. [Google Scholar] [CrossRef]
- Website of the World Health Organization: Health Topics: Obesity. Available online: https://www.who.int/topics/obesity/en/ (accessed on 6 April 2019).
- McMurray, J.J.; Adamopoulos, S.; Anker, S.D.; Auricchio, A.; Bohm, M.; Dickstein, K.; Falk, V.; Filippatos, G.; Fonseca, C.; Gomez-Sanchez, M.A.; et al. ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: The Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure 2012 of the European Society of Cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2012, 33, 1787–1847. [Google Scholar] [PubMed]



| Characteristic | Total Population n = 111 | BMI < 25 (n = 46) | BMI ≥ 25 (n = 65) | P Value | |
|---|---|---|---|---|---|
| Age, years | 56 ± 15 | 54 ± 17 | 58 ± 13 | 0.1 | |
| Female sex, n (%) | 32 (29) | 18 (39) | 14 (22) | 0.14 | |
| Body-mass-index, kg/m2 | 26.6 ± 4.8 | 22.4 ± 1.8 | 29.6 ± 4.0 | 0.000 | |
| NYHA class, n (%) | n = 108 | 2.0 ± 0.7 | 1.8 ± 0.6 | 2.1 ± 0.7 | 0.015 |
| I | 29 (27) | 15 (33) | 14 (22) | ||
| II | 55 (51) | 25 (54) | 30 (46) | ||
| III | 24 (22) | 4 (9) | 20 (31) | ||
| IV | 0 (0) | 0 (0) | 0 (0) | ||
| Medical history, n (%) | |||||
| Smoking, n (%) | n = 108 | 49 | 17 (37) | 32 (49) | 0.17 |
| Diabetes, n (%) | 19 (17) | 5 (11) | 14 (22) | 0.2 | |
| Atrial fibrillation, n (%) | 42 (38) | 10 (22) | 32 (49) | 0.005 | |
| LVEF, n (%) | 38.2 ± 14.9 | 39.9 ± 14.2 | 37.0 ± 15.4 | 0.29 | |
| ACE-inhibitor, n (%) | 55 (50) | 24 (48) | 31 (48) | 0.7 | |
| ARB, n (%) | 33 (30) | 10 (22) | 23 (36) | 0.14 | |
| Beta-blocker | 90 (81) | 34 (74) | 56 (86) | 0.14 | |
| Clinical features | |||||
| NT-proBNP, ng/L | n = 108 | 2094 ± 4421 | 1748 ± 2118 | 2341 ± 5514 | 0.44 |
| TroponinT, ng/L | n = 109 | 28 ± 53 | 32 ± 72 | 24 ± 33 | 0.48 |
| Serum creatinine, mg/dL | 1.2 ± 0.46 | 1.19 ± 0.54 | 1.21 ± 0.40 | 0.77 | |
| GFR, mL/min | 54 ± 10 | 54 ± 10 | 54 ± 10 | 0.55 | |
| Urea, mg/dL | 50 ± 37 | 48 ± 46 | 51 ± 30 | 0.73 | |
| HbA1c, % | n = 110 | 5.9 ± 1.0 | 5.7 ± 1.1 | 6.1 ± 1.1 | 0.017 |
| Triglycerides, mg/dL | 135 ± 68 | 119 ± 59 | 145 ± 73 | 0.041 | |
| Total cholesterol, mg/dL | 172 ± 44 | 181 ± 43 | 166± 45 | 0.08 | |
| LDL, mg/dL | n = 109 | 107 ± 37 | 110 ± 38 | 105 ± 39 | 0.47 |
| HDL, mg/dL | n = 109 | 52 ± 19 | 59.0 ± 22.7 | 46.5 ± 14.0 | 0.003 |
| Hs-CRP, mg/dL | n = 110 | 0.64 ± 1.07 | 0.28 ± 0.24 | 0.9 ± 1.33 | 0.01 |
| GGT, U/I | 86 ± 146 | 87 ± 184 | 86 ± 114 | 0.96 | |
| GOT, U/I | 29.2 ± 12.4 | 28.5 ±12.0 | 29.6 ± 12.7 | 0.65 | |
| Potassium | 4.2 ± 0.5 | 4.3 ± 0.4 | 4.2 ± 0.5 | 0.12 | |
| Sodium | 138 ± 13 | 139 ± 4 | 137 ± 17 | 0.19 | |
| Leucocyte count, G/l | 7.7 ± 2.2 | 7.5 ± 1.9 | 7.9 ± 2.3 | 0.37 | |
| Thrombocytes, G/I | 217 ± 71 | 223 ± 89 | 212 ± 54 | 0.44 | |
| Hemoglobin, G/I | 140 ± 15 | 138 | 136 | 0.26 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messner, M.; Ghadge, S.K.; Schuetz, T.; Seiringer, H.; Pölzl, G.; Zaruba, M.-M. High Body Mass Index is Associated with Elevated Blood Levels of Progerin mRNA. Int. J. Mol. Sci. 2019, 20, 1976. https://doi.org/10.3390/ijms20081976
Messner M, Ghadge SK, Schuetz T, Seiringer H, Pölzl G, Zaruba M-M. High Body Mass Index is Associated with Elevated Blood Levels of Progerin mRNA. International Journal of Molecular Sciences. 2019; 20(8):1976. https://doi.org/10.3390/ijms20081976
Chicago/Turabian StyleMessner, Moritz, Santhosh Kumar Ghadge, Thomas Schuetz, Herbert Seiringer, Gerhard Pölzl, and Marc-Michael Zaruba. 2019. "High Body Mass Index is Associated with Elevated Blood Levels of Progerin mRNA" International Journal of Molecular Sciences 20, no. 8: 1976. https://doi.org/10.3390/ijms20081976
APA StyleMessner, M., Ghadge, S. K., Schuetz, T., Seiringer, H., Pölzl, G., & Zaruba, M.-M. (2019). High Body Mass Index is Associated with Elevated Blood Levels of Progerin mRNA. International Journal of Molecular Sciences, 20(8), 1976. https://doi.org/10.3390/ijms20081976