The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy


Abstract
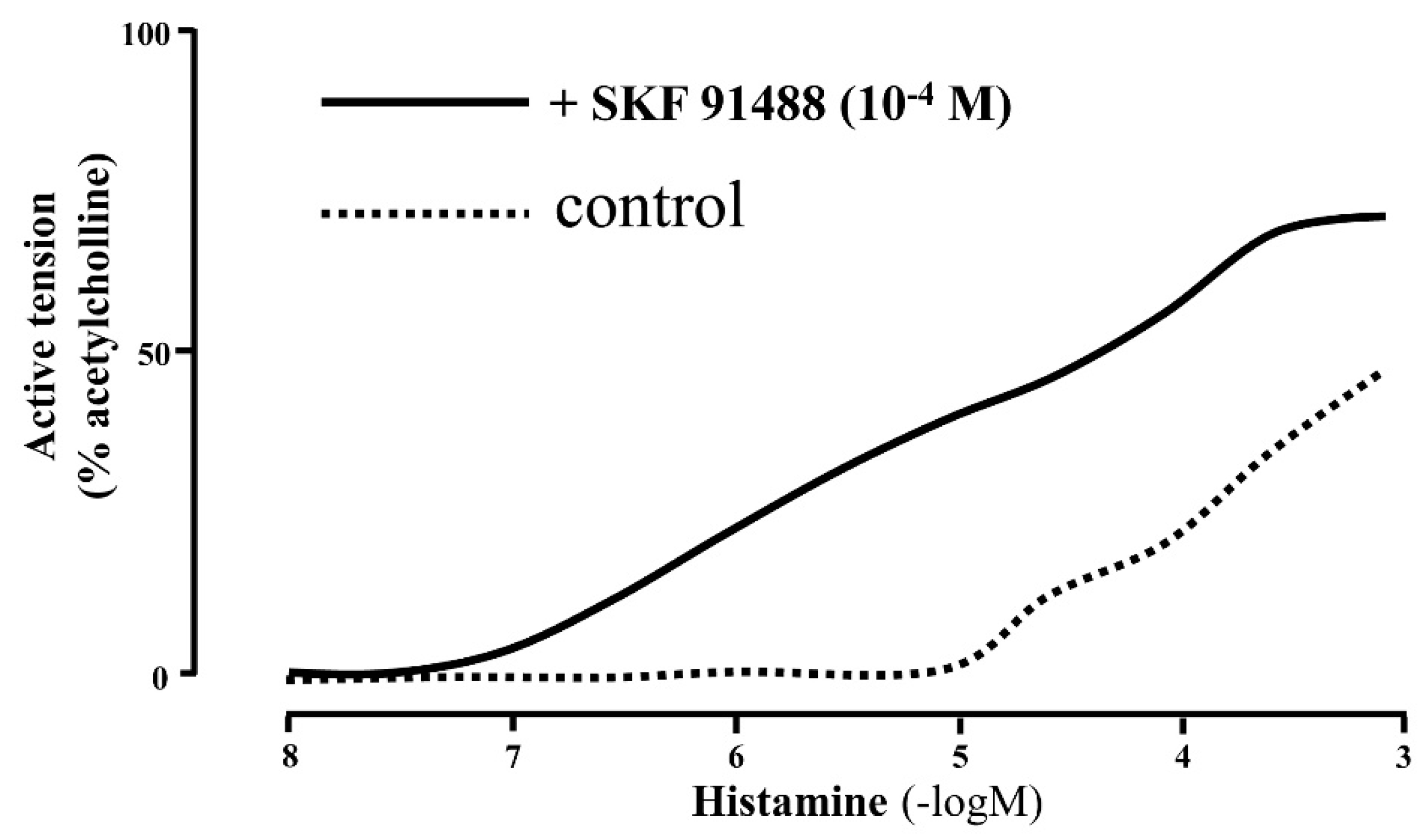
1. Histamine Receptors in the Lung
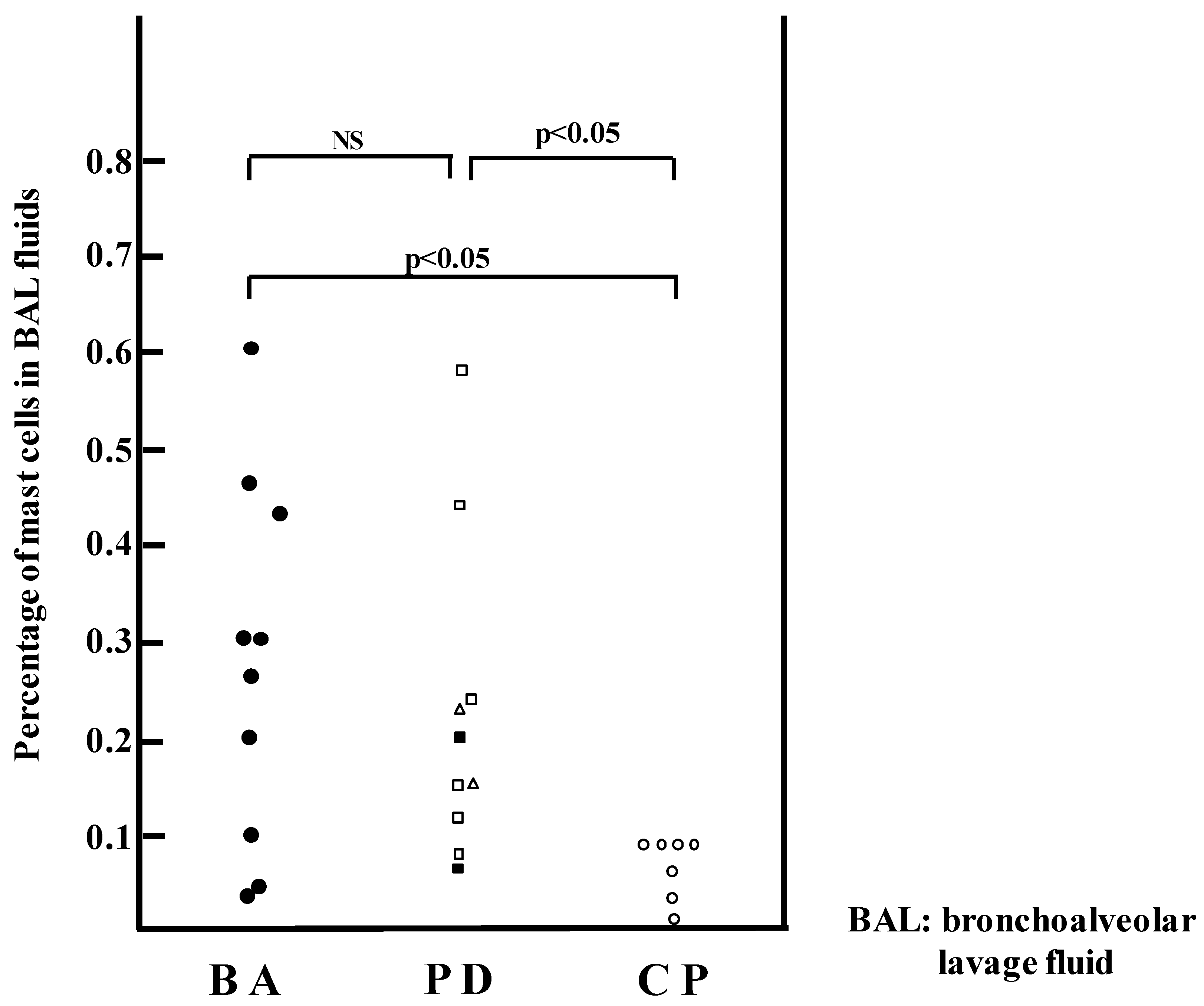
2. Histamine and Mast Cells in Asthma
3. Histamine in the Pathophysiology of Asthma
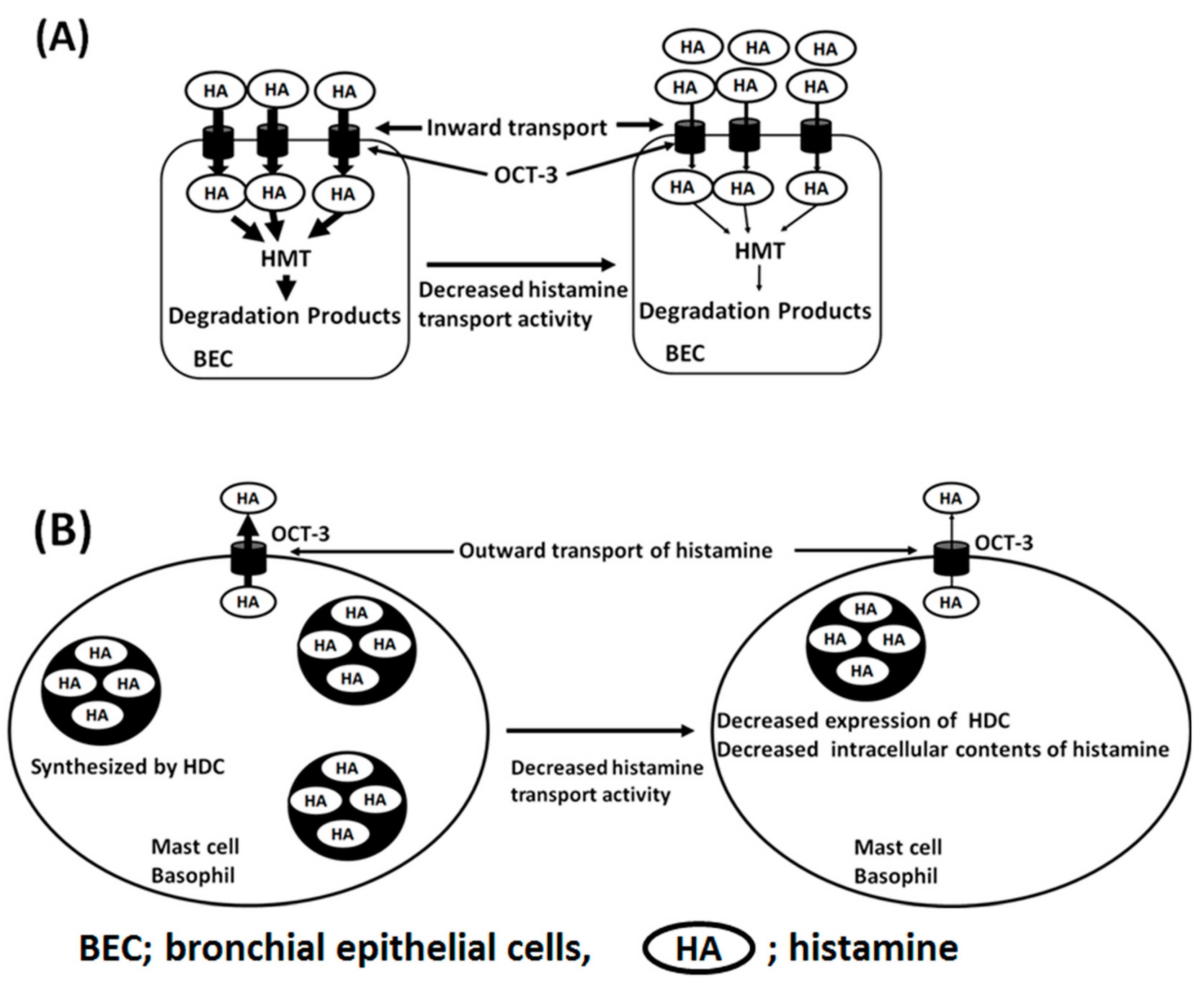
4. Histamine Transport in the Pathophysiology of Asthma
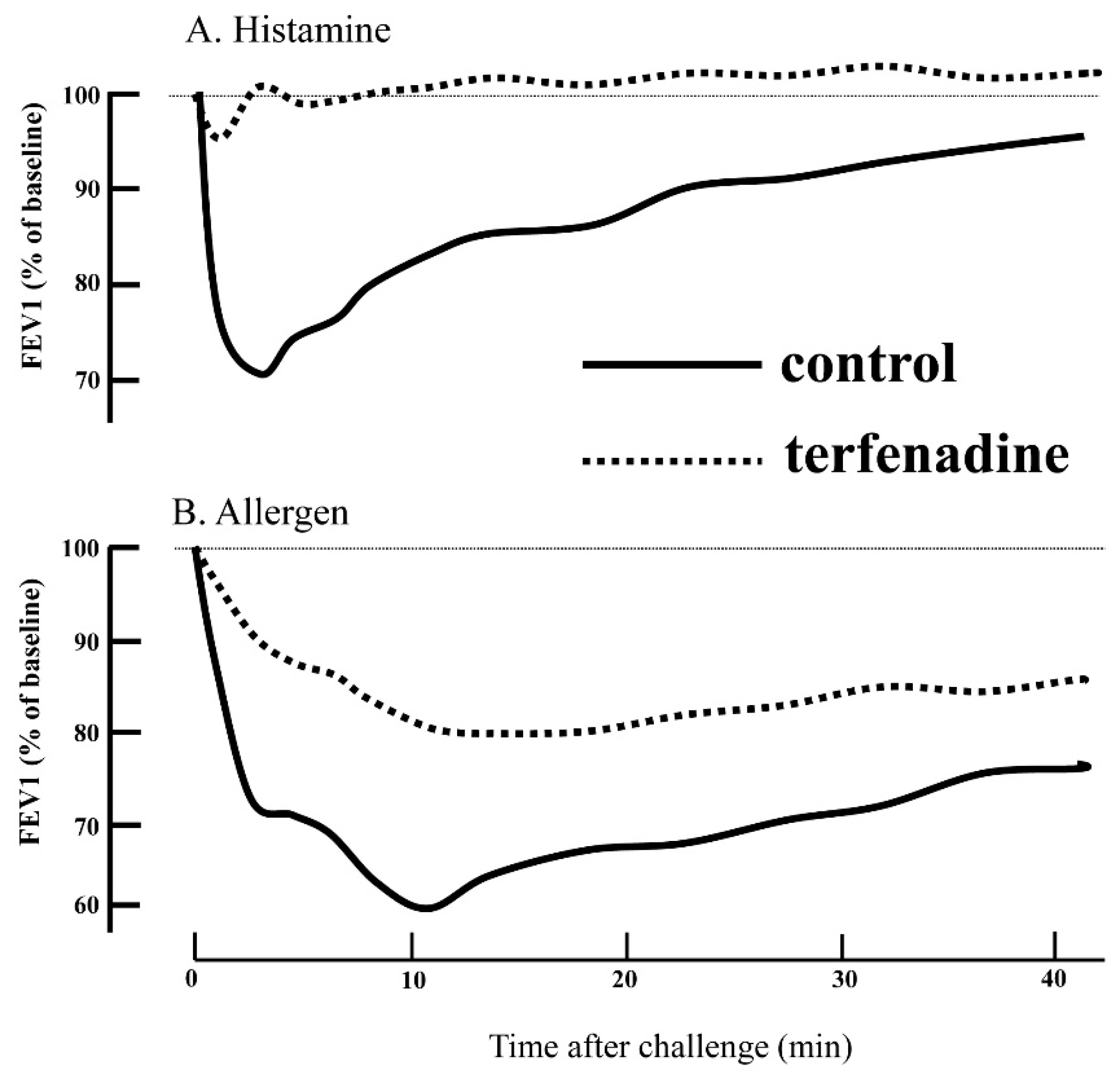
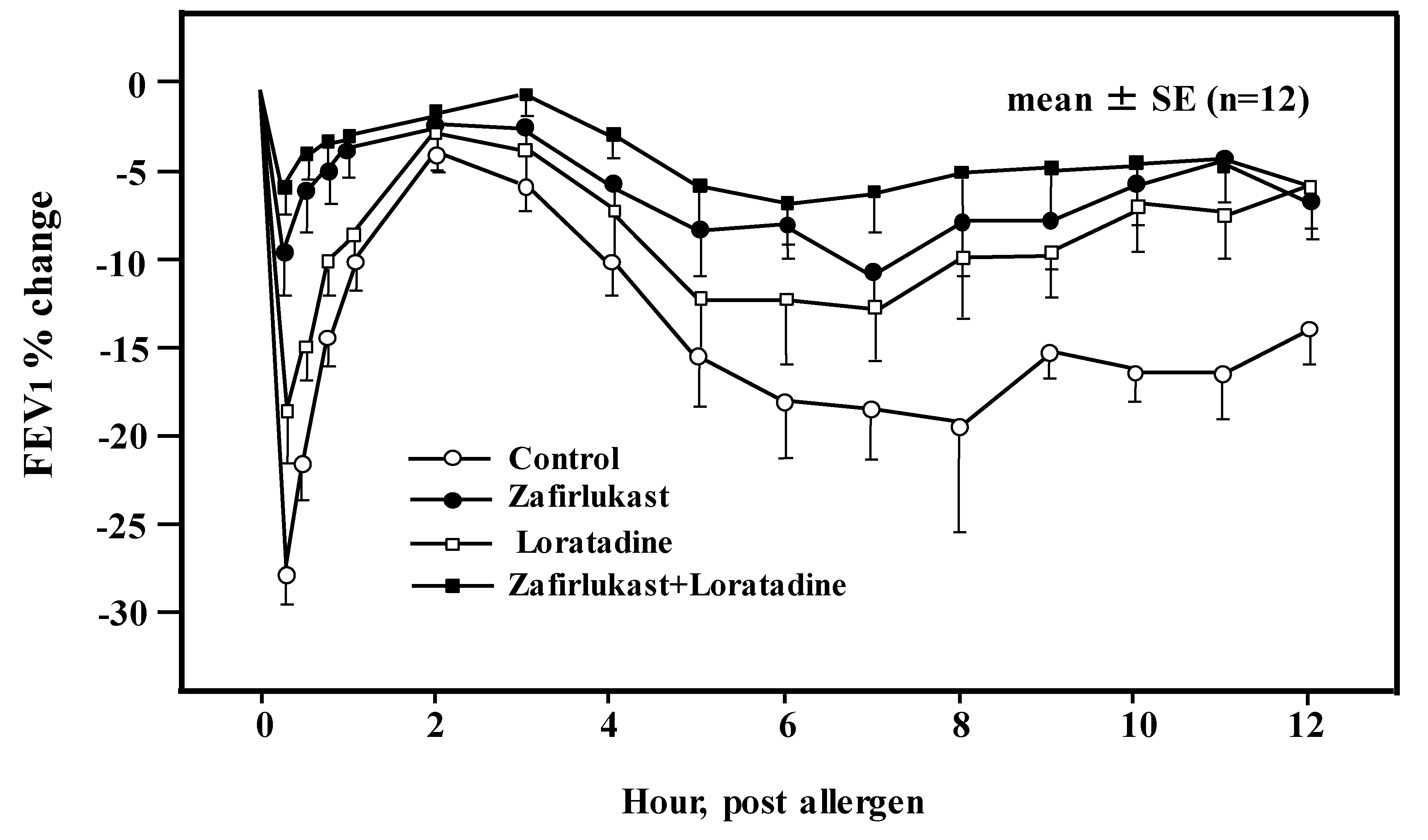
5. H1RA in Asthma Therapy
6. The Immunological Roles of Histamine in Allergic Reactions
7. A New Insight on the Immunological Pathway in Asthma
8. A New Aspect of H1RA and H4RA in Asthma Therapy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, J.V. The nature of the pulmonary receptors excited by antihistamines. Br. J. Pharmacol. Chemother. 1952, 7, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Tucker, A.; Weir, E.K.; Reeves, J.T.; Grover, R.F. Histamine H1- and H2-receptors in pulmonary and systemic vasculature of the dog. Am. J. Physiol. 1975, 229, 1008–1013. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ichinose, M.; Barnes, P.J. Inhibitory histamine H3-receptors on cholinergic nerves in human airways. Eur. J. Pharmacol. 1989, 163, 383–386. [Google Scholar] [CrossRef]
- Kay, L.J.; Suvarna, S.K.; Peachell, P.T. Histamine H4 receptor mediates chemotaxis of human lung mast cells. Eur. J. Pharmacol. 2018, 837, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Curry, J.J. The effect of antihistamine substances and other drugs on histamine bronchoconstriction in asthmatic subjects. J. Clin. Investig. 1946, 25, 792–799. [Google Scholar] [CrossRef]
- Drazen, J.M.; Schneider, M.W. Comparative responses of tracheal spirals and parenchymal strips to histamine and carbachol in vitro. J. Clin. Investig. 1978, 61, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Mirbahar, K.B.; Oliver, W., Jr.; Eyre, P.; Wanner, A. Characterization of H1- and H2-receptor function in pulmonary and systemic circulations of sheep. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1982, 53, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Nadel, J.A.; Davis, B.; Phipps, R.J. Control of mucus secretion and ion transport in airways. Annu. Rev. Physiol. 1979, 41, 369–381. [Google Scholar] [CrossRef] [PubMed]
- White, M.V. The role of histamine in allergic diseases. J. Allergy Clin. Immunol. 1990, 86, 599–605. [Google Scholar] [CrossRef]
- Casale, T.B.; Wood, D.; Richerson, H.B.; Trapp, S.; Metzger, W.J.; Zavala, D.; Hunninghake, G.W. Elevated bronchoalveolar lavage fluid histamine levels in allergic asthmatics are associated with methacholine bronchial hyperresponsiveness. J. Clin. Investig. 1987, 79, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, M.; Ida, S.; Shindoh, Y.; Ishihara, T.; Takishima, T. Mast cells in bronchoalveolar lumen of patients with bronchial asthma. Am. Rev. Respir. Dis. 1984, 129, 1000–1005. [Google Scholar]
- Dale, H.H.; Laidlaw, P.P. Histamine shock. J. Physiol. 1919, 52, 355–390. [Google Scholar] [CrossRef]
- Ishizaka, T.; Ishizaka, K.; Tomioka, H. Release of histamine and slow reacting substance of anaphylaxis (SRS-A) by IgE-anti-IgE reactions on monkey mast cells. J. Immunol. 1972, 108, 513–520. [Google Scholar]
- Curry, J.J. The action of histamine on the respiratory tract in normal and asthmatic subjects. J. Clin. Investig. 1946, 25, 785–791. [Google Scholar] [CrossRef]
- Hogg, J.C.; Paré, P.D.; Boucher, R.C.; Michoud, M.C. The pathophysiology of asthma. Can. Med. Assoc. J. 1979, 121, 409–414. [Google Scholar]
- Rafferty, P.; Beasley, R.; Holgate, S.T. The contribution of histamine to immediate bronchoconstriction provoked by inhaled allergen and adenosine 5′ monophosphate in atopic asthma. Am. Rev. Respir. Dis. 1987, 136, 369–373. [Google Scholar] [CrossRef]
- Yamauchi, K. Regulation of gene expression of l-histidine decarboxylase and histamine N-methyl-transferase, and its relevance to the pathogenesis of bronchial asthma. Nihon Rinsho 1996, 54, 377–388. [Google Scholar]
- Sekizawa, K.; Nakazawa, H.; Morikawa, M.; Yamauchi, K.; Maeyama, K.; Watanabe, T.; Sasaki, H. Histamine N-methyltransferase inhibitor potentiates histamine- and antigen-induced airway microvascular leakage in guinea pigs. J. Allergy Clin. Immunol. 1995, 96, 910–916. [Google Scholar] [CrossRef]
- Yamauchi, K.; Sekizawa, K.; Suzuki, H.; Nakazawa, H.; Ohkawara, Y.; Katayose, D.; Ohtsu, H.; Tamura, G.; Shibahara, S.; Takemura, M.; et al. Structure and function of human histamine N-methyltransferase: Critical enzyme in histamine metabolism in airway. Am. J. Physiol. 1994, 267, L342–L349. [Google Scholar] [CrossRef]
- Anvari, S.; Vyhlidal, C.A.; Dai, H.; Jones, B.L. Genetic Variation along the Histamine Pathway in Children with Allergic versus Nonallergic Asthma. Am. J. Respir. Cell Mol. Biol. 2015, 53, 802–809. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and histamine receptors in Mast cell-mediated allergy and inflammation: The Hunt for New therapeutic Targets. Front. Immunol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Ohtsu, H. Progress in allergy signal research on mast cells: The role of histamine in immunological and cardiovascular disease and the transporting system of histamine in the cell. J. Pharmacol. Sci. 2008, 106, 347–353. [Google Scholar] [CrossRef]
- Ogasawara, M.; Yamauchi, K.; Satoh, Y.; Yamaji, R.; Inui, K.; Jonker, J.W.; Schinkel, A.H.; Maeyama, K. Recent advances in Molecular Pharmacology of the histamine systems: Organic cation Transporters as a Histamine transporter and Histamine Metabolism. J. Pharmacol. Sci. 2006, 101, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Endou, H. The SLC22 drug transporter family. Pflugers Arch. 2004, 447, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Yanai, K. Histamine Clearance through Polyspecific Transporters in the Brain. Handb. Exp. Pharmacol. 2017, 241, 173–187. [Google Scholar] [PubMed]
- Zwart, R.; Verhaagh, S.; Buitelaar, M.; Popp-Snijders, C.; Barlow, D.P. Impaired activity of the extraneuronal monoamine transporter system known as uptake-2 in Orct3/Slc22a3-deficient mice. Mol. Cell Biol. 2001, 21, 4188–4196. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Hata, R.; Ogasawara, M.; Cao, F.; Kameda, K.; Yamauchi, K.; Schinkel, A.H.; Maeyama, K.; Sakanaka, M. Targeted disruption of organic cation transporter 3(Oct3) ameliorates ischemic brain damage through modulating histamine and regulatory T cells. J. Cereb. Blood Flow Metab. 2012, 32, 1897–1908. [Google Scholar] [CrossRef]
- Yamauchi, K.; Shikanai, T.; Nakamura, Y.; Kobayashi, H.; Ogasawara, M.; Maeyama, K. Roles of histamine in the pathogenesis of bronchial asthma and reevaluation of the clinical usefulness of antihistamines. Yakugaku Zasshi 2011, 131, 185–191. [Google Scholar] [CrossRef][Green Version]
- Sakata, T.; Anzai, N.; Kimura, T.; Miura, D.; Fukutomi, T.; Takeda, M.; Sakurai, H.; Endou, H. Functional analysis of human Organic cation Transporter OCT3(SLC22A3) polymorphisms. J. Pharmacol. Sci. 2010, 113, 263–266. [Google Scholar] [CrossRef]
- Lee, N.; Hebert, M.F.; Wagner, D.J.; Easterling, T.R.; Liang, C.J.; Rice, K.; Wang, J. Organic Cation Transporter 3 facilitates Fetal Exposure to Metformin during Pregnancy. Mol. Pharmacol. 2018, 94, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Mahrooz, A.; Alizadeh, A.; Hashemi-Soteh, M.B.; Ghaffari-Cherati, M.; Hosseyni-Talei, S.R. Polymorphic Variants rs3088442 and rs2292334 in the Organic cation Transporter 3(OCT3) gene and Susceptibility Against Type 2 Diabetes: Role of their Interaction. Arch. Med. Res. 2017, 48, 162–168. [Google Scholar] [CrossRef]
- Hosseyni-Talei, S.R.; Mahrooz, A.; Hashemi-Soteh, M.B.; Ghaffari-Cherati, M.; Alizadeh, A. Association between the synonymous variant organic cation transporter 3(OCT3)-1233>A and the glycemic response following metformin therapy in patients with type 2 diabetes. Iran. J. Basic Med. Sci. 2017, 20, 250–255. [Google Scholar]
- Ghaffari-Cherati, M.; Mahrooz, A.; Hashemi-Soteh, M.B.; Hosseyni-Talei, S.R.; Alizadeh, A.; Nakhaei, S.M. Allele frequency and genotype distribution of a common variant in the 3′-untranslated region of the SLC22A3 gene in patients with type 2 diabetes: Association with response to metformin. J. Res. Med. Sci. 2016, 21, 92. [Google Scholar] [CrossRef]
- Zaharenko, L.; Kalnina, I.; Geldnere, K.; Konrade, I.; Grinberga, S.; Židzik, J.; Javorský, M.; Lejnieks, A.; Nikitina-Zake, L.; Fridmanis, D.; et al. Single nucleotide polymorphisms in the intergenic region between metformin transporter OCT2 and OCT3 coding genes are associated with short-term response to metformin monotherapy in type 2 diabetes mellitus patients. Eur. J. Endocrinol. 2016, 175, 531–540. [Google Scholar] [CrossRef]
- Chen, E.C.; Liang, X.; Yee, S.W.; Geier, E.G.; Stocker, S.L.; Chen, L.; Giacomini, K.M. Targeted disruption of organic cation transporter 3 attenuates the pharmacological response to metformin. Mol. Pharmacol. 2015, 88, 75–83. [Google Scholar] [CrossRef]
- Chen, L.; Hong, C.; Chen, E.C.; Yee, S.W.; Xu, L.; Almof, E.U.; Wen, C.; Fujii, K.; Johns, S.J.; Stryke, D.; et al. Genetics and epigenetic regulation of the organic cation transporter 3, SLC22A3. Pharm. J. 2013, 13, 110–120. [Google Scholar] [CrossRef]
- Aoyama, N.; Takahashi, N.; Kitaichi, K.; Ishihara, R.; Saito, S.; Maeno, N.; Ji, X.; Takagi, K.; Sekina, Y.; Iyo, M.; et al. Association between gene polymorphisms of SLA22A3 and methamphetamine use disorder. Alchol. Clin. Exp. Res. 2006, 30, 1644–1649. [Google Scholar] [CrossRef]
- Ferstl, R.; Frei, R.; Barcik, W.; Sciavi, E.; Wanke, K.; Ziegler, M.; Rodriguez-Perez, N.; Groeger, D.; Konieczna, P.; Zeiter, S.; et al. Histamine receptor 2 modified iNKT cell activity within the inflamed lung. Allergy 2017, 72, 1925–1935. [Google Scholar] [CrossRef]
- Schneider, E.; Machavoine, F.; Plěau, J.M.; Berton, A.F.; Thurmond, R.L.; Otsu, H.; Watanabe, T.; Schinkel, A.H.; Dy, M. Organic cation transporter 3 modulates murine basophil functions by controlling intracellular histamine levels. J. Exp. Med. 2005, 202, 387–393. [Google Scholar] [CrossRef]
- Herxheimer, H. Antihistamines in bronchial asthma. Proc. R. Soc. Med. 1949, 42, 615–629. [Google Scholar] [CrossRef]
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzaki, H.; Fujieda, S.; Masuyama, K. Japanese Society of Allergology. Japanese guidelines for allergic rhinitis 2017. Allergol. Int. 2017, 66, 205–219. [Google Scholar] [CrossRef]
- Wheatley, L.M.; Togias, A. Clinical practice. Allergic rhinitis. N. Engl. J. Med. 2015, 372, 456–463. [Google Scholar] [CrossRef]
- Zuberbier, T.; Aberer, W.; Asero, R. The EAACI/GA2LEN/EDF/WAO guideline for the definition, classification, diagnosis and management of urticaria. Allergy 2018, 73, 1393–1414. [Google Scholar] [CrossRef]
- The Global Initiative for Asthma; Global Strategy for Asthma Management and Prevention (2018 Update). Available online: https://ginasthma.org/about-us/ (accessed on 3 February 2019).
- Roquet, A.; Dahlén, B.; Kumlin, M.; Ihre, E.; Anstrén, G.; Binks, S.; Dahlén, S.E. Combined antagonism of leukotrienes and histamine produces predominant inhibition of allergen-induced early and late phase airway obstruction in asthmatics. Am. J. Respir. Crit. Care Med. 1997, 155, 1856–1863. [Google Scholar] [CrossRef]
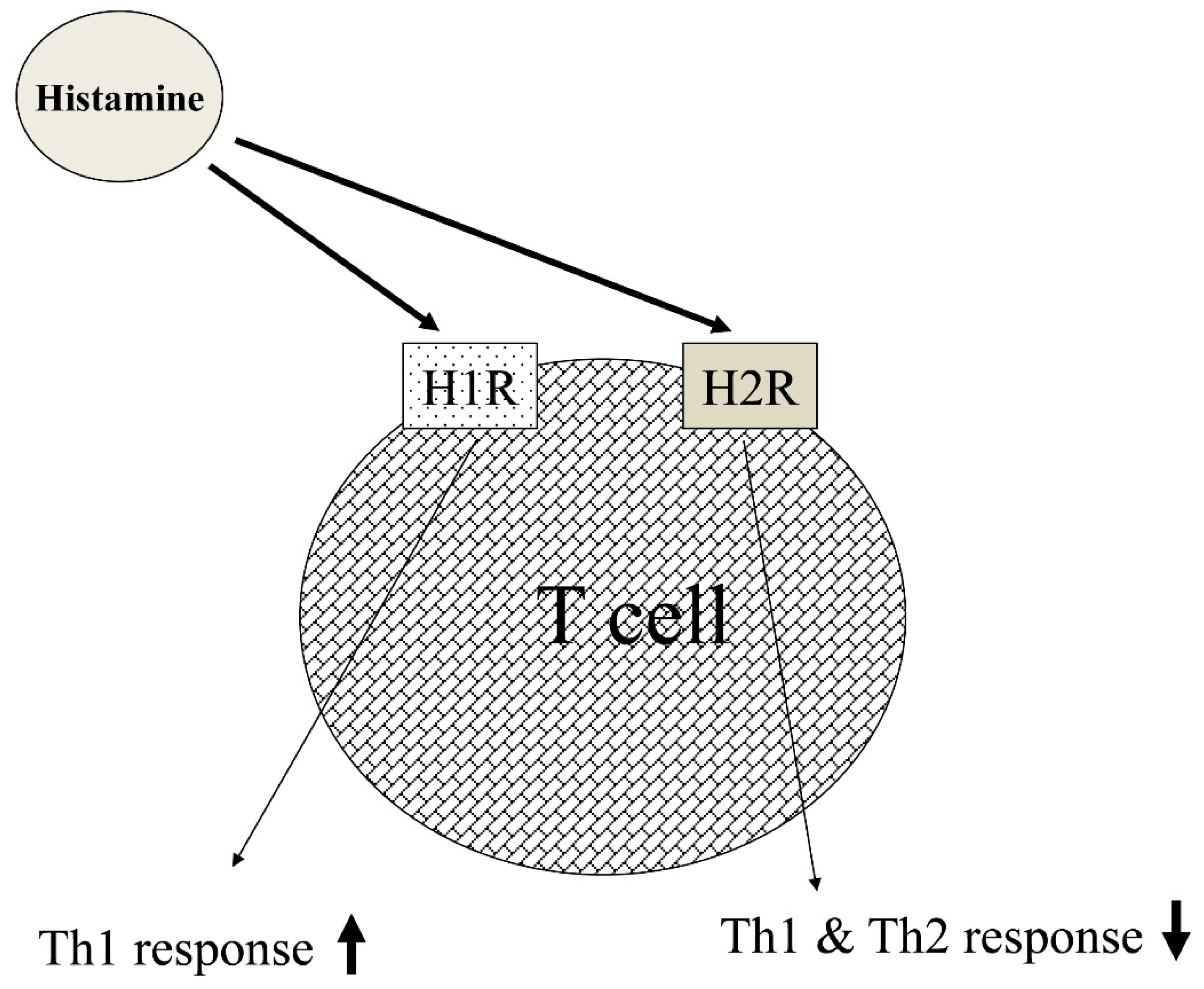
- Jutel, M.; Watanabe, T.; Klunker, S.; Akdis, M.; Thomet, O.A.; Malolepszy, J.; Zak-Nejmark, T.; Koga, R.; Kobayashi, T.; Blaser, K.; et al. Histamine regulates T-cell and antibody responses by differential expression of H1 and H2 receptors. Nature 2001, 413, 420–425. [Google Scholar] [CrossRef]
- Del Prete, G.F.; De Carli, M.; Mastromauro, C. Purified protein derivate of Mycobacterium tuberculosis and excretory-secretory antigen(s) of Toxocara canis expand in vitro human T cells with stable and opposite (type 1 T helper or type 2 T helper) profiles of cytokine production. J. Clin. Investig. 1991, 88, 344–350. [Google Scholar] [CrossRef]
- Robinson, D.S.; Hamid, Q.; Ying, S.; Tsicopoulos, A.; Barkans, J.; Bentley, A.M.; Corrigan, C.; Durham, S.R.; Kay, A.B. Predominant Th2-like bronchoalveolar T lymphocyte population in atopic asthma. N. Engl. J. Med. 1992, 326, 298–304. [Google Scholar] [CrossRef]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N.; et al. Mice lacking histidine decarboxylase exhibit abnormal mast cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef]
- Koarai, A.; Ichinose, M.; Ishigaki-Suzuki, S.; Yamagata, S.; Sugiura, H.; Sakurai, E.; Makabe-Kobayashi, Y.; Kuramasu, A.; Watanabe, T.; Shirato, K.; et al. Disruption of L-histidine decarboxylase reduces airway eosinophilia but not hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2003, 167, 758–763. [Google Scholar] [CrossRef]
- O’Reilly, M.; Alpert, R.; Jenkinson, S.; Gladue, R.P.; Foo, S.; Trim, S.; Peter, B.; Trevethick, M.; Fidock, M. Identification of a histamine H4 receptor on human eosinophils—Role in eosinophil chemotaxis. J. Recept. Signal Transduct. Res. 2002, 22, 431–448. [Google Scholar] [CrossRef]
- Makabe-Kobayashi, Y.; Hori, Y.; Adachi, T.; Ishigaki-Suzuki, S.; Kikuchi, Y.; Kagaya, Y.; Shirato, K.; Nagy, A.; Ujike, A.; Takai, T.; et al. The control effect of histamine on body temperature and respiratory function in IgE-dependent systemic anaphylaxis. J. Allergy Clin. Immunol. 2002, 110, 298–303. [Google Scholar] [CrossRef]
- Yamauchi, K.; Piao, H.M.; Nakadate, T.; Shikanai, T.; Nakamura, Y.; Ito, H.; Mouri, T.; Kobayashi, H.; Maesawa, C.; Sawai, T.; et al. Enhanced goblet cell hyperplasia in HDC knockout mice with allergic airway inflammation. Allergol. Int. 2009, 58, 125–134. [Google Scholar]
- Nakanishi, A.; Morita, S.; Iwashita HSagiya, Y.; Ashida, Y.; Shirafuji, H.; Fujisawa, Y.; Nishimura, O.; Fujino, M. Role of gob-5 in mucus overproduction and airway hyperresponsiveness in asthma. Proc. Natl. Acad. Sci. USA 2001, 98, 5175–5180. [Google Scholar] [CrossRef]
- Gosset, P.; Tsicopoulos, A.; Wallaert, B.; Joseph, M.; Capron, A.; Tonnel, A.B. Tumor necrosis factor-α and interleukin-6 production by human mononuclear phagocytes from allergic asthmatics after IgEdependent stimulation. Am. Rev. Respir. Dis. 1992, 146, 768–774. [Google Scholar] [CrossRef]
- Busse, P.J.; Zhang, T.F.; Srivastava KLin, B.P.; Schofield, B.; Sealfon, S.C.; Li, X.M. Chronic exposure to TNF-alpha increases airway mucus gene expression in vivo. J. Allergy Clin. Immunol. 2005, 116, 1256–1263. [Google Scholar] [CrossRef]
- Hirose, K.; Iwata, A.; Tamachi, T.; Nakajima, H. Allergic airway inflammation: Key players beyond the Th2 cell pathway. Immunol. Rev. 2017, 278, 145–161. [Google Scholar] [CrossRef]
- Moldaver, D.M.; Larché, M.; Rudulier, C.D. An Update on Lymphocyte Subtypes in Asthma and Airway Disease. Chest 2017, 151, 1122–1130. [Google Scholar] [CrossRef]
- Li, B.W.; Hendriks, R.W. Group 2 innate lymphoid cells in lung inflammation. Immunology 2013, 140, 281–287. [Google Scholar] [CrossRef]
- Barlow, J.L.; McKenzie, A.N. Type-2 innate lymphoid cells in human allergic disease. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 397–403. [Google Scholar] [CrossRef]
- Becker, A.B.; Abrams, E.M. Asthma guidelines: The Global Initiative for Asthma in relation to national guidelines. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 99–103. [Google Scholar] [CrossRef]
- Ichinose, M.; Sugiura, H.; Nagase, H.; Yamaguchi, M.; Inoue, H.; Sagara, H.; Tamaoki, J.; Tohda, Y.; Munakata, M.; Yamauchi, K.; et al. Japanese Society of Allergology Japanese guidelines for adult asthma 2017. Allergol. Int. 2017, 66, 163–189. [Google Scholar] [CrossRef]
- Busse, W.; Corren, J.; Lanier, B.Q.; McAlary, M.; Fowler-Taylor, A.; Cioppa, G.D.; van As, A.; Gupta, N. Omalizumab, anti-IgE recombinant humanized monoclonal antibody, for the treatment of severe allergic asthma. J. Allergy Clin. Immunol. 2001, 108, 184–190. [Google Scholar] [CrossRef]
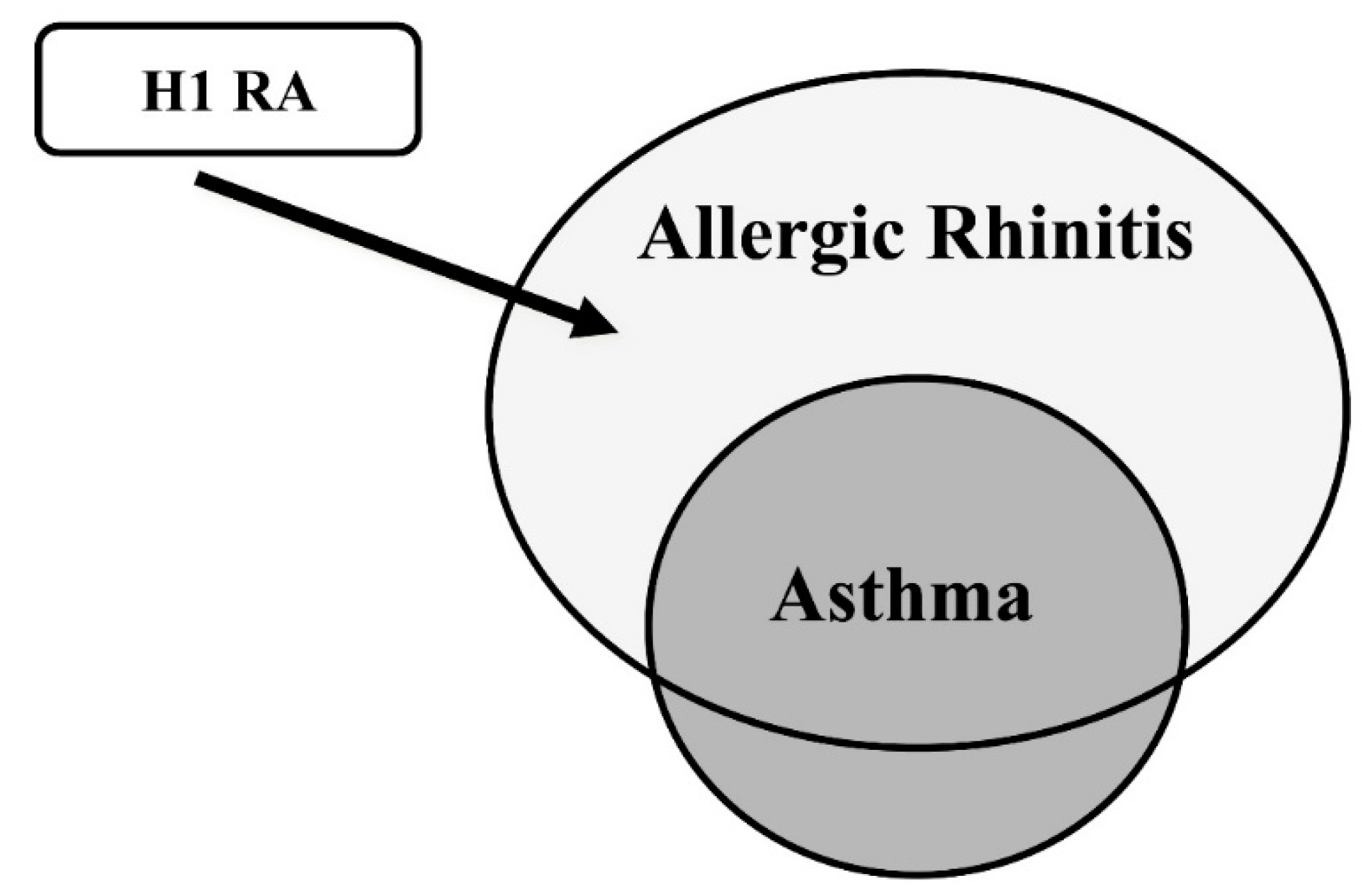
- Yamauchi, K.; Tamura, G.; Akasaka, T.; Chiba, T.; Honda, K.; Kishi, M.; Kobayashi, H.; Kuronuma, T.; Matsubara, A.; Morikawa, T.; et al. Analysis of the comorbidity of bronchial asthma and allergic rhinitis by questionnaire in 10,009 patients. Allergol. Int. 2009, 58, 55–61. [Google Scholar] [CrossRef]
- Buckland, K.F.; Williams, T.J.; Conroy, D.M. Histamine induces cytoskeletal changes in human eosinophils via the H(4) receptor. Br. J. Pharmacol. 2003, 140, 1117–1127. [Google Scholar] [CrossRef]
- Neumann, D.; Beermann, S.; Seifert, R. Does the histamine H4 receptor have a pro- or anti-inflammatory role in murine bronchial asthma? Pharmacology 2010, 85, 217–223. [Google Scholar] [CrossRef]
- Hofstra, C.L.; Desai, P.J.; Thurmond, R.L.; Fung-Leung, W.P. Histamine H4 receptor mediates chemotaxis and calcium mobilization of mast cells. J. Pharmacol. Exp. Ther. 2003, 305, 1212–1221. [Google Scholar] [CrossRef]
- Dunford, P.J.; O’Donnell, N.; Riley, J.P.; Williams, K.N.; Karlsson, L.; Thurmond, R.L. The Histamine H4 Receptor Mediates Allergic Airway Inflammation by Regulating the Activation of CD4+ T Cells. J. Immunol. 2006, 176, 7062–7070. [Google Scholar] [CrossRef]
- Cowden, J.M.; Riley, J.P.; Ma, J.Y.; Thurmond, R.L.; Dunford, P.J. Histamine H4 receptor antagonism diminishes existing airway inflammation and dysfunction via modulation of Th2 cytokines. Respir. Res. 2010, 11, 86. [Google Scholar] [CrossRef]
- Thurmond, R.L.; Chen, B.; Dunford, P.J.; Greenspan, A.J.; Karlsson, L.; La, D.; Ward, P.; Xu, X.L. Clinical and preclinical characterization of the histamine H(4) receptor antagonist JNJ-39758979. J. Pharmacol. Exp. Ther. 2014, 349, 176–184. [Google Scholar] [CrossRef]
- Kollmeier, A.P.; Barnathan, E.S.; O’Brien, C.; Chen, B.; Xia, Y.K.; Zhou, B.; Loza, M.J.; Silkoff, P.E.; Ge, M.; Thurmond, R.L. A phase 2a study of toreforant, a histamine H4 receptor antagonist, in eosinophilic asthma. Ann. Allergy Asthma Immunol. 2018, 121, 568–574. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step 1 | Step 2 | Step 3 | Step 4 | Step 5 | |
|---|---|---|---|---|---|
| Controllers | Low dose ICS | Low dose ICS/LABA | Med/high ICS/LABA | Add-on Tiotropium, Anti-IgE, Anti-IL-5 | |
| Other controllers options | ICS (low dose) | LTRA Low dose Theophylline | Med/high dose ICS; low dose ICS + LTRA (or + Theophylline) | Add Tiotropium Med/high dose ICS; low dose ICS + LTRA (or + Theophylline) | Add low dose OCS |
| Relievers | SABA | SABA | SABA or low dose ICS/formoterol | ||
| Number | Yes | No | n.d. | |
|---|---|---|---|---|
| Adult asthma | 2781 | 1693 (60.8%) * | 1044 (37.5%) | 44 (1.6%) |
| Child asthma | 3283 | 2238 (68.2%) ** | 1035 (31.5%) | 10 (0.3%) |
| Allergic rhinitis (AR) | 3945 | 1935(49.0%) | 2010 (51.0%) | – |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. https://doi.org/10.3390/ijms20071733
Yamauchi K, Ogasawara M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. International Journal of Molecular Sciences. 2019; 20(7):1733. https://doi.org/10.3390/ijms20071733
Chicago/Turabian StyleYamauchi, Kohei, and Masahito Ogasawara. 2019. "The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy" International Journal of Molecular Sciences 20, no. 7: 1733. https://doi.org/10.3390/ijms20071733
APA StyleYamauchi, K., & Ogasawara, M. (2019). The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. International Journal of Molecular Sciences, 20(7), 1733. https://doi.org/10.3390/ijms20071733