The Interplay between miRNA-Related Variants and Age-Related Macular Degeneration: EVIDENCE of Association of MIR146A and MIR27A

 ,
,  ,
,
Abstract
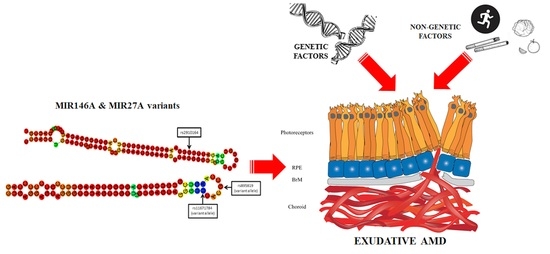
:
1. Introduction
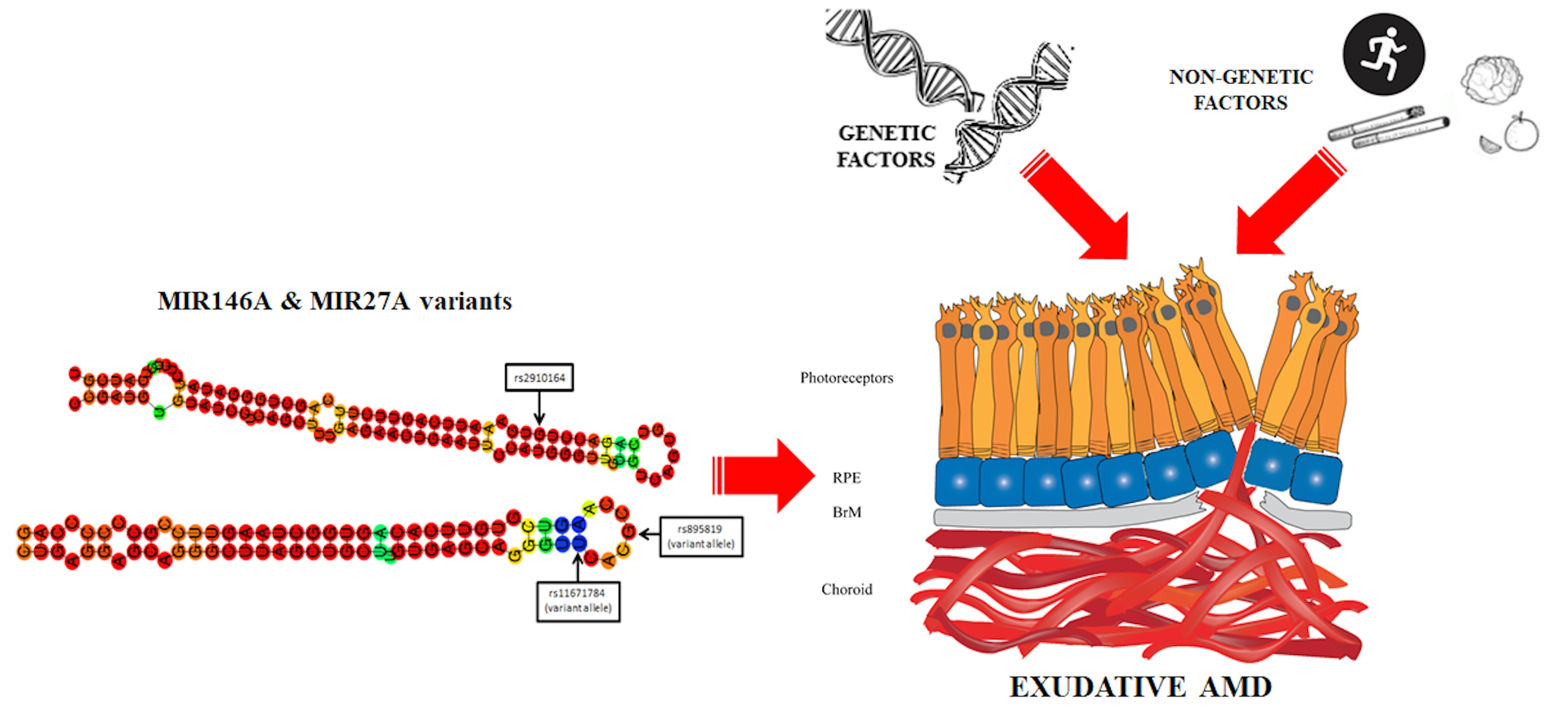
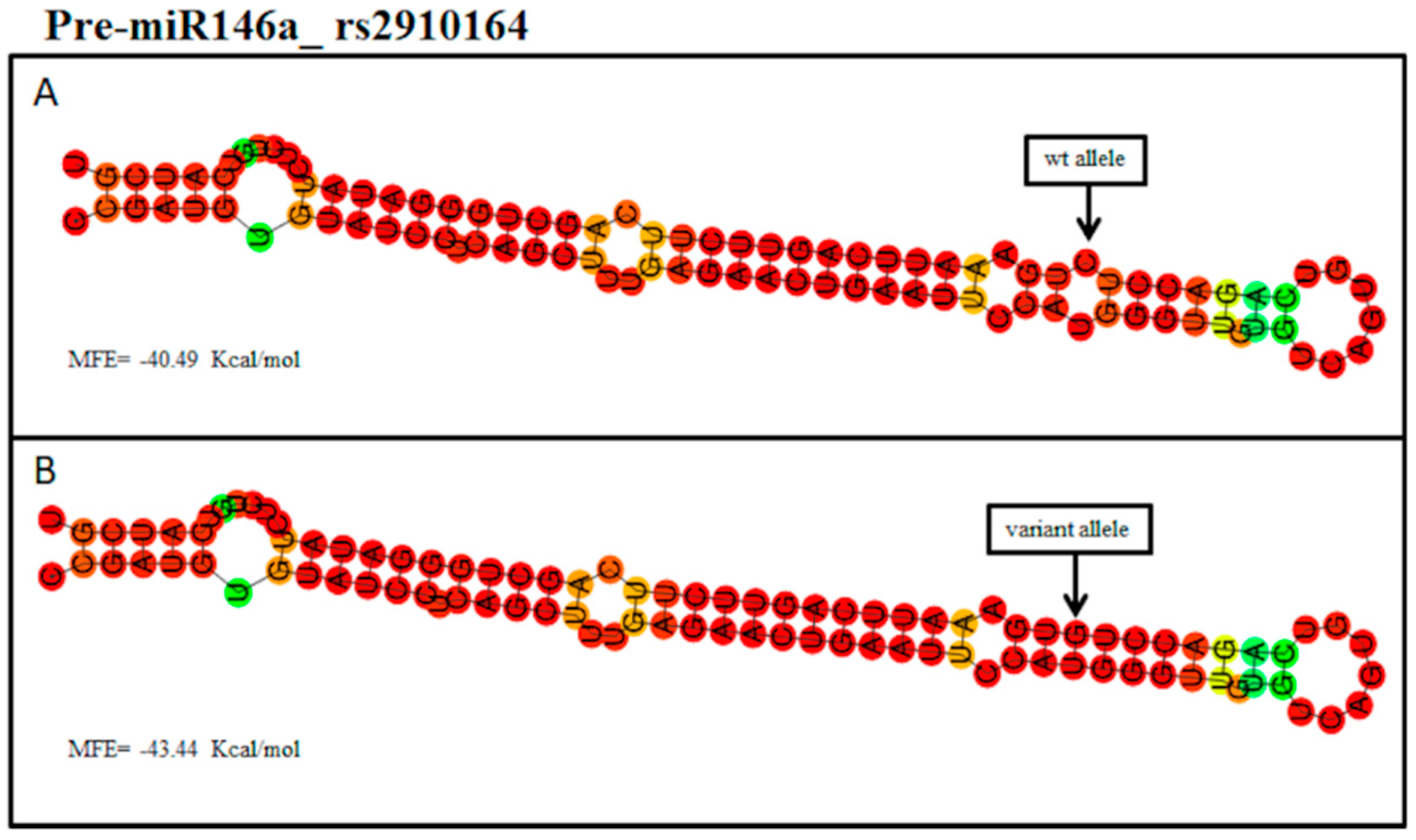
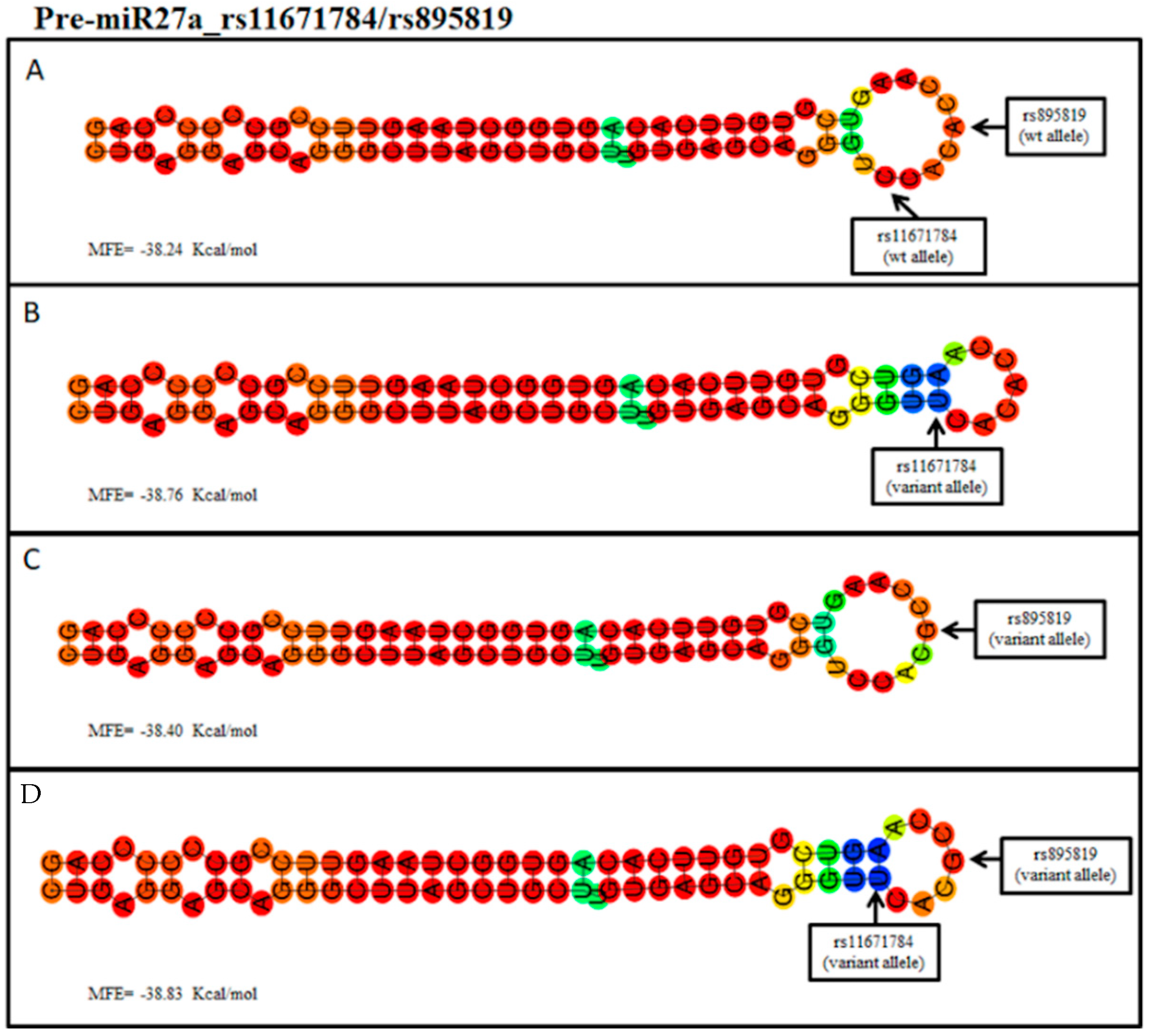
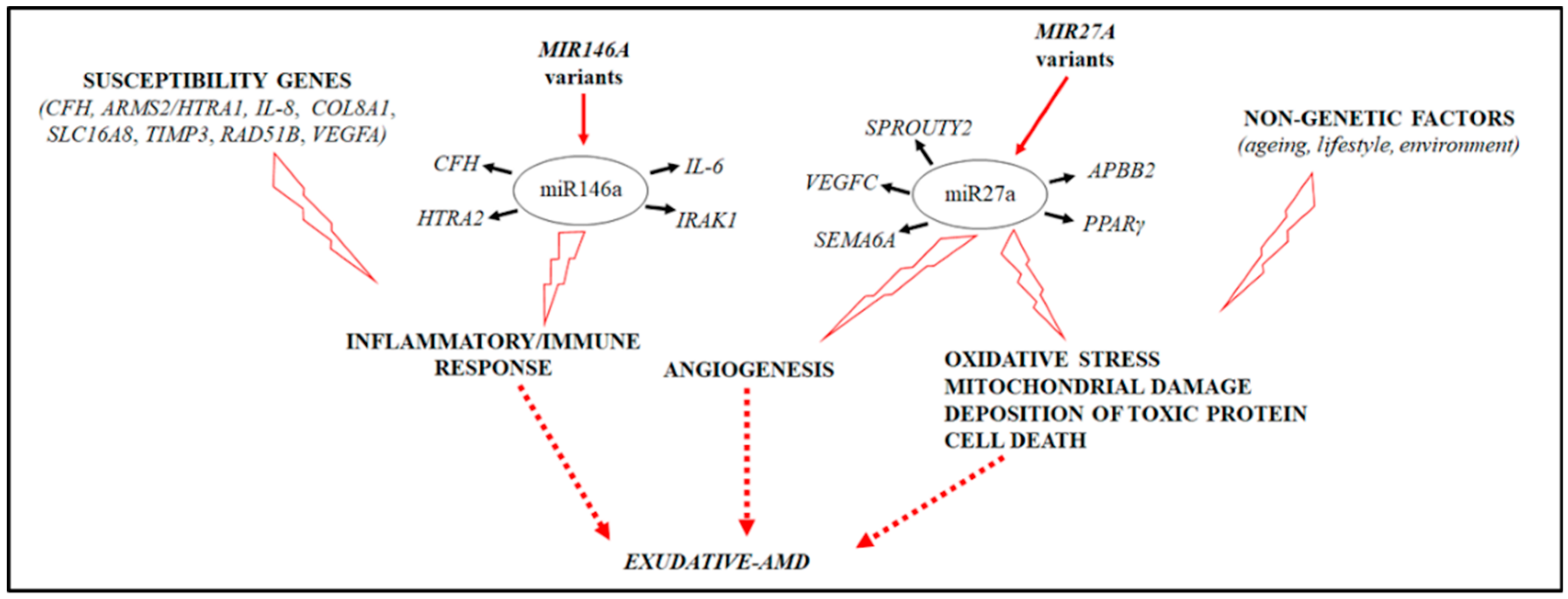
2. Results
3. Discussion
4. Material and Methods
4.1. Study Cohort and DNA Extraction
4.2. Genotyping Analysis
4.3. Biostatistical and Bioinformatic Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cascella, R.; Strafella, C.; Caputo, V.; Errichiello, V.; Zampatti, S.; Milano, F.; Potenza, S.; Mauriello, S.; Novelli, G.; Ricci, F.; et al. Towards the application of precision medicine in age-related macular degeneration. Prog. Retin. Eye Res. 2017, 63, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Strafella, C.; Caputo, V.; Galota, M.R.; Zampatti, S.; Marella, G.; Mauriello, S.; Cascella, R.; Giardina, E. Application of precision medicine in neurodegenerative diseases. Front. Neurol. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Cascella, R.; Strafella, C.; Germani, C.; Novelli, G.; Ricci, F.; Zampatti, S.; Giardina, E. The genetics and the genomics of primary congenital glaucoma. Biomed. Res. Int. 2015, 2015, 321291. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.N.; Pericak-Vance, M.A.; Haines, J.L. The impact of the human genome project on complex disease. Genes 2014, 5, 518–535. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.G.; Fariss, R.N.; Stambolian, D.; Abecasis, G.R.; Curcio, C.A.; Swaroop, A. Age-related macular degeneration: Genetics and biology coming together. Annu. Rev. Genomics Hum. Genet. 2014, 15, 151–171. [Google Scholar] [CrossRef]
- Horie-Inoue, K.; Inoue, S. Genomic aspects of age-related macular degeneration. Biochem. Biophys. Res. Commun. 2014, 452, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.G.; Igl, W.; Bailey, J.N.; Grassmann, F.; Sengupta, S.; Bragg-Gresham, J.L.; Burdon, K.P.; Hebbring, S.J.; Wen, C.; Gorski, M.; et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2016, 48, 134e–143e. [Google Scholar] [CrossRef] [PubMed]
- Cascella, R.; Strafella, C.; Longo, G.; Ragazzo, M.; Manzo, L.; De Felici, C.; Errichiello, V.; Caputo, V.; Viola, F.; Eandi, C.M.; et al. Uncovering genetic and non-genetic biomarkers specific for exudative age-related macular degeneration: Significant association of twelve variants. Oncotarget 2017, 9, 7812–7821. [Google Scholar] [CrossRef]
- Ricci, F.; Zampatti, S.; D’Abbruzzi, F.; Missiroli, F.; Martone, C.; Lepre, T.; Pietrangeli, I.; Sinibaldi, C.; Peconi, C.; Novelli, G.; et al. Typing of ARMS2 and CFH in age-related macular degeneration: Case-control study and assessment of frequency in the Italian population. Arch. Ophthalmol. 2009, 127, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Lepre, T.; Cascella, R.; Missiroli, F.; De Felici, C.; Taglia, F.; Zampatti, S.; Cusumano, A.; Ricci, F.; Giardina, E.; Eandi, C.M.; et al. Polymorphisms in ARMS2 (LOC387715) and LOXL1 Genes in the Japanese with age-related macular degeneration. Am. J. Ophthalmol. 2011, 152, 325–326. [Google Scholar] [CrossRef]
- Ricci, F.; Staurenghi, G.; Lepre, T.; Missiroli, F.; Zampatti, S.; Cascella, R.; Borgiani, P.; Marsella, L.T.; Eandi, C.M.; Cusumano, A.; et al. Haplotypes in IL-8 gene are associated to age-related macular degeneration: A case-control study. PLoS ONE 2013, 8, e66978. [Google Scholar] [CrossRef]
- Sobrin, L.; Seddon, J.M. Nature and nurture- genes and environment- predict onset and progression of macular degeneration. Prog. Retin. Eye Res. 2014, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lambert, N.G.; ElShelmani, H.; Singh, M.K.; Mansergh, F.C.; Wride, M.A.; Padilla, M.; Keegan, D.; Hogg, R.E.; Ambati, B.K. Risk factors and biomarkers of age-related macular degeneration. Prog. Reti. Eye Res. 2016, 54, 64–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SanGiovanni, J.P.; SanGiovanni, P.M.; Sapieha, P.; De Guire, V. miRNAs, single nucleotide polymorphisms (SNPs) and age-related macular degeneration (AMD). Clin. Chem. Lab. Med. 2017, 55, 763–775. [Google Scholar] [CrossRef]
- Gemenetzi, M.; Lotery, A.J. The role of epigenetics in age-related macular degeneration. Eye 2014, 28, 1407–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, G.L.; Platania, C.B.M.; Drago, F.; Salomone, S.; Ragusa, M.; Barbagallo, C.; Di Pietro, C.; Purrello, M.; Reibaldi, M.; Avitabile, T.; et al. Retinal and circulating miRNAs in age-related macular degeneration: An in vivo animal and human study. Front. Pharmacol. 2017, 8, 168. [Google Scholar] [CrossRef]
- Lukiw, W.J.; Surjyadipta, B.; Dua, P.; Alexandrov, P.N. Common micro RNAs (miRNAs) target complement factor H (CFH) regulation in Alzheimer’s disease (AD) and in age-related macular degeneration (AMD). Int. J. Biochem. Mol. Biol. 2012, 3, 105–116. [Google Scholar] [PubMed]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Orjalo, A.V.; Rodier, F.; Lithgow, G.J.; Campisi, J. MicroRNAs miR-145a/b negatively modulates the senescence-associated inflammatory mediators IL-6 and IL-8. Aging 2009, 1, 402–411. [Google Scholar] [CrossRef]
- Kutty, R.K.; Nagineni, C.N.; Samuel, W.; Vijayasarathy, C.; Jaworski, C.; Duncan, T.; Cameron, J.E.; Flemington, E.K.; Hooks, J.J.; Redmond, T.M. Differential regulation of microRNA-146a and microRNA-146b-5p in human retinal pigment epithelial cells by interleukin-1β, tumor necrosis factor-α, and interferon-γ. Mol. Vis. 2013, 19, 737–750. [Google Scholar] [PubMed]
- Wagh, A.R.; Bose, K. Emerging Roles of Mitochondrial Serine Protease HtrA2 in Neurodegeneration. In Proteases in Physiology and Pathology; Chakraborti, S., Dhalla, N., Eds.; Springer Nature: Singapore, 2017; Volume 15, pp. 325–353. [Google Scholar] [CrossRef]
- Ding, X.; Patel, M.; Shen, D.; Herzlich, A.A.; Cao, X.; Villasmil, R.; Klupsch, K.; Tuo, J.; Downward, J.; Chan, C.C. Enhanced HtrA2/Omi expression in oxidative injury to retinal pigment epithelial cells and murine models of neurodegeneration. Invest. Ophthalmol. Vis. Sci. 2009, 50, 4957–4966. [Google Scholar] [CrossRef] [PubMed]
- Kaidonis, G.; Gillies, M.C.; Abhary, S.; Liu, E.; Essex, R.W.; Chang, J.H.; Pal, B.; Sivaprasad, S.; Pefkianaki, M.; Daniell, M.; et al. A single-nucleotide polymorphism in the MicroRNA-146a gene is associated with diabetic nephropathy and sight-threatening diabetic retinopathy in Caucasian patients. Acta Diabetol. 2016, 53, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Chatzikyriakidou, A.; Voulgari, P.; Georgiou, I.; Drosos, A.A. The role of microRNA-146a (miR-146a) and its target IL-1R-associated kinase (IRAK1) in psoriatic arthritis susceptibility. Scand. J. Immunol. 2010, 71, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yi, X.; Guo, S.; Shi, Q.; Wei, C.; Li, X.; Gao, L.; Wang, G.; Gao, T.; Wang, L.; et al. A single-nucleotide polymorphism of miR-146a and psoriasis: An association and functional study. J. Cell Mol. Med. 2014, 18, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Ramkaran, P.; Khan, S.; Phulukdaree, A.; Moodley, D.; Chuturgoon, A.A. miR-146a polymorphism influences levels of miR-146a, IRAK-1, and TRAF-6 in young patients with coronary artery disease. Cell Biochem. Biophys. 2014, 68, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Cusumano, A.; Falsini, B.; Giardina, E.; Cascella, R. Doyne honeycomb retinal dystrophy—Functional improvement following subthreshold nanopulse laser treatment: A case report. J. Med. Case Rep. 2019, 13, 5. [Google Scholar] [CrossRef]
- Grassmann, F.; Kiel, C.; Zimmermann, M.E.; Gorski, M.; Grassmann, V.; Stark, K.; Heid, I.M.; Weber, B.H. International AMD Genomics Consortium (IAMDGC) Genetic pleiotropy between age-related macular degeneration and 16 complex diseases and traits. Genome Med. 2017, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Dubey, R.; Saini, N. Cooperative and individualistic functions of the microRNAs in the miR-23a~27a~24-2 cluster and its implication in human diseases. Mol. Cancer 2010, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- Zhoua, Q.; Gallaghera, R.; Ufret-Vincentya, R.; Lia, X.; Olsonb, E.N.; Wanga, S. Regulation of angiogenesis and choroidal neovascularization by members of microRNA-23∼27∼24 clusters. Proc. Natl. Acad. Sci. USA 2011, 108, 8287–8292. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Jie, Z.; Ye, S.; Li, Z.; Han, Z.; Wu, J.; Yang, C.; Jiang, Y. Genetic variations in miR-27a gene decrease mature miR-27a level and reduce gastric cancer susceptibility. Oncogene 2014, 33, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Gu, H.; Zeng, Y.; Xia, Y.; Wang, Y.; Jing, Y.; Yang, L.; Wang, B. Hsa-mir-27a genetic variant contributes to gastric cancer susceptibility through affecting miR-27a and target gene expression. Cancer Sci. 2010, 101, 2241–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giri, M.; Shah, A.; Upreti, B.; Rai, J.C. Unraveling the genes implicated in Alzheimer’s disease. Biomed. Rep. 2017, 7, 105–114. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhattacharjee, S.; Jones, B.M.; Hill, J.M.; Clement, C.; Sambamurti, K.; Dua, P.; Lukiw, W.J. Beta-amyloid precursor protein (βAPP) processing in Alzheimer’s disease (AD) and age-related macular degeneration (AMD). Mol. Neurobiol. 2015, 52, 533–544. [Google Scholar] [CrossRef]
- Van der Heide, C.J.; Flamme-Wiese, M.; Khor, C.C.; Aung, T.; Rotter, J.I.; Weinreb, R.N.; Allingham, R.R.; Mullins, R.; Hauser, M.; Fingert, J.H.; The Eyes of Africa Consortium. Increased Retinal APBB2 and β-amyloid in Eyes with an APBB2 Risk Allele Associated with POAG. In Abstract Book of XXIII Biennial Meeting of the International Society for Eye Research; ISER: Northern Ireland, Belfast, UK, 2018. [Google Scholar]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.N. PPARγ agonists: Potential treatments for exudative age-related macular degeneration. Life Sci. 2017, 188, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Cascella, R.; Strafella, C.; Longo, G.; Manzo, L.; Ragazzo, M.; De Felici, C.; Gambardella, S.; Marsella, L.T.; Novelli, G.; Borgiani, P.; et al. Assessing individual risk for AMD with genetic counseling, family history, and genetic testing. Eye 2018, 32, 446–450. [Google Scholar] [CrossRef]
- Seddon, J.M.; Rosner, B. Validated prediction models for macular degeneration progression and predictors of visual acuity loss identify high-risk individuals. Am. J. Ophthalmol. 2019, 198, 223–261. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic. Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010, 11, R90. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Ziebarth, J.D.; Cui, Y. PolymiRTS Database 3.0: Linking polymorphisms in microRNAs and their target sites with human diseases and biological pathways. Nucleic Acids Res. 2014, 42, D86–D91. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Höner Zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithm. Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
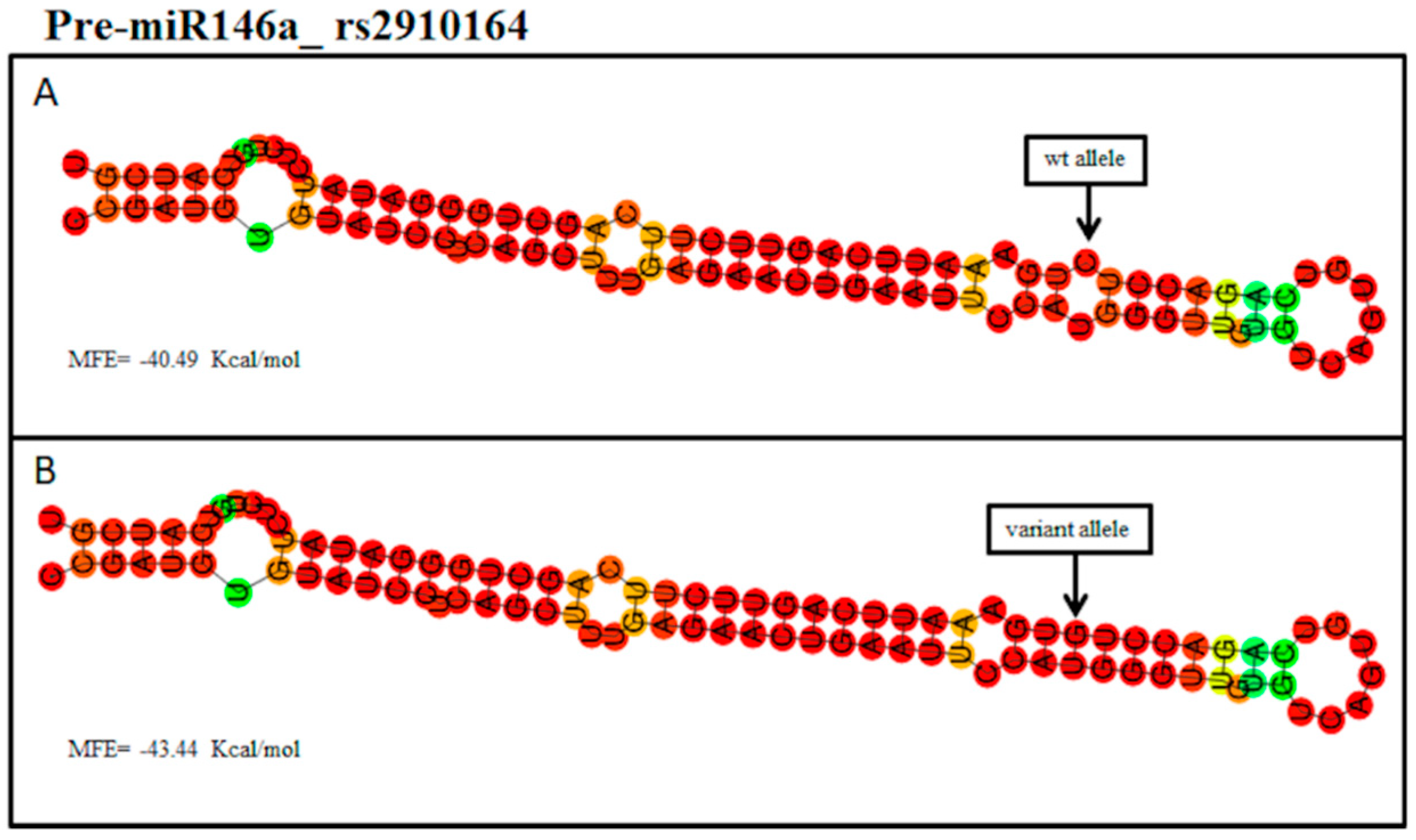
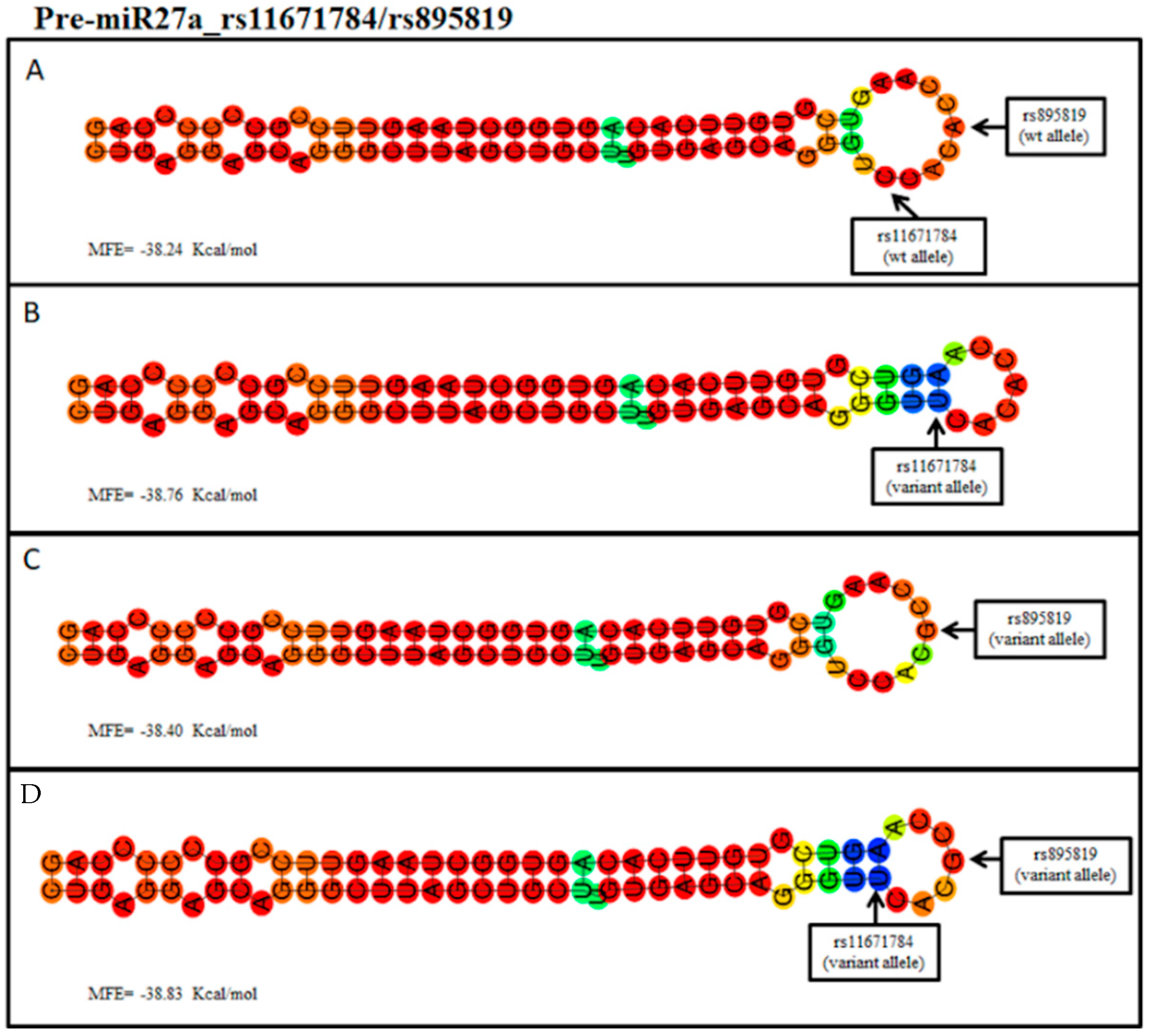
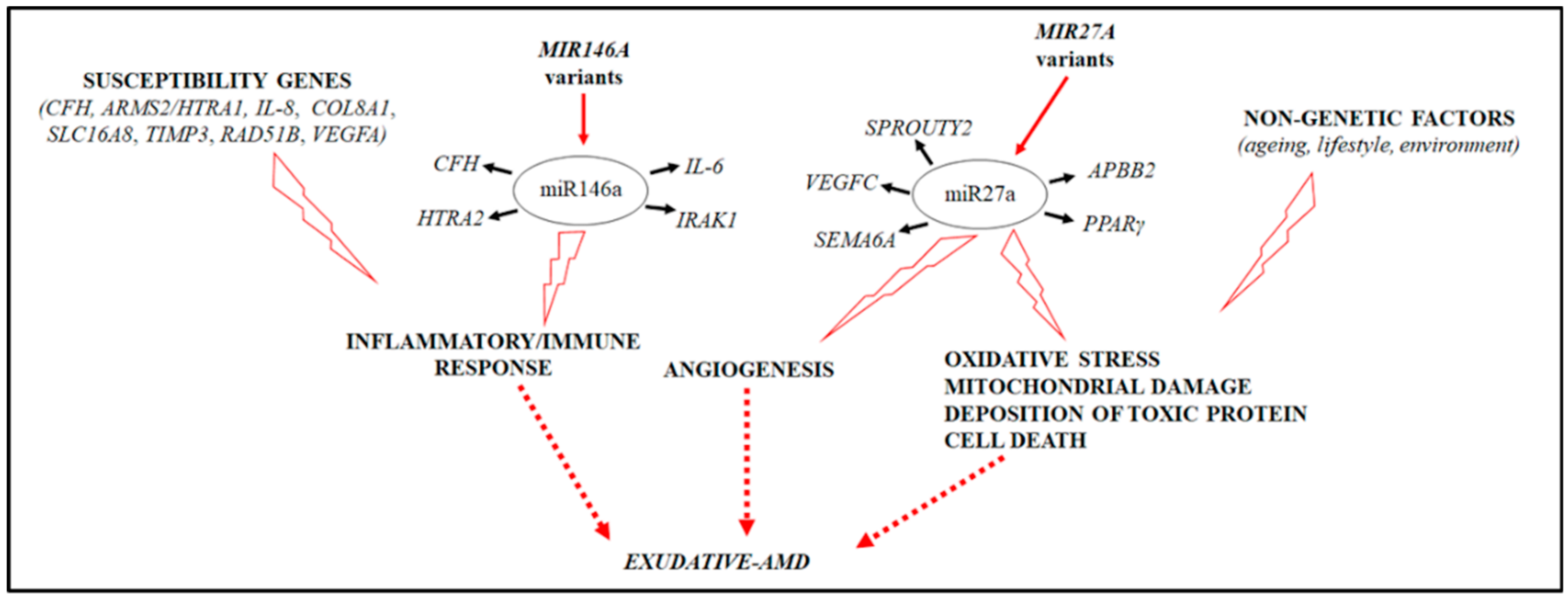

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzed Variants | Genotypes Number and Frequencies (%) in Cases | Genotypes Number and Frequencies (%) in Controls | Allelic p-Value | Allelic OR (CI:95%) | Impact |
|---|---|---|---|---|---|
| MIR146A rs2910164 (C/G) | C/C: 57 (7%) | C/C: 100 (10%) | 0.005 | Risk | |
| G/C: 303 (37%) | G/C: 388 (39%) | G = 1.23 (1.06–1.43) | |||
| G/G: 456 (56%) | G/G: 500 (50%) | ||||
| MIR31 rs772646842 (G/A) | G/G: 921 (99%) | G/G:977 (99%) | ns | - | |
| G/A: 10 (1%) | G/A: 10 (1%) | ||||
| A/A: 0 (0%) | A/A:0 (0%) | ||||
| MIR31 rs149347978 (C/T) | C/C: 921 (99%) | C/C:977 (99%) | ns | - | |
| C/T: 10 (1%) | C/T: 10 (1%) | ||||
| T/T: 0 (0%) | T/T:0 (0%) | ||||
| MIR31 rs192240130 (T/C) | T/T: 873 (99%) | T/T:922 (98%) | ns | - | |
| T/C: 9 (1%) | T/C: 18 (2%) | ||||
| C/C: 0 (0%) | C/C: 0 (0%) | ||||
| MIR23A rs771610178 (G/C) | G/G: 975 (99%) | G/G: 940 (99%) | ns | - | |
| G/C: 10 (1%) | G/C: 10 (1%) | ||||
| C/C: 0 (0%) | C/C: 0 (0%) | ||||
| MIR27A rs11671784 (G/A) | G/G: 742 (93%) | G/G: 768 (97%) | 0.001 | Risk | |
| G/A: 50 (6%) | G/A: 24 (3%) | A = 2.29 (1.41–3.72) | |||
| A/A: 2 (1%) | A/A: 0 (0%) | ||||
| MIR27A rs895819 (T/C) | T/T: 399 (52%) | T/T: 495 (57%) | 0.03 | Risk | |
| T/C: 301 (39%) | T/C: 315 (36%) | C = 1.19 (1.02–1.39) | |||
| C/C: 67 (9%) | C/C: 60 (7%) | ||||
| MIR20A (completely wild-type) | - | - | na | - | |
| MIR150 rs138052193 (-/AG) | 541 (62%) | 608 (67%) | na | - | |
| 323 (37%) | 272 (30%) | ||||
| 9 (1%) | 27 (3%) |
| Analyzed Variants | Genotype p-Value | Genotypic OR (CI:95%) | Dominant Model OR (CI:95%) | Recessive Model OR (CI:95%) |
|---|---|---|---|---|
| MIR146A rs2910164 (C/G) | 0.019 | GC = 1.38 (1.00–1.96) GG = 1.60 (1.12–2.26) | 1.50 (1.10–2.10) | 1.23 (1.02–1.48) |
| MIR27A rs11671784 (G/A) | 0.003 | GA = 2.15 (1.31–3.54) AA = na | 2.24 (1.36–3.67) | na |
| MIR27A rs895819 (T/C) | ns | - | - | - |
| Data | Cases | Controls |
|---|---|---|
| Age | ±77 years old | ±72 years old |
| Sex | F: 54% M: 46% | F: 56% M: 44% |
| Type of CNV | Type 1:53% Type 2: 47% | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strafella, C.; Errichiello, V.; Caputo, V.; Aloe, G.; Ricci, F.; Cusumano, A.; Novelli, G.; Giardina, E.; Cascella, R. The Interplay between miRNA-Related Variants and Age-Related Macular Degeneration: EVIDENCE of Association of MIR146A and MIR27A. Int. J. Mol. Sci. 2019, 20, 1578. https://doi.org/10.3390/ijms20071578
Strafella C, Errichiello V, Caputo V, Aloe G, Ricci F, Cusumano A, Novelli G, Giardina E, Cascella R. The Interplay between miRNA-Related Variants and Age-Related Macular Degeneration: EVIDENCE of Association of MIR146A and MIR27A. International Journal of Molecular Sciences. 2019; 20(7):1578. https://doi.org/10.3390/ijms20071578
Chicago/Turabian StyleStrafella, Claudia, Valeria Errichiello, Valerio Caputo, Gianluca Aloe, Federico Ricci, Andrea Cusumano, Giuseppe Novelli, Emiliano Giardina, and Raffaella Cascella. 2019. "The Interplay between miRNA-Related Variants and Age-Related Macular Degeneration: EVIDENCE of Association of MIR146A and MIR27A" International Journal of Molecular Sciences 20, no. 7: 1578. https://doi.org/10.3390/ijms20071578
APA StyleStrafella, C., Errichiello, V., Caputo, V., Aloe, G., Ricci, F., Cusumano, A., Novelli, G., Giardina, E., & Cascella, R. (2019). The Interplay between miRNA-Related Variants and Age-Related Macular Degeneration: EVIDENCE of Association of MIR146A and MIR27A. International Journal of Molecular Sciences, 20(7), 1578. https://doi.org/10.3390/ijms20071578