Human Vascular Pericytes and Cytomegalovirus Pathobiology

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
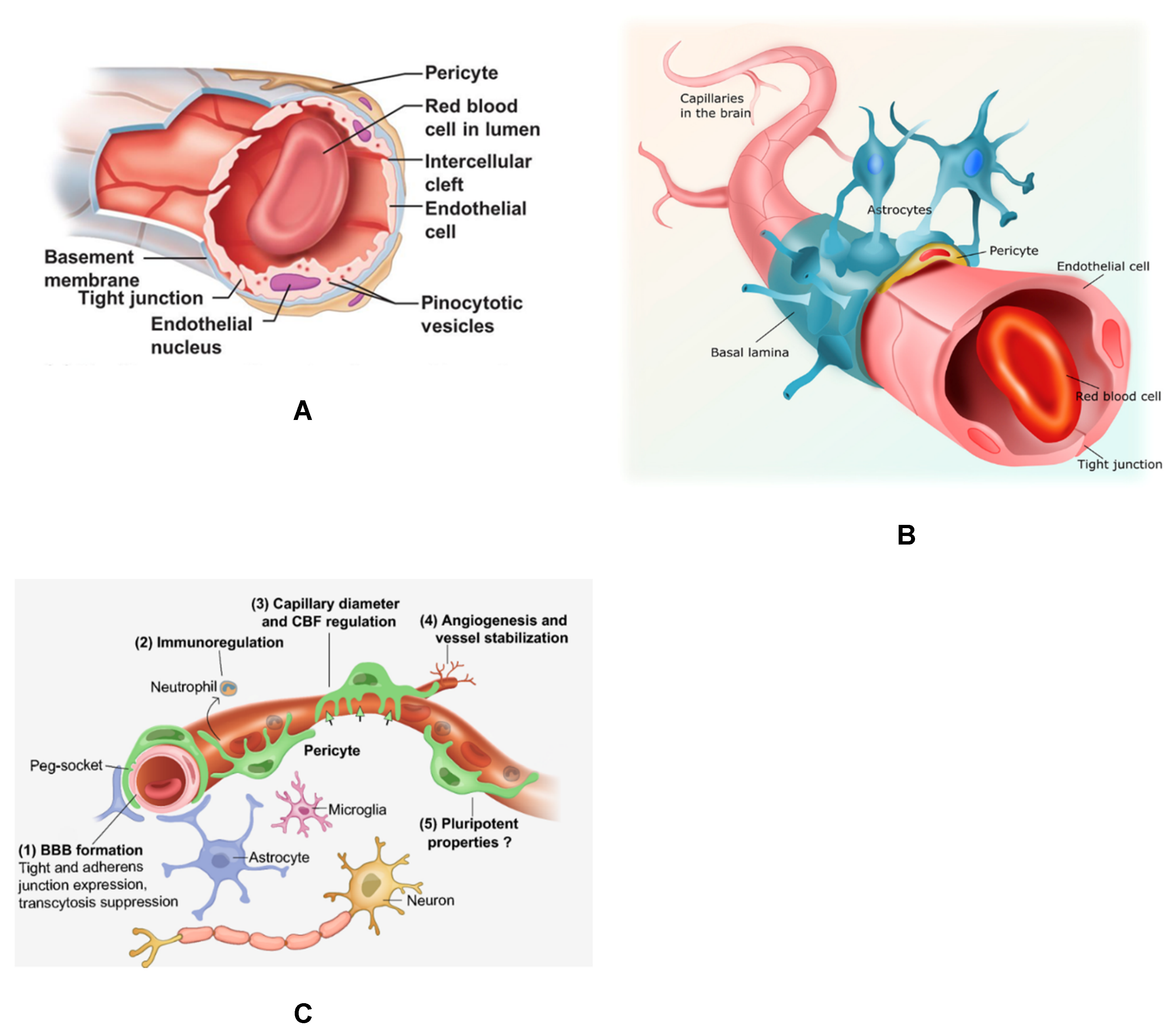
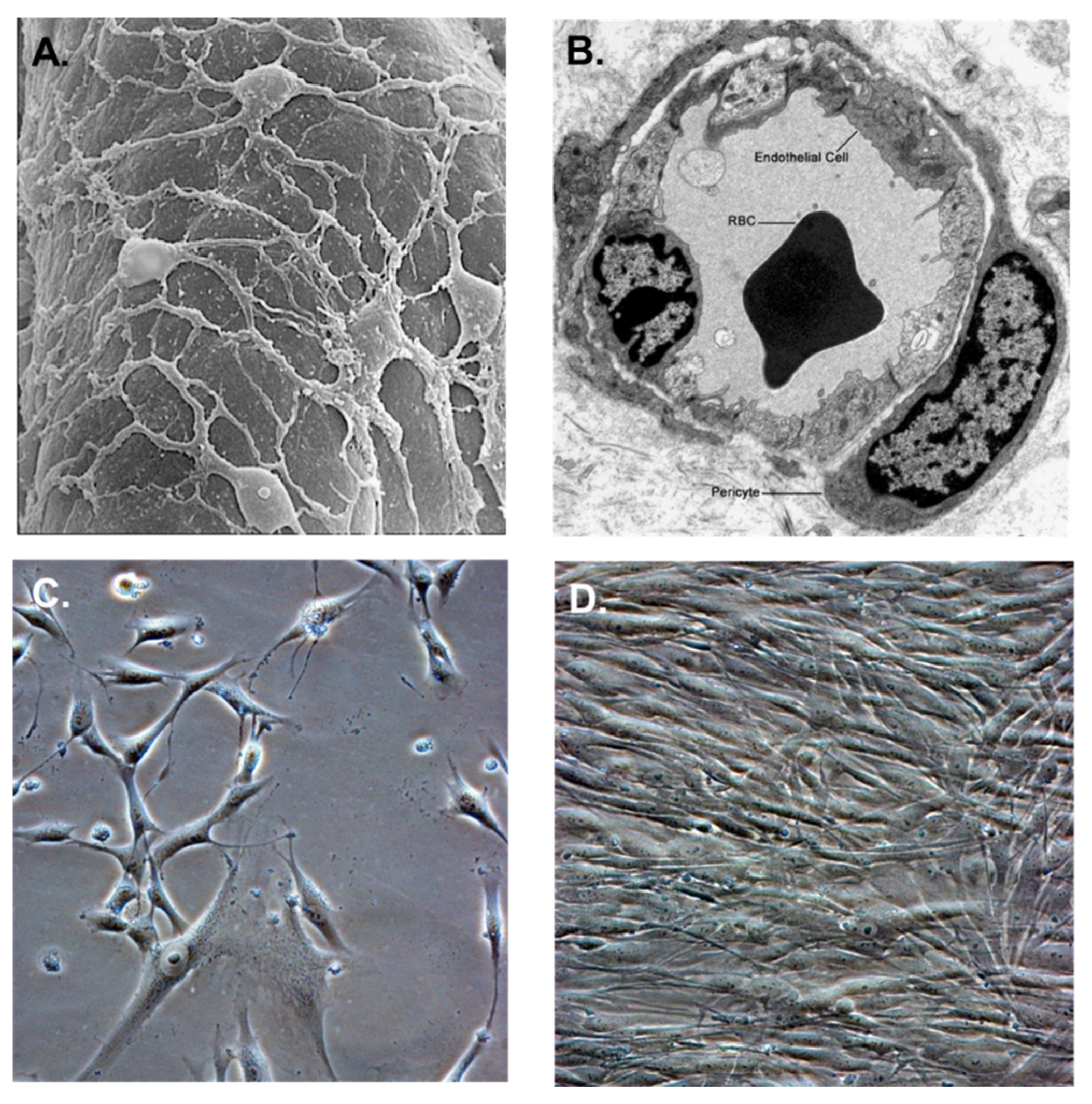
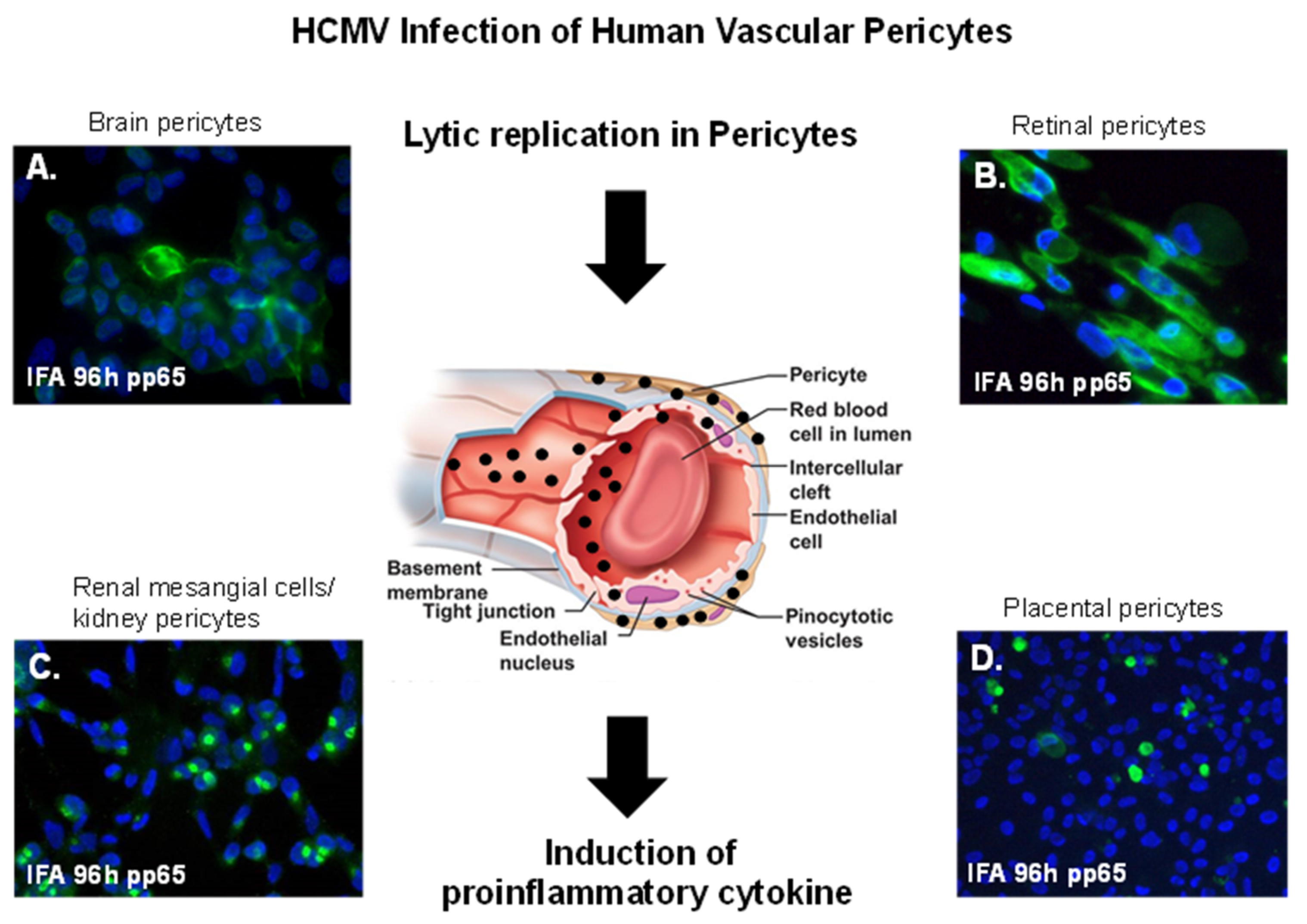
2. CMV Infection of Brain Vascular Pericytes: Implications for HCMV-Associated CNS Vascular Disease
3. Cytomegalovirus and Retinal Pericytes: Implications in Ocular Disease
4. Placental Pericytes and Cytomegalovirus Infection: Implication for CMV-Induced Congenital Diseases
5. Mesangial Cells (Specialized Renal Pericytes) and Cytomegalovirus Infectivity: Implications for Kidney Disease
6. HCMV Infection as A Risk Factor for Vascular Disease
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eberth, C.J. Handbuch der Lehre von der Gewegen des Menschen und der Tiere; Engelmann: Leipzig, Germany, 1871; Volume 1. [Google Scholar]
- Rouget, C. Me’moire sur le de´veloppement, la structure et les propriete´s physiologiques des capillaires sanguins et lymphatiques. Arch. Physiol. Norm Path. 1873, 5, 603–663. [Google Scholar]
- Zimmermann, K.W. Der feinere Bau der Blutkapillaren. Z. Anat. Entwicklungsgesch 1923, 68, 29–109. [Google Scholar] [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What is a pericyte? J. Cereb. Blood Flow Metab. 2016, 2, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.E. The pericyte—A review. Tissue Cell 1986, 18, 153–174. [Google Scholar] [CrossRef]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genove, G.; Betsholtz, C. Pericytes: Developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, N.B.; Attwell, D.; Hall, C.N. Pericyte-mediated regulation of capillary diameter: A component of neurovascular coupling in health and disease. Front. Neuroenerg. 2010, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Korn, C.; Augustin, H.G. Mechanisms of vessel pruning and regression. Dev. Cell 2015, 34, 5–17. [Google Scholar] [CrossRef]
- Alcendor, D.J.; Charest, A.M.; Zhu, W.Q.; Vigil, H.E.; Knobel, S.M. Infection and upregulation of proinflammatory cytokines in human brain vascular pericytes by human cytomegalovirus. J. Neuroinflamm. 2012, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, I.; Laban, J.; Mitchell, J.; Sheibani, N.; Alcendor, D.J. Retinal pericytes and cytomegalovirus infectivity: Implications for both CMV induced retinopathy and congenital ocular disease. J. Neuroinflamm. 2015, 12, 2. [Google Scholar] [CrossRef]
- Benjamin, L.E.; Hemo, I.; Keshet, E. A plasticity window for blood vessel remodelling is defined by pericyte coverage of the preformed endothelial network and is regulated by PDGF-B and VEGF. Development 1998, 125, 1591–1598. [Google Scholar]
- Enge, M.; Bjarnegård, M.; Gerhardt, H.; Gustafsson, E.; Kalén, M.; Asker, N.; Hammes, H.P.; Shani, M.; Fässler, R.; Betsholtz, C. Endothelium-specific platelet-derived growth factor-B ablation mimics diabetic retinopathy. EMBO J. 2002, 21, 4307–4316. [Google Scholar] [CrossRef]
- Müller, E.K.; Gräfe, C.; Wiekhorst, F.; Bergemann, C.; Weidner, A.; Dutz, S.; Clement, J.H. Magnetic Nanoparticles Interact and Pass an In Vitro Co-Culture Blood-Placenta Barrier Model. Nanomaterials 2018, 8, 108. [Google Scholar] [CrossRef]
- Aronoff, D.M.; Correa, H.; Rogers, L.M.; Arav-Boger, R.; Alcendor, D.J. A Placental pericytes and cytomegalovirus infectivity: Implications for HCMV placental pathology and congenital disease. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef]
- Stapor, P.C.; Sweat, R.S.; Dashti, D.C.; Betancourt, A.M.; Murfee, W.L. Pericyte dynamics during angiogenesis: New insights from new identities. J. Vasc. Res. 2014, 3, 163–174. [Google Scholar] [CrossRef]
- Challier, J.C.; Galtier, M.; Kacemi, A.; Guillaumin, D. Pericytes of term human foeto-placental microvessels: Ultrastructure and visualization. Cell Mol. Biol. 1999, 1, 89–100. [Google Scholar]
- Maier, C.L.; Pober, J.S. Human placental pericytes poorly stimulate and actively regulate allogeneic CD4 T cell responses. Arterioscler. Thromb. Vasc. Biol. 2011, 1, 183–189. [Google Scholar] [CrossRef]
- Popik, W.; Correa, H.; Khatua, A.; Aronoff, D.M.; Alcendor, D.J. Mesangial cells, specialized renal pericytes and cytomegalovirus infectivity: Implications for HCMV pathology in the glomerular vascular unit and post-transplant renal disease. J. Transl. Sci. 2019, 5. [Google Scholar] [CrossRef]
- Kurihara, H.; Sakai, T. Cell biology of mesangial cells: The third cell that maintains the glomerular capillary. Anat. Sci. Int. 2017, 2, 173–186. [Google Scholar] [CrossRef]
- Schlondorff, D. The glomerular mesangial cell: An expanding role for a specialized pericyte. FASEB J. 1987, 1, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N. Development of the glomerular mesangium. Pediatr. Nephrol. 1988, 2, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Flores, L.; Gutierrez, R.; Madrid, J.F.; Varela, H.; Valladares, F.; Acosta, E.; Martin-Vasallo, P.; Díaz-Flores, J.L. Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol. Histopathol. 2009, 24, 909–969. [Google Scholar] [PubMed]
- Lindahl, P.; Hellstrom, M.; Kalen, M.; Karlsson, L.; Pekny, M.; Pekna, M.; Soriano, P.; Betsholtz, C. Paracrine PDGF-B/PDGF-Rbeta signaling controls mesangial cell development in kidney glomeruli. Development 1998, 125, 3313–3322. [Google Scholar] [PubMed]
- Smith, S.W.; Chand, S.; Savage, C.O. Biology of the renal pericyte. Nephrol. Dial. Transplant. 2012, 27, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Balladh, P.; Braun, A.; Nedergaard, M. The blood-brain barrier: An overview, structure, regulation and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [CrossRef]
- Englehardt, B. Development of the blood-brain barrier. Cell Tissue Res. 2002, 314, 119–129. [Google Scholar]
- Garberg, P.; Ball, M.; Borg, N.; Cecchelli, R.; Fenart, L.; Hurst, R.D.; Lindmark, T.; Mabondzo, A.; Nilsson, J.E.; Raub, T.J.; et al. In vitro models for the blood–brain barrier. Toxicol. In Vitro 2005, 19, 299–334. [Google Scholar] [CrossRef]
- Ramsauer, M.; Krause, D.; Dermietzel, R. Angiogenesis of the blood-brain barrier in vitro and the function of cerebral pericytes. FASEB J. 2002, 16, 1274–1276. [Google Scholar] [CrossRef]
- Shepro, D.; Morel, N.M. Pericyte physiology. FASEB J. 1993, 7, 1031–1038. [Google Scholar] [CrossRef]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol. 2001, 2, 285–293. [Google Scholar] [CrossRef]
- Balabanov, R.; Dore-Duffy, P. Role of CNS microvascular pericytes in the blood brain barrier. J. Neurosci. Res. 1998, 53, 637–644. [Google Scholar] [CrossRef]
- Dore-Duffy, P. Pericytes: Pluripotent cells of the blood brain barrier. Curr. Pharm. Des. 2008, 16, 1581–1593. [Google Scholar] [CrossRef]
- Kelly-Goss, M.R.; Sweat, R.S.; Stapor, P.C.; Peirce, S.M.; Murfee, W.L. Targeting pericytes for angiogenic therapies. Microcirculation 2014, 4, 345–357. [Google Scholar] [CrossRef]
- Stark, K.; Pekayvaz, K.; Massberg, S. Role of pericytes in vascular immunosurveillance. Front. Biosci. 2018, 23, 767–781. [Google Scholar] [CrossRef]
- Lai, C.H.; Kuo, K.H. The critical component to establish in vitro BBB model: Pericyte. Brain Res. Rev. 2005, 50, 258–265. [Google Scholar] [CrossRef]
- Cheeran, M.C.J.; Lokensgard, J.R.; Schleiss, M.R. Neuropathogenesis of congenital cytomegalovirus infection: Disease mechanisms and prospects for intervention. Clin. Microbiol. Rev. 2009, 22, 99–126. [Google Scholar] [CrossRef]
- von Tell, D.; Armulik, A.; Betsholtz, C. Pericytes and vascular stability. Exp. Cell Res. 2006, 312, 623–629. [Google Scholar] [CrossRef]
- Park, D.Y.; Lee, J.; Kim, J.; Kim, K.; Hong, S.; Han, S.; Kubota, Y.; Augustin, H.G.; Ding, L.; Kim, J.W.; et al. Plastic roles of pericytes in the blood-retinal barrier. Nat. Commun. 2017, 8, 15296. [Google Scholar] [CrossRef]
- Dore-Duffy, P.; Cleary, K. Morphology and properties of pericytes. Methods Mol. Biol. 2011, 686, 49–68. [Google Scholar]
- Stagno, S.; Pass, R.F.; Cloud, G.; Britt, W.J.; Henderson, R.E.; Walton, P.D.; Veren, D.A.; Page, F.; Alford, C.A. Primary cytomegalovirus infection in pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA 1986, 256, 1904–1908. [Google Scholar] [CrossRef]
- Boppana, S.B.; Fowler, K.B.; Pass, R.F.; Rivera, L.B.; Bradford, R.D.; Lakeman, F.D.; Britt, W.J. Congenital cytomegalovirus infection: Association between virus burden in infancy and hearing loss. J. Pediatr. 2005, 146, 817–823. [Google Scholar] [CrossRef]
- Fowler, K.B.; Stagno, S.; Pass, R.F.; Britt, W.J.; Boll, T.J.; Alford, C.A. The outcome of congenital cytomegalovirus infection in relation to maternal antibody status. N. Engl. J. Med. 1992, 10, 663–667. [Google Scholar] [CrossRef]
- Boppana, S.B.; Fowler, K.B.; Vaid, Y.; Hedlund, G.; Stagno, S.; Britt, W.J.; Pass, R.F. Neuroradiographic Findings in the Newborn Period and Long-Term Outcome in Children with Symptomatic Congenital Cytomegalovirus Infection. Pediatrics 1997, 99, 409–414. [Google Scholar] [CrossRef]
- Fowler, K.; McCollister, F.P.; Dahle, A.J.; Boppana, S.; Britt, W.J.; Pass, R.F. Progressive and fluctuating sensorineural hearing loss in children with asymptomatic congenital cytomegalovirus infection. J. Pediatr. 1997, 130, 624–630. [Google Scholar] [CrossRef]
- Jabs, D.A. Acquired Immunodeficiency syndrome and the eye. Arch. Ophthalmol. 1996, 114, 863–866. [Google Scholar] [CrossRef]
- Iyer, J.V.; Connolly, J.; Agrawal, R.; Yeo, T.K.; Lee, B.; Au, B.; Teoh, S.C. Analysis of aqueous humor in HIV patients with cytomegalovirus retinitis. Cytokine 2013, 2, 541–547. [Google Scholar] [CrossRef]
- Cousins, S.W.; Espinosa-Heidmann, D.G.; Miller, D.M.; Pereira-Simon, S.; Hernandez, E.P.; Chien, H.; Meier-Jewett, C.; Dix, R.D. Macrophage activation associated with chronic murine cytomegalovirus infection results in more severe experimental choroidal neovascularization. PLoS Pathog. 2012, 4, e1002671. [Google Scholar] [CrossRef]
- Miller, D.M.; Espinosa-Heidmann, D.G.; Legra, J.; Dubovy, S.R.; Sũner, I.J.; Sedmak, D.D.; Dix, R.D.; Cousins, S.W. The association of prior cytomegalovirus infection with neovascular age-related macular degeneration. Am. J. Ophthalmol. 2004, 3, 323–328. [Google Scholar] [CrossRef]
- Yinon, Y.; Farine, D.; Yudin, M.H.; Maternal Fetal Medicine Committee; Infectious Diseases Committee. Cytomegalovirus infection in pregnancy. J. Obstet. Gynaecol. Can. 2010, 4, 348–354. [Google Scholar] [CrossRef]
- Coll, O.; Benoist, G.; Ville, Y.; Weisman, L.E.; Botet, F.; Anceschi, M.M.; Greenough, A.; Gibbs, R.S.; Carbonell-Estrany, X.; WAPM Perinatal Infections Working Group. Guidelines on CMV congenital infection. J. Perinat. Med. 2009, 5, 433–445. [Google Scholar]
- Nigro, G.; Adler, S.P. Cytomegalovirus infections during pregnancy. Curr. Opin. Obstet. Gynecol. 2011, 23, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Boppana, S.B.; Pass, R.F.; Britt, W.J.; Stagno, S.; Alford, C.A. Symptomatic congenital cytomegalovirus infection: Neonatal morbidity and mortality. Pediatr. Infect. Dis. J. 1992, 11, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Pass, R.F.; Anderson, B. Mother-to-child transmission of cytomegalovirus and prevention of congenital infection. J. Pediatr. Infect. Dis. Soc. 2014, 3, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Bialek, S.; Cannon, M.J. Attribution of congenital cytomegalovirus infection to primary versus non-primary maternal infection. Clin. Infect. Dis. 2011, 2, e11–e13. [Google Scholar] [CrossRef]
- Kylat, R.I.; Kelly, E.N.; Ford-Jones, E.L. Clinical findings and adverse outcomes in neonates with symptomatic congenital cytomegalovirus (SCCMV) infection. Eur. J. Pediatr. 2006, 165, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Lichtner, M.; Cicconi, P.; Vita, S.; Cozzi-Lepri, A.; Galli, M.; Lo Caputo, S.; Saracino, A.; De Luca, A.; Moioli, M.; Maggiolo, F.; et al. Cytomegalovirus Coinfection is Associated with an Increased Risk of Severe Non-AIDS-Defining Events in a Large Cohort of HIV-Infected Patients. J. Infect. Dis. 2015, 2, 178–186. [Google Scholar] [CrossRef]
- Beam, E.; Dioverti, V.; Razonable, R.R. Emerging cytomegalovirus management strategies after solid organ transplantation: Challenges and opportunities. Curr. Infect. Dis. Rep. 2014, 9, 419. [Google Scholar] [CrossRef]
- Colugnati, F.A.; Staras, S.A.; Dollard, S.C.; Cannon, M.J. Incidence of cytomegalovirus infection among the general population and pregnant women in the United States. BMC Infect. Dis. 2007, 7, 71. [Google Scholar] [CrossRef]
- Pereira, L.; Petitt, M.; Fong, A.; Tsuge, M.; Tabata, T.; Fang-Hoover, J.; Maidji, E.; Zydek, M.; Zhou, Y.; Inoue, N.; et al. Intrauterine growth restriction caused by underlying congenital cytomegalovirus infection. J. Infect. Dis. 2014, 10, 1573–1584. [Google Scholar] [CrossRef]
- Birbrair, A. Pericyte Biology: Development, Homeostasis, and Disease. Adv. Exp. Med. Biol. 2018, 1109, 1–3. [Google Scholar] [PubMed]
- Jones, C.J.; Desoye, G. A new possible function for placental pericytes. Cells Tissue Organs 2011, 1, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.T.; Scott, G.; Naing, Z.; Iwasenko, J.; Hall, B.; Graf, N.; Arbuckle, S.; Craig, M.E.; Rawlinson, W.D. Human cytomegalovirus-induces cytokine changes in the placenta with implications for adverse pregnancy outcomes. PLoS ONE 2012, 7, e52899. [Google Scholar] [CrossRef]
- Ahmad, S.; Ahmed, A. Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ. Res. 2004, 9, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Gealy, C.; Denson, M.; Humphreys, C.; McSharry, B.; Wilkinson, G.; Caswell, R. Posttranscriptional suppression of interleukin-6 production by human cytomegalovirus. J. Virol. 2005, 79, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R.H. Infection in the renal and liver transplant patient. In Clinical Approach to Infections in the Compromised Host, 2nd ed.; Plenum Press: New York, NY, USA, 1988. [Google Scholar]
- Kotton, C.N.; Fishman, J.A. Viral infection in the renal transplant recipient. J. Am. Soc. Nephrol. 2005, 16, 1758–1774. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.R.; Butterly, D.W.; Alexander, B.D.; Greenberg, A. Viral infections after renal transplantation. Am. J. Kidney Dis. 2001, 37, 659–676. [Google Scholar] [CrossRef]
- Brennan, D.C. Cytomegalovirus in renal transplantation. J. Am. Soc. Nephrol. 2001, 12, 848–855. [Google Scholar]
- Erdbrügger, U.; Scheffner, I.; Mengel, M.; Schwarz, A.; Haller, H.; Gwinner, W. Long-term impact of CMV infection on allografts and on patient survival in renal transplant patients with protocol biopsies. Am. J. Physiol. Renal Physiol. 2015, 11, F925–F932. [Google Scholar] [CrossRef]
- Sagedal, S.; Nordal, K.P.; Hartmann, A.; Sund, S.; Scott, H.; Degré, M.; Foss, A.; Leivestad, T.; Osnes, K.; Fauchald, P.; et al. The impact of cytomegalovirus infection and disease on rejection episodes in renal allograft recipients. Am. J. Transplant. 2002, 9, 850–885. [Google Scholar] [CrossRef]
- Sagedal, S.; Hartmann, A.; Nordal, K.P.; Osnes, K.; Leivestad, T.; Foss, A.; Degré, M.; Fauchald, P.; Rollag, H. Impact of early cytomegalovirus infection and disease on long-term recipient and kidney graft survival. Kidney Int. 2004, 1, 329–337. [Google Scholar] [CrossRef] [PubMed]
- De Keyzer, K.; Van Laecke, S.; Peeters, P.; Vanholder, R. Human cytomegalovirus and kidney transplantation: A clinician’s update. Am. J. Kidney Dis. 2011, 58, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Chiasakul, T.; Townamchai, N.; Jutivorakool, K.; Chancharoenthana, W.; Thongprayoon, C.; Watanatorn, S.; Avihingsanon, Y.; Praditpornsilpa, K.; Srisawat, N. Risk factors of cytomegalovirus disease in kidney transplant recipients: A single-center study in Thailand. Transplant. Proc. 2015, 8, 2460–2464. [Google Scholar] [CrossRef]
- Yalci, A.; Celebi, Z.K.; Ozbas, B.; Sengezer, O.L.; Unal, H.; Memikoğlu, K.O.; Sengul, S.; Tuzuner, A.; Keven, K. Evaluation of infectious complications in the first year after kidney transplantation. Transplant. Proc. 2015, 5, 1429–1432. [Google Scholar] [CrossRef] [PubMed]
- Bataille, S.; Moal, V.; Gaudart, J.; Indreies, M.; Purgus, R.; Dussol, B.; Zandotti, C.; Berland, Y.; Vacher-Coponat, H. Cytomegalovirus risk factors in renal transplantation with modern immunosuppression. Transpl. Infect. Dis. 2010, 6, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Besbas, N.; Bayrakci, U.S.; Kale, G.; Cengiz, A.B.; Akcoren, Z.; Akinci, D.; Kilic, I.; Bakkaloglu, A. Cytomegalovirus related congenital nephrotic syndrome with diffuse mesangial sclerosis. Pediatr. Nephrol. 2006, 5, 740–742. [Google Scholar] [CrossRef] [PubMed]
- Ortmanns, A.; Ittel, T.H.; Schnitzler, N.; Handt, S.; Helmchen, U.; Sieberth, G. Remission of IgA nephropathy following treatment of cytomegalovirus infection with ganciclovir. Clin. Nephrol. 1998, 6, 379–384. [Google Scholar]
- Ustinov, J.A.; Loginov, R.J.; Mattila, P.M.; Nieminen, V.K.; Suni, J.I.; Häyry, P.J.; Lautenschlager, I.T. Cytomegalovirus infection of human kidney cells in vitro. Kidney Int. 1991, 40, 954–960. [Google Scholar] [CrossRef]
- Ustinov, J.; Mattila, P.; Häyry, P.; Lautenschlager, I. CMV infection in various cell types of human kidney. Transplant. Proc. 1992, 24, 285. [Google Scholar]
- Heieren, M.H.; van der Woude, F.J.; Balfour, H.H. Cytomegalovirus replicates efficiently in human kidney mesangial cells. Proc. Natl. Acad. Sci. USA 1988, 85, 1642–1646. [Google Scholar] [CrossRef]
- Ho, M. (Ed.) Cytomegalovirus: Biology and Infection, 2nd ed.; Plenum Medical Book Company: New York, NY, USA, 1991; pp. 75–99. [Google Scholar]
- Humar, A.; St Louis, P.; Mazzulli, T.; McGeer, A.; Lipton, J.; Messner, H.; MacDonald, K.S. Elevated serum cytokines are associated with cytomegalovirus infection and disease in bone marrow transplant recipients. J. Infect. Dis. 1999, 2, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Varani, S.; Landini, M.P. Cytomegalovirus-induced immunopathology and its clinical consequences. Herpesviridae 2011, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.J.; Liu, J.; Han, F.F.; Wan, Z.R.; Gong, L.L.; Liu, H.; Zhang, W.; Wardell, T.; Lv, Y.L.; Liu, L.H. Cytomegalovirus infection and atherosclerosis risk: A meta-analysis. J. Med. Virol. 2017, 12, 2196–2206. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.L.; Han, F.F.; Gong, L.L.; Liu, H.; Ma, J.; Yu, W.Y.; Wan, Z.R.; Jia, Y.J.; Zhang, W.; Shi, M.; et al. Human cytomegalovirus infection and vascular disease risk: A meta-analysis. Virus Res. 2017, 227, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.R.; Yu, L.P.; Yang, X.C.; Zhang, F.; Chen, Y.R.; Feng, F.; Qian, X.S.; Cai, J. Human cytomegalovirus linked to stroke in a Chinese population. CNS Neurosci. Ther. 2012, 6, 457–460. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcendor, D.J. Human Vascular Pericytes and Cytomegalovirus Pathobiology. Int. J. Mol. Sci. 2019, 20, 1456. https://doi.org/10.3390/ijms20061456
Alcendor DJ. Human Vascular Pericytes and Cytomegalovirus Pathobiology. International Journal of Molecular Sciences. 2019; 20(6):1456. https://doi.org/10.3390/ijms20061456
Chicago/Turabian StyleAlcendor, Donald J. 2019. "Human Vascular Pericytes and Cytomegalovirus Pathobiology" International Journal of Molecular Sciences 20, no. 6: 1456. https://doi.org/10.3390/ijms20061456
APA StyleAlcendor, D. J. (2019). Human Vascular Pericytes and Cytomegalovirus Pathobiology. International Journal of Molecular Sciences, 20(6), 1456. https://doi.org/10.3390/ijms20061456