Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures

 ,
,  ,
,
Abstract
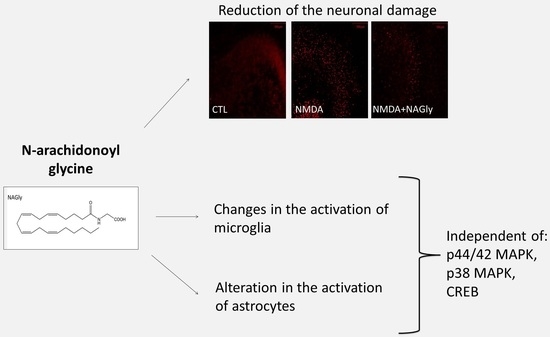
1. Introduction
2. Results
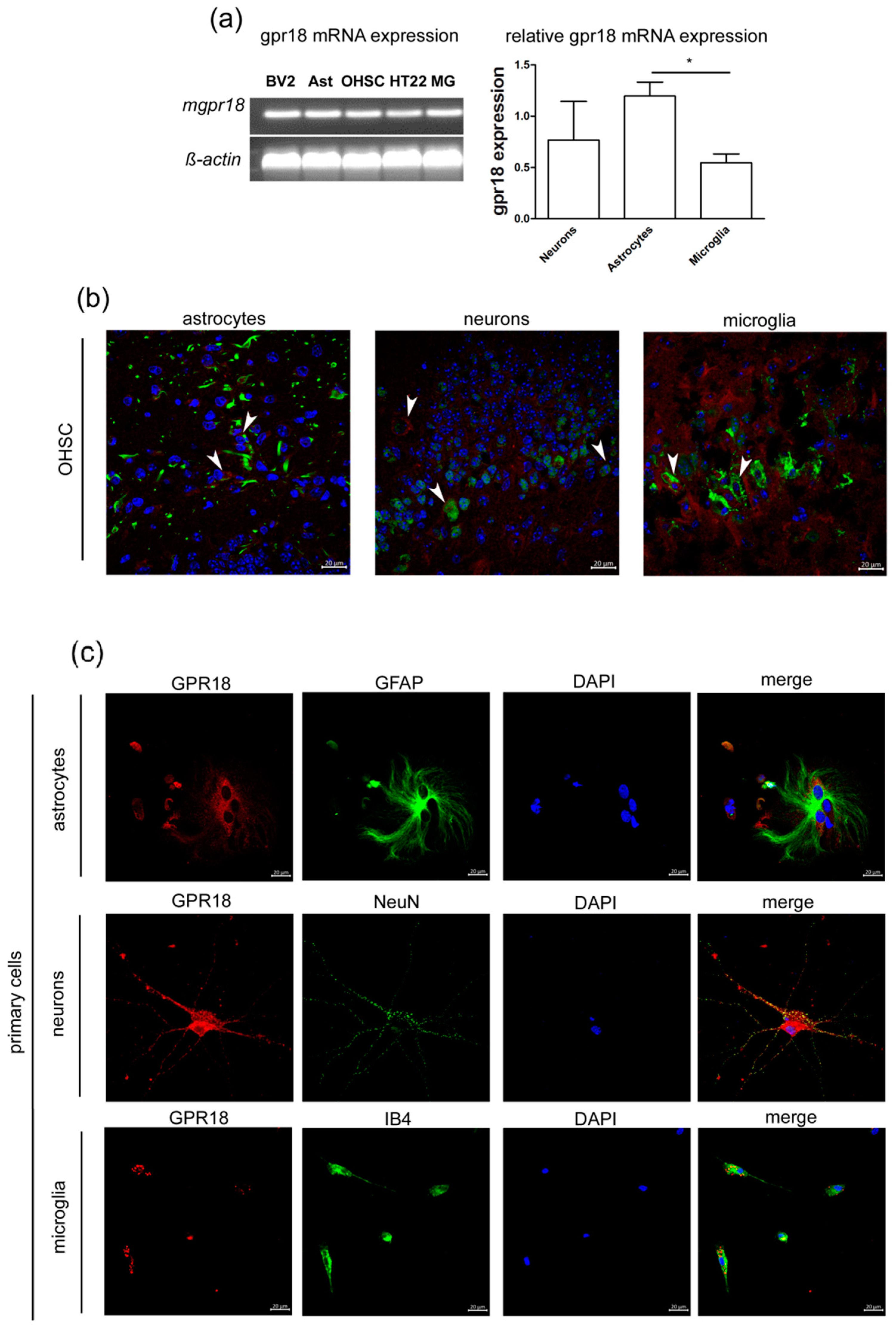
2.1. GPR18 mRNA and Protein are Expressed in Primary Astrocytes, Microglia and Neurons
2.2. Slight Changes in gpr18 mRNA and No Changes in GPR18 Protein Expression in OHSC
2.3. NAGly is Neuroprotective in NMDA Lesioned OHSC
2.4. NAGly Mediated Neuroprotection is Possibly Mediated by GPR18 and CB2 Receptors in OHSC
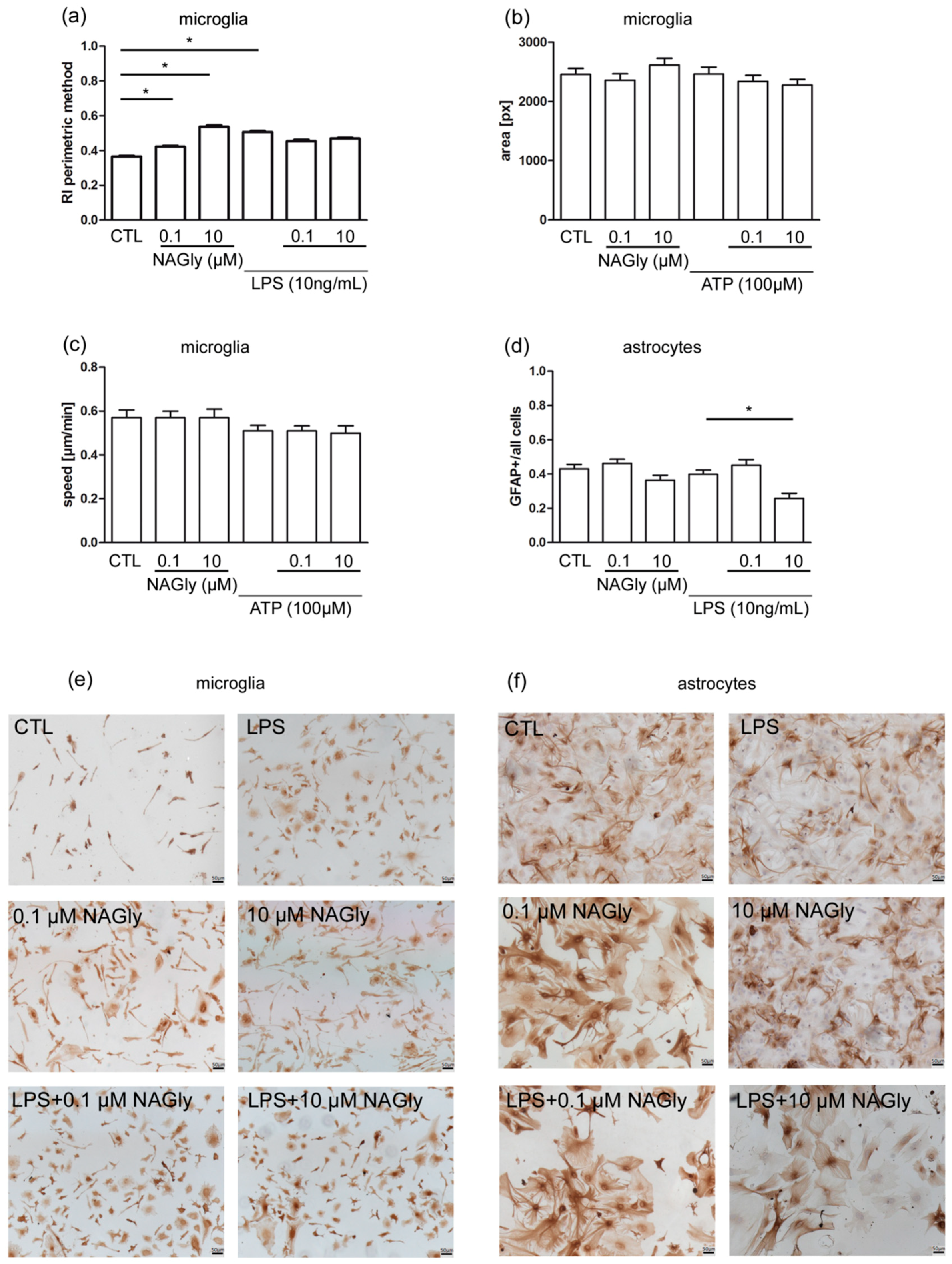
2.5. Ramification Index of Isolated Primary Microglia
2.6. NAGly Did Not Induce Changes in the Motility of Isolated Wild Type Microglia
2.7. Activation of Cultured Astrocytes Measured as GFAP Index
2.8. Activation of 44/42 MAPK and pCREB is Not Altered, Whereas pAkt is Downregulated Only at 30 min in NAGly Treated BV2 Cells
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Chemicals
4.3. Organotypic Hippocampal Slice Cultures (OHSC)
4.4. Primary Cell Cultures and Cell Lines
4.5. qRT-PCR
4.6. Western Blot Analysis
4.7. Immunohistochemistry
4.8. Propidium Iodide Staining and Confocal Laser Scanning Microscopy
4.9. Analysis of Cell Motility
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CB1 | cannabinoid receptor 1 |
| CB2 | cannabinoid receptor 2 |
| GFAP | glial fibrillary acidic protein |
| GPR18 | G protein-coupled receptor 18 |
| GPR55 | G protein-coupled receptor 55 |
| LPS | lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| NAGly | N-arachidonoyl glycine |
| NMDA | N-methyl-D-aspartate receptor |
| OHSC | organotypic hippocampal slice cultures |
| SEM | standard error of the mean |
| SD | standard deviation |
| THC | Δ9-tetrahydrocannabinol |
Appendix A


References
- Huang, S.M.; Bisogno, T.; Petros, T.J.; Chang, S.Y.; Zavitsanos, P.A.; Zipkin, R.E.; Sivakumar, R.; Coop, A.; Maeda, D.Y.; De Petrocellis, L.; et al. Identification of a New Class of Molecules, the Arachidonyl Amino Acids, and Characterization of One Member That Inhibits Pain. J. Biol. Chem. 2001, 276, 42639–42644. [Google Scholar] [CrossRef] [PubMed]
- Parmar, N.; Ho, W.S.V. N-arachidonoyl glycine, an endogenous lipid that acts as a vasorelaxant via nitric oxide and large conductance calcium-activated potassium channels. Br. J. Pharmacol. 2010, 160, 594–603. [Google Scholar] [CrossRef]
- Sheskin, T.; Hanuš, L.; Slager, J.; Vogel, Z.; Mechoulam, R. Structural requirements for binding of anandamide-type compounds to the brain cannabinoid receptor. J. Med. Chem. 1997, 40, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Malfitano, A.M.; Basu, S.; Maresz, K.; Bifulco, M.; Dittel, B.N. What We Know and Don’t Know About the Cannabinoid Receptor 2 (CB2). Semin. Immunol. 2014, 26, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Takenouchi, R.; Inoue, K.; Kambe, Y.; Miyata, A. N-arachidonoyl glycine induces macrophage apoptosis via GPR18. Biochem. Biophys. Res. Commun. 2012, 418, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Gantz, I.; Muraoka, A.; Yang, Y.; Samuelson, L.C.; Zimmerman, E.M.; Cook, H.; Yamada, T. Cloning and Chromosomal Localization of a Gene (GPR18) Encoding a Novel Seven Transmembrane Receptor Highly Expressed in Spleen and Testis. Genomics 1997, 466, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Hasegawa, H.; Inoue, A.; Muraoka, M.; Miyazaki, T.; Oka, K.; Yasukawa, M. Identification of N-arachidonylglycine as the endogenous ligand for orphan G-protein-coupled receptor GPR18. Biochem. Biophys. Res. Commun. 2006, 347, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Finlay, D.B.; Joseph, W.R.; Grimsey, N.L.; Glass, M. GPR18 undergoes a high degree of constitutive trafficking but is unresponsive to N-Arachidonoyl Glycine. PeerJ 2016, 4, e1835. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Hu, S.S.J.; Rimmerman, N.; Juknat, A.; Vogel, Z.; Walker, J.M.; Bradshaw, H.B. N-arachidonoyl glycine, an abundant endogenous lipid, potently drives directed cellular migration through GPR18, the putative abnormal cannabidiol receptor. BMC Neurosci. 2010, 11, 44. [Google Scholar] [CrossRef]
- Succar, R.; Mitchell, V.A.; Vaughan, C.W. Actions of N-arachidonyl-glycine in a rat inflammatory pain model. Mol. Pain 2007, 3, 24. [Google Scholar] [CrossRef]
- Burstein, S.H.; Mcquain, C.A.; Ross, A.H.; Salmonsen, R.A.; Zurier, R.E. Resolution of Inflammation by N-Arachidonoylglycine Sumner. Cell. Biochem. 2011, 113, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Wager-Miller, J.; Page, J.; Bradshaw, H.B. siRNA knockdown of GPR18 receptors in BV-2 microglia attenuates N-arachidonoyl glycine-induced cell migration. J. Mol. Signal. 2012, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.B.; Puhl, H.L.; Ikeda, S.R. N-Arachidonyl glycine does not activate G protein-coupled receptor 18 signaling via canonical pathways. Mol. Pharmacol. 2013, 83, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chu, A.; Li, W.; Wang, B.; Shelton, F.; Otero, F.; Nguyen, D.G.; Caldwell, J.S.; Chen, Y.A. Lipid G protein-coupled receptor ligand identification using beta-arrestin PathHunterTM assay. J. Biol. Chem. 2009, 284, 12328–12338. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, F.; Koch, M.; Pieroh, P.; Ghadban, C.; Hobusch, C.; Bechmann, I.; Dehghani, F. Time dependent neuroprotection of mycophenolate mofetil: Effects on temporal dynamics in glial proliferation, apoptosis, and scar formation. J. Neuroinflamm. 2012, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Gähwiler, B.H.; Capogna, M.; Debanne, D.; McKinney, R.A.; Thompson, S.M. Organotypic slice sultures: A technique has come of age. Trends Neurosci. 1997, 20, 471–477. [Google Scholar] [CrossRef]
- Grabiec, U.; Koch, M.; Kallendrusch, S.; Kraft, R.; Hill, K.; Merkwitz, C.; Ghadban, C.; Lutz, B.; Straiker, A.; Dehghani, F. The endocannabinoid N-arachidonoyldopamine (NADA) exerts neuroprotective effects after excitotoxic neuronal damage via cannabinoid receptor 1 (CB 1). Neuropharmacology 2012, 62, 1797–1807. [Google Scholar] [CrossRef]
- Grabiec, U.; Hohmann, T.; Hammer, N.; Dehghani, F. Organotypic Hippocampal Slice Cultures As a Model to Study Neuroprotection and Invasiveness of Tumor Cells. J. Vis. Exp. 2017, 126, e55359. [Google Scholar] [CrossRef]
- Langlois, J.A.; Rutland-Brown, W.; Wald, M.M. The Epidemiology and Impact of Traumatic Brain Injury A Brief Overview. J. Head Trauma Rehabil. 2006, 21, 375–378. [Google Scholar] [CrossRef]
- Koch, M.; Kreutz, S.; Boettger, C.; Benz, A.; Maronde, E.; Ghadban, C.; Korf, H.-W.; Dehghani, F. Palmitoylethanolamide Protects Dentate Gyrus Granule Cells via Peroxisome Proliferator-Activated Receptor-Alpha. Neurotox. Res. 2011, 19, 330–340. [Google Scholar] [CrossRef]
- Kreutz, S.; Koch, M.; Böttger, C.; Ghadban, C.; Korf, H.W.; Dehghani, F. 2-Arachidonoylglycerol Elicits Neuroprotective Effects on Excitotoxically Lesioned Dentate Gyrus Granule Cells Via Abnormal-Cannabidiol-Sensitive Receptors on Microglial Cells. Glia 2009, 57, 286–294. [Google Scholar] [CrossRef]
- Kreutz, S.; Koch, M.; Ghadban, C.; Korf, H.; Dehghani, F. Cannabinoids and neuronal damage: Differential effects of THC, AEA and 2-AG on activated microglial cells and degenerating neurons in excitotoxically lesioned rat organotypic hippocampal slice cultures. Exp. Neurol. 2007, 203, 246–257. [Google Scholar] [CrossRef]
- Shohami, E.; Cohen-Yeshurun, A.; Magid, L.; Algali, M.; Mechoulam, R. Endocannabinoids and traumatic brain injury. Br. J. Pharmacol. 2011, 163, 1402–1410. [Google Scholar] [CrossRef]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ 9-tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef]
- Fumagalli, S.; Perego, C.; Pischiutta, F.; Zanier, E.R.; De Simoni, M.-G. The ischemic environment drives microglia and macrophage function. Front. Neurol. 2015, 6, 81. [Google Scholar] [CrossRef]
- Kohl, A.; Dehghani, F.; Korf, H.; Hailer, N.P. The bisphosphonate clodronate depletes microglial cells in excitotoxically injured organotypic hippocampal slice cultures. Exp. Neurol. 2003, 181, 1–11. [Google Scholar] [CrossRef]
- Kunz, A.; Dirnagl, U.; Mergenthaler, P. Acute pathophysiological processes after ischaemic and traumatic brain injury. Best Pract. Res. Clin. Anaesthesiol. 2010, 24, 495–509. [Google Scholar] [CrossRef]
- Szalay, G.; Martinecz, B.; Lénárt, N.; Környei, Z.; Orsolits, B.; Judák, L.; Császár, E.; Fekete, R.; West, B.L.; Katona, G.; et al. Microglia protect against brain injury and their selective elimination dysregulates neuronal network activity after stroke. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Xiong, X.-Y.; Liu, L.; Yang, Q.-W. Functions and Mechanisms of Microglia/Macrophages in Neuroinflammation and Neurogenesis during Stroke. Prog. Neurobiol. 2016, 142, 23–44. [Google Scholar] [CrossRef]
- Vassilatis, D.K.; Hohmann, J.G.; Zeng, H.; Li, F.; Ranchalis, J.E.; Mortrud, M.T.; Brown, A.; Rodriguez, S.S.; Weller, J.R.; Wright, A.C.; et al. The G protein-coupled receptor repertoires of human and mouse. Proc. Natl. Acad. Sci. USA 2003, 100, 4903–4908. [Google Scholar] [CrossRef]
- Adamchik, Y.; Frantseva, M.V.; Weisspapir, M.; Carlen, P.L.; Velazquez, J.L.P. Methods to induce primary and secondary traumatic damage in organotypic hippocampal slice cultures. Brain Res. Protoc. 2000, 5, 153–158. [Google Scholar] [CrossRef]
- Koch, M.; Kreutz, S.; Böttger, C.; Grabiec, U.; Ghadban, C.; Korf, H.W.; Dehghani, F. The cannabinoid WIN 55,212-2-mediated protection of dentate gyrus granule cells is driven by CB1 receptors and modulated by TRPA1 and Ca v2.2 channels. Hippocampus 2011, 21, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Shohami, E. Endocannabinoids and traumatic brain injury. Mol. Neurobiol. 2007, 36, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Panikashvili, D.; Simeonidou, C.; Ben-shabat, S.; Breuer, A.; Mechoulam, R.; Shohami, E. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature 2001, 413, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Kallendrusch, S.; Hobusch, C.; Ehrlich, A.; Ziebell, S.; Ueda, N.; Geisslinger, G.; Koch, M.; Dehghani, F. Site-specific and time-dependent activation of the endocannabinoid system after transection of long-range projections. PLoS ONE 2012, 7, e33537. [Google Scholar] [CrossRef] [PubMed]
- Sipe, J.C.; Arbour, N.; Gerber, A.; Beutler, E. Reduced endocannabinoid immune modulation by a common cannabinoid 2 (CB2) receptor gene polymorphism: Possible risk for autoimmune disorders. J. Leukoc. Biol. 2005, 78, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Resina, I.; Navarro, G.; Aguinaga, D.; Canela, E.I.; Schoeder, C.T.; Kie, K.; Saura, C.A.; Müller, C.E.; Franco, R. Molecular and functional interaction between GPR18 and cannabinoid CB 2 G-protein-coupled receptors. Relevance in neurodegenerative diseases. Biochem. Pharmacol. 2018. [Google Scholar] [CrossRef]
- Franklin, A.; Stella, N. Arachidonylcyclopropylamide increases microglial cell migration through cannabinoid CB 2 and abnormal-cannabidiol-sensitive receptors. Eur. J. Pharmacol. 2003, 474, 195–198. [Google Scholar] [CrossRef]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef]
- Qin, L.; Li, G.; Qian, X.; Liu, Y.; Wu, X.; Liu, B.; Hong, J.S.; Block, M.L. Interactive role of the toll-like receptor 4 and reactive oxygen species in LPS-induced microglia activation. Glia 2005, 52, 78–84. [Google Scholar] [CrossRef]
- Hohmann, T.; Grabiec, U.; Ghadban, C.; Feese, K.; Dehghani, F. The Influence of Biomechanical Properties and Cannabinoids on Tumor Invasion. Cell Adh. Migr. 2017, 11, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef]
- Schwartzfarb, L.; Needle, M.; Chavez-Chase, M. Dose-related inhibition of leukocyte migration by marihuana and delta-9-tetrahydrocannabinol (THC) in vitro. J. Clin. Pharmacol. 1974, 14, 35–41. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E.M. Progress in Neurobiology GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef] [PubMed]
- Brahmachari, S.; Fung, Y.K.; Pahan, K. Induction of Glial Fibrillary Acidic Protein Expression in Astrocytes by Nitric Oxide. J. Neurosci. 2006, 26, 4930–4939. [Google Scholar] [CrossRef] [PubMed]
- Menet, V.; Gimenez y Ribotta, M.; Chauvet, N.; Drian, M.J.; Lannoy, J.; Colucci-Guyon, E.; Privat, A. Inactivation of the Glial Fibrillary Acidic Protein Gene, But Not That of Vimentin, Improves Neuronal Survival and Neurite Growth by Modifying Adhesion Molecule Expression. J. Neurosci. 2001, 21, 6147–6158. [Google Scholar] [CrossRef]
- Brenner, M. Role of GFAP in CNS injuries Michael. Neurosci. Lett. 2014, 565, 7–13. [Google Scholar] [CrossRef]
- Atwood, B.K.; MacKie, K. CB 2: A cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef]
- Schmöle, A.-C.; Lundt, R.; Gennequin, B.; Schrage, H. Expression Analysis of CB2-GFP BAC Transgenic Mice. PLoS ONE 2015, 10, e0138986. [Google Scholar] [CrossRef]
- Baur, R.; Gertsch, J.; Sigel, E. Do N-arachidonyl-glycine (NA-glycine) and 2-arachidonoyl glycerol (2-AG) share mode of action and the binding site on the β2 subunit of GABAA receptors? PeerJ 2013, 1, e149. [Google Scholar] [CrossRef]
- McCoy, K.L. Interaction between Cannabinoid System and Toll-Like Receptors Controls Inflammation. Mediators Inflamm. 2016, 2016, 5831315. [Google Scholar] [CrossRef]
- Vasudevan, K.M.; Garraway, L.A. Intradermal Rabies Vaccination: The Evolution and Future of Pre- and Post-exposure Prophylaxis. Curr. Top. Microbiol. Immunol. 2010, 347, 105–133. [Google Scholar] [CrossRef]
- Tarassishin, L.; Suh, H.-S.; Lee, S.C. Interferon regulatory factor 3 plays an anti-inflammatory role in microglia by activating the PI3K/Akt pathway. J. Neuroinflamm. 2011, 8, 187. [Google Scholar] [CrossRef]
- McHugh, D.; Roskowski, D.; Xie, S.; Bradshaw, H.B. Δ9-THC and N-arachidonoyl glycine regulate BV-2 microglial morphology and cytokine release plasticity: Implications for signaling at GPR18. Front. Pharmacol. 2014, 4, 162. [Google Scholar] [CrossRef]
- Console-Bram, L.; Brailoiu, E.; Brailoiu, G.C.; Sharir, H.; Abood, M.E. Activation of GPR18 by cannabinoid compounds: A tale of biased agonism. Br. J. Pharmacol. 2014, 171, 3908–3917. [Google Scholar] [CrossRef]
- Dehghani, F.; Hischebeth, G.T.R.; Wirjatijasa, F.; Kohl, A.; Korf, H.W.; Hailer, N.P. The immunosuppressant mycophenolate mofetil attenuates neuronal damage after excitotoxic injury in hippocampal slice cultures. Eur. J. Neurosci. 2003, 18, 1061–1072. [Google Scholar] [CrossRef]
- Benz, A.H.; Shajari, M.; Peruzki, N.; Dehghani, F.; Maronde, E. Early growth response-1 induction by fibroblast growth factor-1 via increase of mitogen-activated protein kinase and inhibition of protein kinase B in hippocampal neurons. Br. J. Pharmacol. 2010, 160, 1621–1630. [Google Scholar] [CrossRef]
- Blasi, E.; Barluzzi, R.; Bocchini, V.; Mazzolla, R.; Bistoni, F. Immortalization of murine microglial cells by a v-raf/v-myc carrying retrovirus. J. Neuroimmunol. 1990, 27, 229–237. [Google Scholar] [CrossRef]
- Morimoto, B.H.; Koshland, E. Induction and Expression of long- and Short-Term Neurosecretory Potentiation in a Neural Cell line. Neuron 1990, 5, 875–880. [Google Scholar] [CrossRef]
- Davis, J.B.; Maher, P. Protein kinase C activation inhibits glutamate-induced cytotoxicity in a neuronal cell line. Brain Res. 1994, 652, 169–173. [Google Scholar] [CrossRef]
- Chomczynski, P. Single-Step Method of RNA Isolation by Acid Guanidinium Extraction. Anal. Biochem. 1987, 159, 156–159. [Google Scholar] [CrossRef]
- Ebrahimi, F.; Hezel, M.; Koch, M.; Ghadban, C.; Korf, H.W.; Dehghani, F. Analyses of neuronal damage in excitotoxically lesioned organotypic hippocampal slice cultures. Ann. Anat. 2010, 192, 199–204. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Left | Right | Product Size | Sequence |
|---|---|---|---|---|
| ß-actin | ACTCCTACGTGGGCGACGAGG | CAGGTCCAGACGCAGGATGGC | 389 bp | NM_007393 |
| gpr18 | TGAAGCCCAAGGTCAAGGAGAAGT | TTCATGAGGAAGGTGGTGAAGGCT | 163 bp | NM_182806.1 |
| Antibody | Concentration | Company | Article Number | Antibody ID |
|---|---|---|---|---|
| ß-actin | 1:5000 | Cell Signaling, Boston, USA | 3700 | AB_2242334 |
| pAkt | 1:2000 | Cell Signaling | 9271 | AB_329825 |
| phospho-CREB (Ser133) | 1:300 | Merck Millipore | 06-519 | AB_310153 |
| t-CREB clone E306 | 1:1000 | Merck Millipore | 04-218 | AB_1586958 |
| GAPDH (14C10) | 1:1000 | Cell Signaling | 2188 | AB_561053 |
| GPR18 | 1:5000 | Dr Ken Mackie’s Lab | Antibody is described in [9] | |
| phospho-p38 MAPK (Thr180/Tyr182) | 1:2000 | Cell Signaling | 4511S | AB_2139682 |
| t-p38 MAPK | 1:2000 | Cell Signaling | 9212S | AB_330713 |
| phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) | 1:4000 | Cell Signaling | 9101 | AB_331646 |
| t-p44/42 MAPK (Erk1/2) | 1:4000 | Cell Signaling | 9102 | AB_330744 |
| horse anti-mouse IgG, HRP conjugated | 1:10000 | Vector laboratories, Burlingame, USA | PI-2000 | AB_2336177 |
| goat anti-rabbit IgG, HRP conjugated | 1:20000 | Vector laboratories | PI-1000 | AB_2336198 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabiec, U.; Hohmann, T.; Ghadban, C.; Rothgänger, C.; Wong, D.; Antonietti, A.; Groth, T.; Mackie, K.; Dehghani, F. Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures. Int. J. Mol. Sci. 2019, 20, 1266. https://doi.org/10.3390/ijms20061266
Grabiec U, Hohmann T, Ghadban C, Rothgänger C, Wong D, Antonietti A, Groth T, Mackie K, Dehghani F. Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures. International Journal of Molecular Sciences. 2019; 20(6):1266. https://doi.org/10.3390/ijms20061266
Chicago/Turabian StyleGrabiec, Urszula, Tim Hohmann, Chalid Ghadban, Candy Rothgänger, Daniel Wong, Alexandra Antonietti, Thomas Groth, Ken Mackie, and Faramarz Dehghani. 2019. "Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures" International Journal of Molecular Sciences 20, no. 6: 1266. https://doi.org/10.3390/ijms20061266
APA StyleGrabiec, U., Hohmann, T., Ghadban, C., Rothgänger, C., Wong, D., Antonietti, A., Groth, T., Mackie, K., & Dehghani, F. (2019). Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures. International Journal of Molecular Sciences, 20(6), 1266. https://doi.org/10.3390/ijms20061266