Plasma Levels of Preβ1-HDL Are Significantly Elevated in Non-Dialyzed Patients with Advanced Stages of Chronic Kidney Disease

 , , , and
, , , and
Abstract
:
1. Introduction
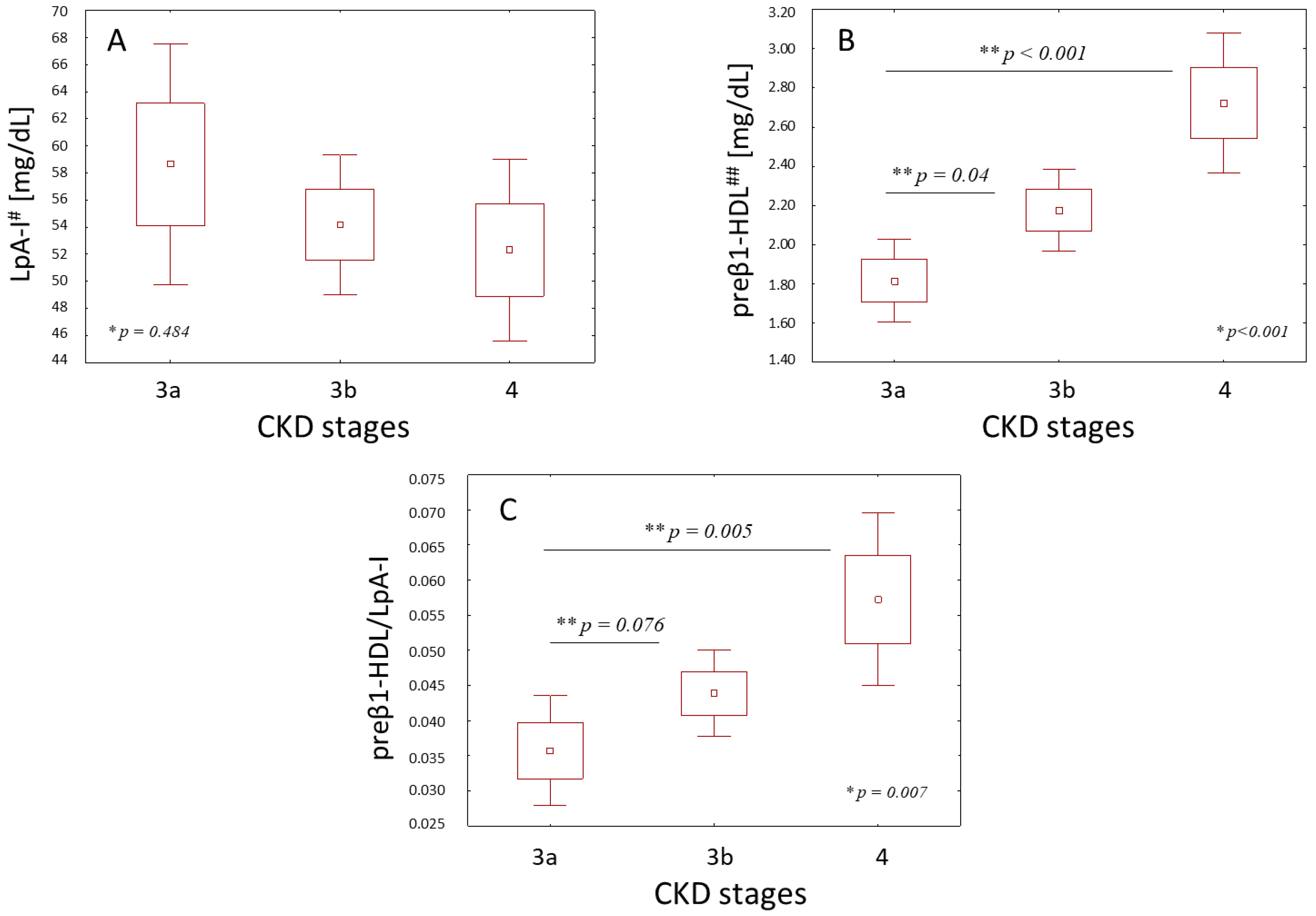
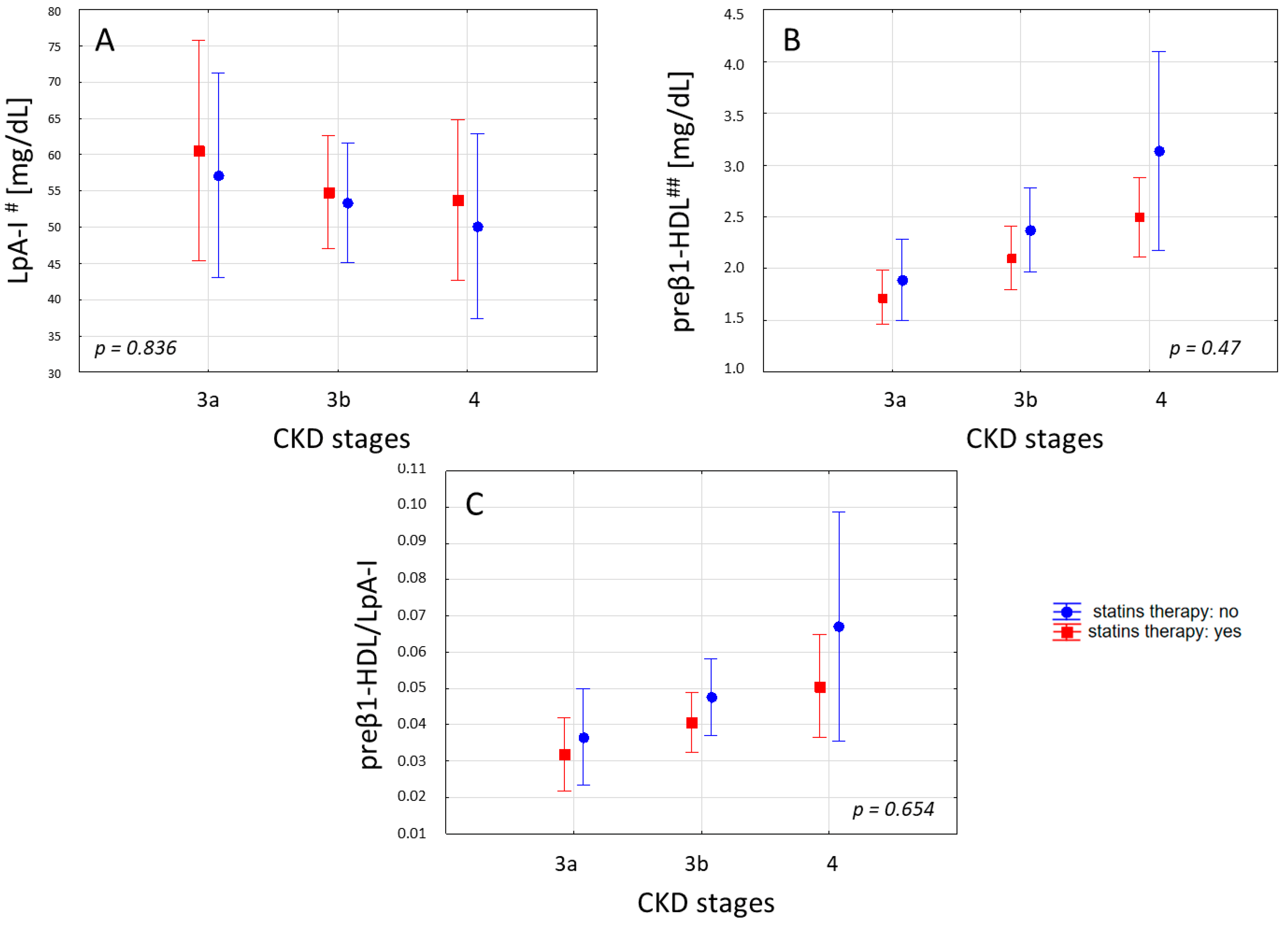
2. Results
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Laboratory Measurements
4.3. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Go, A.S.; Chertow, G.M.; Fan, D.; McCulloch, C.E.; Hsu, C.Y. Chronic kidney disease and the risks of death, cardiovascular events, and hospitalization. N. Engl. J. Med. 2004, 13, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Gluba-Brzozka, A.; Michalska-Kasiczak, M.; Franczyk-Skora, B.; Nocun, M.; Banach, M.; Rysz, J. Markers of increased cardiovascular risk in patients with chronic kidney disease. Lipids Health Dis. 2014, 13, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronda, E.; Genovese, S.; Padeletti, L.; Cacciatore, F.; Vitale, D.F.; Bragato, R.; Innocenti, L.; Schiano, C.; Sommese, L.; De Pascale, M.R.; et al. Renal function impairment predicts mortality in patients with chronic heart failure treated with resynchronization therapy. Cardiol. J. 2015, 4, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Navab, M.; Fogelman, A.M. HDL metabolism and activity in chronic kidney disease. Nat. Rev. Nephrol. 2010, 5, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Reiss, A.B.; Voloshyna, I.; DeLeon, J.; Miyawaki, N.; Mattana, J. Cholesterol Metabolism in CKD. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 2015, 6, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Moradi, H.; Vaziri, N.D.; Said, H.M.; Kalantar-Zadeh, K. Role of HDL dysfunction in end-stage renal disease: A doubl-edeged sword. J. Ren. Nutr. Off. J. Council Ren. Nutr. Natl. Kidney Found. 2013, 3, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Mangé, A.; Goux, A.; Badiou, S.; Patrier, L.; Canaud, B.; Maudelonde, T.; Cristol, J.-P.; Solassol, J. HDL Proteome in Hemodialysis Patients: A Quantitative Nanoflow Liquid Chromatography-Tandem Mass Spectrometry Approach. PLoS ONE 2012, 3, e34107. [Google Scholar] [CrossRef] [PubMed]
- Holzer, M.; Birner-Gruenberger, R.; Stojakovic, T.; El-Gamal, D.; Binder, V.; Wadsack, C.; Heinemann, A.; Marsche, G. Uremia Alters HDL Composition and Function. J. Am. Soc. Nephrol. JASN 2011, 9, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Stampfer, M.J.; Sacks, F.M.; Salvini, S.; Willett, W.C.; Hennekens, C.H. A prospective study of cholesterol, apolipoproteins, and the risk of myocardial infarction. N. Engl. J. Med. 1991, 6, 373–381. [Google Scholar] [CrossRef]
- Luc, G.; Bard, J.M.; Ferrières, J.; Evans, A.; Amouyel, P.; Arveiler, D.; Fruchart, J.C.; Ducimetière, P. Value of HDL cholesterol, apolipoprotein A-I, lipoprotein A-I, and lipoprotein A-I/A-II in prediction of coronary heart disease: The PRIME Study. Prospective Epidemiological Study of Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2002, 7, 1155–1161. [Google Scholar] [CrossRef]
- Puchois, P.; Kandoussi, A.; Fievet, P.; Fourrier, J.L.; Bertrand, M.; Koren, E.; Fruchart, J.C. Apolipoprotein A-I containing lipoproteins in coronary artery disease. Atherosclerosis 1987, 68, 35–40. [Google Scholar] [CrossRef]
- Sethi, A.A.; Sampson, M.; Warnick, R.; Muniz, N.; Vaisman, B.; Nordestgaard, B.G.; Tybjaerg-Hansen, A.; Remaley, A.T. High pre-beta1 HDL concentrations and low lecithin: Cholesterol acyltransferase activities are strong positive risk markers for ischemic heart disease and independent of HDL-cholesterol. Clin. Chem. 2010, 7, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, J.; Miyazaki, O.; Nakamura, Y.; Miyazaki, A.; Fukamachi, I.; Bujo, H.; Saito, Y. Plasma pre beta1-HDL level is elevated in unstable angina pectoris. Atherosclerosis 2009, 2, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Miida, T.; Miyazaki, O.; Hanyu, O.; Nakamura, Y.; Hirayama, S.; Narita, I.; Gejyo, F.; Ei, I.; Tasaki, K.; Kohda, Y.; et al. LCAT-dependent conversion of prebeta1-HDL into alpha-migrating HDL is severely delayed in hemodialysis patients. J. Am. Soc. Nephrol. 2003, 3, 732–738. [Google Scholar] [CrossRef]
- Kontush, A.; Chapman, M.J. Antiatherogenic function of HDL particle subpopulations: Focus on antioxidative activities. Curr. Opin. Lipidol. 2010, 4, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Nukuna, B.; Brennan, M.L.; Sun, M.; Goormastic, M.; Settle, M.; Schmitt, D.; Fu, X.; Thomson, L.; Fox, P.L.; et al. Apolipoprotein A-I is a selective target for myeloperoxidase-catalyzed oxidation and functional impairment in subjects with cardiovascular disease. J. Clin. Investig. 2004, 4, 529–541. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, Z.; Riwanto, M.; Gao, S.; Levison, B.S.; Gu, X.; Fu, X.; Wagner, M.A.; Besler, C.; Gerstenecker, G.; et al. Myeloperoxidase, paraoxonase-1, and HDL form a functional ternary complex. J. Clin. Investig. 2013, 9, 3815–3828. [Google Scholar] [CrossRef]
- Matsushita, K.; Van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; De Jong, P.E.; Coresh, J.; Gansevoort, R.T. Association of estimated glomerular filtration rate and albuminuria with all-cause and cardiovascular mortality in general population cohorts: A collaborative meta-analysis. Lancet 2010, 9731, 2073–2081. [Google Scholar]
- Kuchta, A.; Strzelecki, A.; Ćwiklińska, A.; Gruchała, M.; Zdrojewski, Z.; Kortas-Stempak, B.; Wieczorek, E.; Gliwińska, A.; Dąbkowski, K.; Jankowski, M. HDL subpopulations containing apoA-I without apoA-II (LpA-I) in patients with angiographically proven coronary artery disease. J. Cardiol. 2017, 3, 523–528. [Google Scholar] [CrossRef]
- Bu, X.-M.; Niu, D.-M.; Wu, J.; Yuan, Y.-L.; Song, J.-X.; Wang, J.-J. Elevated levels of preβ1-high-density lipoprotein are associated with cholesterol ester transfer protein, the presence and severity of coronary artery disease. Lipids Health Dis. 2017, 16, 4. [Google Scholar] [CrossRef]
- Chen, Y.; Dong, J.; Chen, X.; Jiang, H.; Bakillah, A.; Zhang, X.; Li, Z.; Yin, J.; Liang, D.; Zou, Y.; et al. Human serum preβ1-high density lipoprotein levels are independently and negatively associated with coronary artery diseases. Nutr. Metab. 2016, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Liang, K.; Parks, J.S. Down-regulation of hepatic lecithin:cholesterol acyltransferase gene expression in chronic renal failure. Kidney Int. 2001, 6, 2192–2196. [Google Scholar] [CrossRef] [PubMed]
- Calabresi, L.; Simonelli, S.; Conca, P.; Busnach, G.; Cabibbe, M.; Gesualdo, L.; Gigante, M.; Penco, S.; Veglia, F.; Franceschini, G. Acquired lecithin:cholesterol acyltransferase deficiency as a major factor in lowering plasma HDL levels in chronic kidney disease. J. Intern. Med. 2015, 5, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Asztalos, B.F.; Demissie, S.; Cupples, L.A.; Collins, D.; Cox, C.E.; Horvath, K.V.; Bloomfield, H.E.; Robins, S.J.; Schaefer, E.J. LpA-I, LpA-I:A-II HDL and CHD-risk: The Framingham Offspring Study and the Veterans Affairs HDL Intervention Trial. Atherosclerosis 2006, 1, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Hattori, S.; Nishiyama, S.; Higashi, A.; Matsuda, I. Quantitative and qualitative changes of apolipoprotein AI-containing lipoproteins in patients on continuous ambulatory peritoneal dialysis. Metabolism 1989, 9, 843–849. [Google Scholar] [CrossRef]
- Durrington, P.N.; Mackness, B.; Mackness, M.I. Paraoxonase and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2001, 4, 473–480. [Google Scholar] [CrossRef]
- Miljkovic, M.; Stefanovic, A.; Vekic, J.; Zeljkovic, A.; Gojkovic, T.; Simic-Ogrizovic, S.; Bogavac-Stanojevic, N.; Cerne, D.; Ilic, J.; Stefanovic, I.; et al. Activity of paraoxonase 1 (PON1) on HDL2 and HDL3 subclasses in renal disease. Clin. Biochem. 2018, 60, 52–58. [Google Scholar] [CrossRef]
- Malle, E.; Marsche, G.; Panzenboeck, U.; Sattler, W. Myeloperoxidase-mediated oxidation of high-density lipoproteins: Fingerprints of newly recognized potential proatherogenic lipoproteins. Arch. Biochem. Biophys. 2006, 2, 245–255. [Google Scholar] [CrossRef]
- Teng, N.; Maghzal, G.J.; Talib, J.; Rashid, I.; Lau, A.K.; Stocker, R. The roles of myeloperoxidase in coronary artery disease and its potential implication in plaque rupture. Redox Rep. 2017, 2, 51–73. [Google Scholar] [CrossRef]
- Kowalska, K.; Socha, E.; Milnerowicz, H. Review: The role of paraoxonase in cardiovascular diseases. Ann. Clin. Lab. Sci. 2015, 2, 226–233. [Google Scholar]
- Morena, M.; Cristol, J.P.; Dantoine, T.; Carbonneau, M.A.; Descomps, B.; Canaud, B. Protective effects of high-density lipoprotein against oxidative stress are impaired in haemodialysis patients. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc. Eur. Ren. Assoc. 2000, 3, 389–395. [Google Scholar] [CrossRef]
- Kuchta, A.; Pacanis, A.; Kortas-Stempak, B.; Cwiklińska, A.; Ziętkiewicz, M.; Renke, M.; Rutkowski, B. Estimation of Oxidative Stress Markers in Chronic Kidney Disease. Kidney Blood Press Res. 2010, 1, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.Z.; Varghese, Z.; Moorhead, J.F. An update on the lipid nephrotoxicity hypothesis. Nat. Rev. Nephrol. 2009, 12, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Wanner, C.; Krane, V.; Marz, W.; Olschewski, M.; Mann, J.F.; Ruf, G.; Ritz, E. Atorvastatin in patients with type 2 diabetes mellitus undergoing hemodialysis. N. Engl. J. Med. 2005, 3, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Krane, V.; Winkler, K.; Drechsler, C.; Lilienthal, J.; Marz, W.; Wanner, C. Effect of atorvastatin on inflammation and outcome in patients with type 2 diabetes mellitus on hemodialysis. Kidney Int. 2008, 11, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Fellstrom, B.C.; Jardine, A.G.; Schmieder, R.E.; Holdaas, H.; Bannister, K.; Beutler, J.; Chae, D.W.; Chevaile, A.; Cobbe, S.M.; Gronhagen-Riska, C.; et al. Rosuvastatin and cardiovascular events in patients undergoing hemodialysis. N. Engl. J. Med. 2009, 14, 1395–1407. [Google Scholar] [CrossRef]
- Asztalos, B.F.; Horvath, K.V.; McNamara, J.R.; Roheim, P.S.; Rubinstein, J.J.; Schaefer, E.J. Comparing the effects of five different statins on the HDL subpopulation profiles of coronary heart disease patients. Atherosclerosis 2002, 2, 361–369. [Google Scholar] [CrossRef]
- Franceschini, G.; Calabresi, L.; Colombo, C.; Favari, E.; Bernini, F.; Sirtori, C.R. Effects of fenofibrate and simvastatin on HDL-related biomarkers in low-HDL patients. Atherosclerosis 2007, 2, 385–391. [Google Scholar] [CrossRef]
- Tian, L.; Chen, Y.; Li, C.; Zeng, Z.; Xu, Y.; Long, S.; Fu, M. Statin treatment improves plasma lipid levels but not HDL subclass distribution in patients undergoing percutaneous coronary intervention. Lipids. 2013, 2, 127–137. [Google Scholar] [CrossRef]
- Miyazaki, O.; Kobayashi, J.; Fukamachi, I.; Miida, T.; Bujo, H.; Saito, Y. A new sandwich enzyme immunoassay for measurement of plasma pre-beta1-HDL levels. J. Lipid Res. 2000, 12, 2083–2088. [Google Scholar]
- Nakanishi, M.; Takanami, Y.; Maruyama, T.; Murata, M.; Motohashi, Y.; Nakano, S.; Uchida, K.; Maruyama, C.; Kyotani, S.; Tsushima, M. The ratio of serum paraoxonase/arylesterase activity using an improved assay for arylesterase activity to discriminate PON1(R192) from PON1(Q192). J. Atheroscler. Thromb. 2003, 6, 337–342. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Parameter | Stages of CKD | |||
|---|---|---|---|---|
| 3a | 3b | 4 | p-Value | |
| Gender (M/F) | 12/5 | 19/15 | 12/5 | 0.531 ** |
| Age (years) | 69 ± 5 | 70 ± 9 | 63 ± 5 | 0.06 * |
| BMI (kg/m2) | 28 ± 3 | 29 ± 5 | 26 ± 4 | 0.306 * |
| eGFR | 50 ± 3 | 37 ± 4 | 22 ± 4 | <0.001 * |
| Albumin (g/L) | 43.6 ± 3.2 | 42.8 ± 2.9 | 42.9 ± 2.9 | 0.674 * |
| Statin therapy (%) | 41 | 53 | 64 | 0.333 ** |
| TAG (mg/dL) | 102 ± 30 | 117 ± 45 | 135 ± 63 | 0.215 * |
| TC (mg/dL) | 199 ± 42 | 200 ± 53 | 215 ± 36 | 0.345 * |
| HDL-C (mg/dL) | 51 ± 11 | 50 ± 12 | 48 ± 11 | 0.761 * |
| LDL-C (mg/dL) | 127 ± 39 | 127 ± 49 | 140 ± 33 | 0.335 * |
| ApoA-I (mg/dL) | 172 ± 27 | 164 ± 27 | 160 ± 23 | 0.297 * |
| ApoA-II (mg/dL) | 33 ± 7 | 31 ± 6 | 31 ± 5 | 0.499 * |
| LCAT (390/470 nm) | 1.34 ± 0.03 | 1.32 ± 0.05 | 1.32 ± 0.04 | 0.458 * |
| FC/TC | 0.283 ± 0.05 | 0.283 ± 0.03 | 0.290 ± 0.03 | 0.234 * |
| Parameter | R | p |
|---|---|---|
| HDL-C (mg/dL) | 0.104 | 0.382 |
| ApoA-I (mg/dL) | 0.153 | 0.202 |
| ApoA-II (mg/dL) | 0.109 | 0.358 |
| LpA-I (mg/dL) | 0.025 | 0.837 |
| preβ1-HDL (mg/dL) | −0.456 | <0.001 |
| preβ1-HDL/LpA-I | −0.322 | 0.008 |
| PON-1 (U/L) | 0.068 | 0.566 |
| MPO (ng/mL) | 0.079 | 0.531 |
| LCAT (390/470 nm) | 0.080 | 0.484 |
| Parameter | β | SE | p |
|---|---|---|---|
| preβ1-HDL | −0.41 | 0.105 | <0.001 |
| preβ1-HDL/LpA-I | −0.33 | 0.09 | 0.001 |
| Stages of CKD | ||||
|---|---|---|---|---|
| 3a | 3b | 4 | p-Value * | |
| PON-1 (U/L) | 102 (53–150) | 83 (52–152) | 113 (80–130) | 0.890 |
| MPO (ng/mL) | 235 (136–392) | 199 (139–347) | 273 (160–327) | 0.377 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuchta, A.; Ćwiklińska, A.; Czaplińska, M.; Wieczorek, E.; Kortas-Stempak, B.; Gliwińska, A.; Dąbkowski, K.; Sałaga-Zaleska, K.; Mickiewicz, A.; Dębska-Ślizień, A.; et al. Plasma Levels of Preβ1-HDL Are Significantly Elevated in Non-Dialyzed Patients with Advanced Stages of Chronic Kidney Disease. Int. J. Mol. Sci. 2019, 20, 1202. https://doi.org/10.3390/ijms20051202
Kuchta A, Ćwiklińska A, Czaplińska M, Wieczorek E, Kortas-Stempak B, Gliwińska A, Dąbkowski K, Sałaga-Zaleska K, Mickiewicz A, Dębska-Ślizień A, et al. Plasma Levels of Preβ1-HDL Are Significantly Elevated in Non-Dialyzed Patients with Advanced Stages of Chronic Kidney Disease. International Journal of Molecular Sciences. 2019; 20(5):1202. https://doi.org/10.3390/ijms20051202
Chicago/Turabian StyleKuchta, Agnieszka, Agnieszka Ćwiklińska, Monika Czaplińska, Ewa Wieczorek, Barbara Kortas-Stempak, Anna Gliwińska, Kamil Dąbkowski, Kornelia Sałaga-Zaleska, Agnieszka Mickiewicz, Alicja Dębska-Ślizień, and et al. 2019. "Plasma Levels of Preβ1-HDL Are Significantly Elevated in Non-Dialyzed Patients with Advanced Stages of Chronic Kidney Disease" International Journal of Molecular Sciences 20, no. 5: 1202. https://doi.org/10.3390/ijms20051202
APA StyleKuchta, A., Ćwiklińska, A., Czaplińska, M., Wieczorek, E., Kortas-Stempak, B., Gliwińska, A., Dąbkowski, K., Sałaga-Zaleska, K., Mickiewicz, A., Dębska-Ślizień, A., Król, E., & Jankowski, M. (2019). Plasma Levels of Preβ1-HDL Are Significantly Elevated in Non-Dialyzed Patients with Advanced Stages of Chronic Kidney Disease. International Journal of Molecular Sciences, 20(5), 1202. https://doi.org/10.3390/ijms20051202