Role of HYBID (Hyaluronan Binding Protein Involved in Hyaluronan Depolymerization), Alias KIAA1199/CEMIP, in Hyaluronan Degradation in Normal and Photoaged Skin

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Diverse Properties and Rapid Turnover of HA in the Skin
3. Molecular Mechanism of HA Synthesis in Skin Fibroblasts
4. HA Degradation Mediated by Newly Discovered HYBID in Skin Fibroblasts
5. Characteristics of HYBID-Mediated HA Degradation
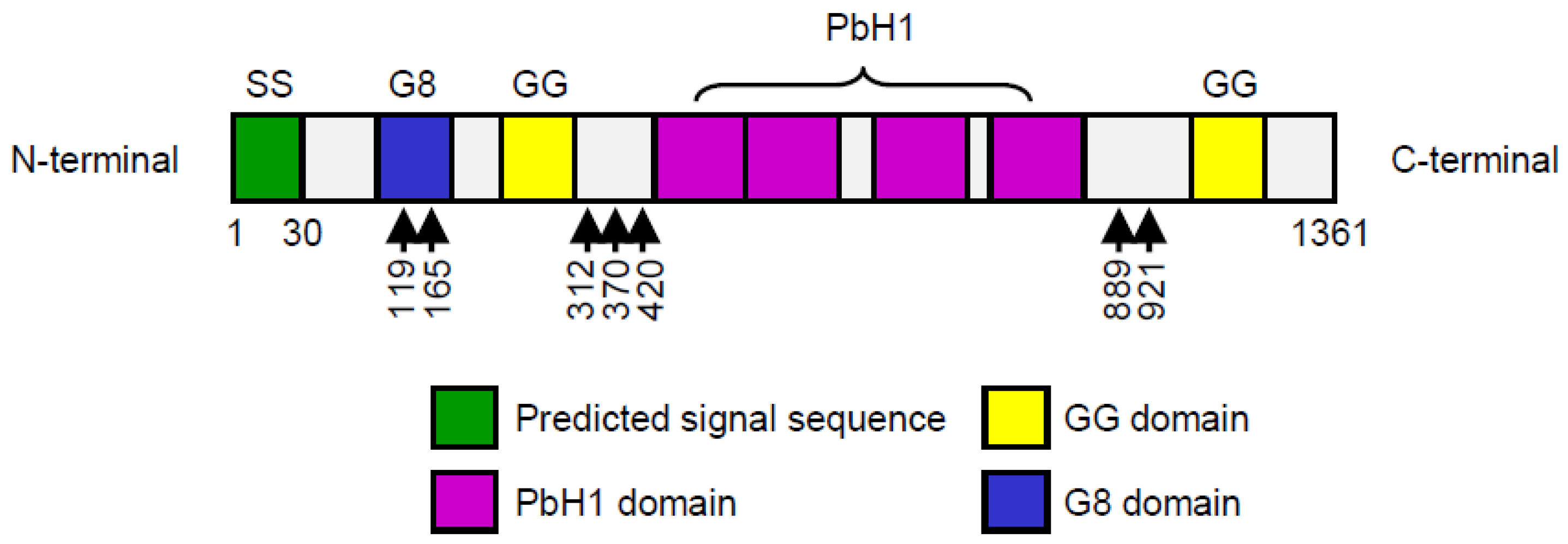
5.1. Molecular Function of HYBID Protein
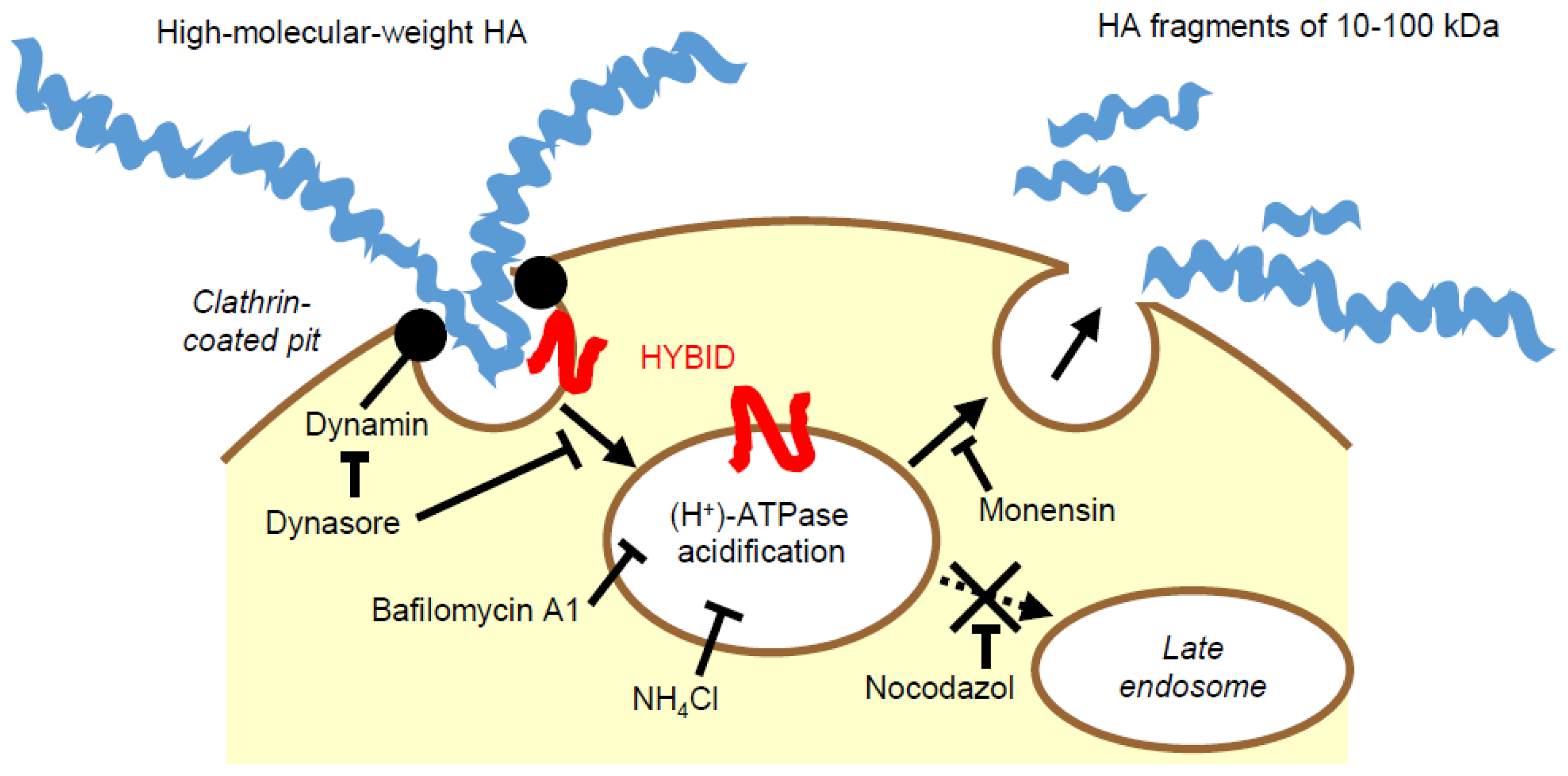
5.2. Cellular Mechanism of HYBID-Mediated HA Degradation
5.3. Characterization of Murine Homologue (mHybid) of Human HYBID
6. Regulation of Expression of HYBID and HAS by Growth Factors in Skin Fibroblasts
7. Possible Involvement of HYBID-Mediated HA Degradation in Photoaging Skin Symptoms
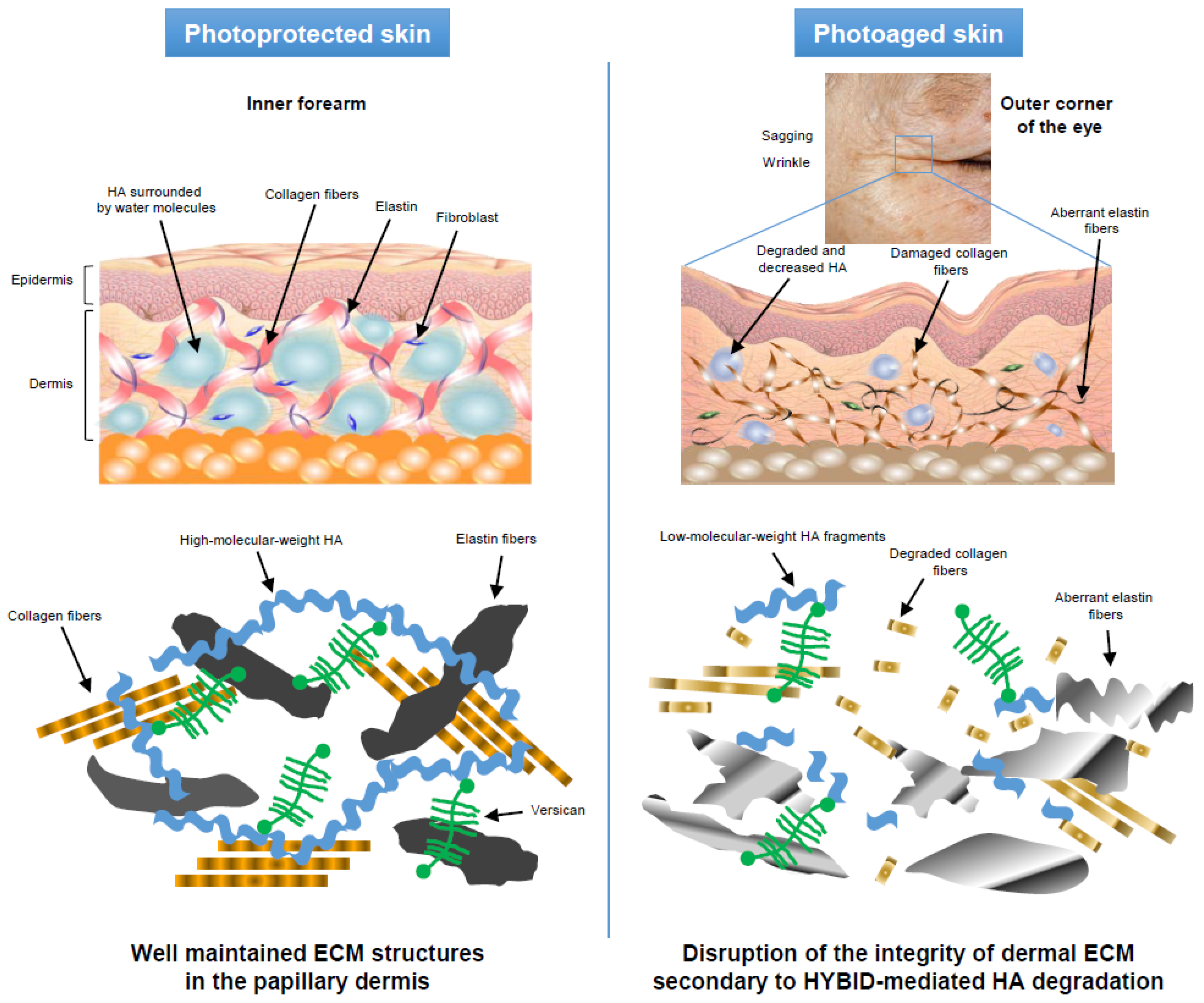
7.1. Decreased HA in the Papillary Dermis of Photoaged Skin and Its Correlation with Photoaging Skin Symptoms
7.2. Overexpression of HYBID in Photoaged Skin and Its Correlation with HA Amount in Papillary Dermis and Photoaging Skin Symptoms
7.3. Relationship of HA and HYBID Expression with Wrinkling of Photoaged Skin in Caucasian Females
8. Screening of Agents that Inhibit the Expression and Activity of HYBID and Their Anti-Wrinkle Effects
9. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| bFGF | basic fibroblast growth factor |
| CHC | clathrin heavy chain |
| ECM | extracellular matrix |
| EGF | epidermal growth factor |
| GAG | glycosaminoglycan |
| HA | hyaluronan |
| HAS | hyaluronan synthase |
| HYAL | hyaluronidase |
| HYBID | hyaluronan binding protein involved in hyaluronan depolymerization |
| MMP | matrix metalloprotease |
| PDGF | platelet-derived growth factor |
| ROS | reactive oxygen species |
| siRNA | short interfering RNA |
| TGF | transforming growth factor |
| TMEM2 | transmembrane protein 2 |
| UV | ultraviolet |
| VEGF | vascular endothelial growth factor |
References
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine aspects of skin aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R.M.; Kligman, A.M. Chronic heliodermatitis: A morphologic evaluation of chronic actinic dermal damage with emphasis on the role of mast cells. J. Investig. Dermatol. 1988, 90, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Baumann, L. Skin ageing and its treatment. J. Pathol. 2007, 211, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Uitto, J. The role of elastin and collagen in cutaneous aging: Intrinsic aging versus photoexposure. J. Drugs. Dermatol. 2008, 7, 12–16. [Google Scholar]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef]
- Jurzak, M.; Latocha, M.; Gojniczek, K.; Kapral, M.; Garncarczyk, A.; Pierzchala, E. Influence of retinoids on skin fibroblasts metabolism in vitro. Acta Pol. Pharm. 2008, 65, 85–91. [Google Scholar]
- Zouboulis, C.C.; Boschnakow, A. Chronological ageing and photoageing of the human sebaceous gland. Clin. Exp. Dermatol. 2001, 26, 600–607. [Google Scholar] [CrossRef]
- Sakai, S.; Nakagawa, N.; Yamanari, M.; Miyazawa, A.; Yasuno, Y.; Matsumoto, M. Relationship between dermal birefringence and the skin surface roughness of photoaged human skin. J. Biomed. Opt. 2009, 14, 044032. [Google Scholar] [CrossRef]
- Bernstein, E.F.; Chen, Y.Q.; Tamai, K.; Shepley, K.J.; Resnik, K.S.; Zhang, H.; Tuan, R.; Mauviel, A.; Uitto, J. Enhanced elastin and fibrillin gene expression in chronically photodamaged skin. J. Investig. Dermatol. 1994, 103, 182–186. [Google Scholar] [PubMed]
- Hasegawa, K.; Yoneda, M.; Kuwabara, H.; Miyaishi, O.; Itano, N.; Ohno, A.; Zako, M.; Isogai, Z. Versican, a major hyaluronan-binding component in the dermis, loses its hyaluronan-binding ability in solar elastosis. J. Investig. Dermatol. 2007, 127, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Nagaoka, A.; Komiya, A.; Aoki, M.; Nakamura, S.; Morikawa, T.; Ohtsuki, R.; Sayo, T.; Okada, Y.; Takahashi, Y. Reduction of hyaluronan and increased expression of HYBID (alias CEMIP and KIAA1199) correlate with clinical symptoms in photoaged skin. Br. J. Dermatol. 2018, 179, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol. 2001, 20, 499–508. [Google Scholar] [CrossRef]
- Yoshida, H.; Nagaoka, A.; Kusaka-Kikushima, A.; Tobiishi, M.; Kawabata, K.; Sayo, T.; Sakai, S.; Sugiyama, Y.; Enomoto, H.; Okada, Y.; et al. KIAA1199, a deafness gene of unknown function, is a new hyaluronan binding protein involved in hyaluronan depolymerization. Proc. Natl. Acad. Sci. USA 2013, 110, 5612–5617. [Google Scholar] [CrossRef]
- Nagaoka, A.; Yoshida, H.; Nakamura, S.; Morikawa, T.; Kawabata, K.; Kobayashi, M.; Sakai, S.; Takahashi, Y.; Okada, Y.; Inoue, S. Regulation of hyaluronan (HA) metabolism mediated by HYBID (hyaluronan-binding protein involved in HA depolymerization, KIAA1199) and HA synthases in growth factor-stimulated fibroblasts. J. Biol. Chem. 2015, 290, 30910–30923. [Google Scholar] [CrossRef]
- Evensen, N.A.; Li, Y.; Kuscu, C.; Liu, J.; Cathcart, J.; Banach, A.; Zhang, Q.; Li, E.; Joshi, S.; Yang, J.; et al. Hypoxia promotes colon cancer dissemination through up-regulation of cell migration-inducing protein (CEMIP). Oncotarget 2015, 6, 20723–20739. [Google Scholar] [CrossRef]
- Yoshida, H.; Komiya, A.; Ohtsuki, R.; Kusaka-Kikushima, A.; Sakai, S.; Kawabata, K.; Kobayashi, M.; Nakamura, S.; Nagaoka, A.; Sayo, T.; et al. Relationship of hyaluronan and HYBID (KIAA1199) expression with roughness parameters of photoaged skin in Caucasian women. Skin Res. Technol. 2018, 24, 562–569. [Google Scholar] [CrossRef]
- Laurent, T.C.; Fraser, J.R. Hyaluronan. FASEB J. 1992, 6, 2397–2404. [Google Scholar] [CrossRef]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef]
- Pandey, M.S.; Harris, E.N.; Weigel, J.A.; Weigel, P.H. The cytoplasmic domain of the hyaluronan receptor for endocytosis (HARE) contains multiple endocytic motifs targeting coated pit-mediated internalization. J. Biol. Chem. 2008, 283, 21453–21461. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Kimata, K. Mammalian hyaluronan synthases. IUBMB Life 2002, 54, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, Y.; Shimada, A.; Sayo, T.; Sakai, S.; Inoue, S. Putative hyaluronan synthase mRNA are expressed in mouse skin and TGF-b upregulates their expression in cultured human skin cells. J. Investig. Dermatol. 1998, 110, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Usami, S.; Nakamura, Y. Mutations in the gene encoding KIAA1199 protein, an inner-ear protein expressed in Deiters’ cells and the fibrocytes, as the cause of nonsyndromic hearing loss. J. Hum. Genet. 2003, 48, 564–570. [Google Scholar] [CrossRef]
- Yamamoto, H.; Tobisawa, Y.; Inubushi, T.; Irie, F.; Ohyama, C.; Yamaguchi, Y. A mammalian homolog of the zebrafish transmembrane protein 2 (TMEM2) is the long-sought-after cell-surface hyaluronidase. J. Biol. Chem. 2017, 292, 7304–7313. [Google Scholar] [CrossRef]
- Yoshino, Y.; Goto, M.; Hara, H.; Inoue, S. The role and regulation of TMEM2 (transmembrane protein 2) in HYBID (hyaluronan (HA)-binding protein involved in HA depolymerization/KIAA1199/CEMIP)-mediated HA depolymerization in human skin fibroblasts. Biochem. Biophys. Res. Commun. 2018, 505, 74–80. [Google Scholar] [CrossRef]
- Michishita, E.; Garces, G.; Barrett, J.C.; Horikawa, I. Upregulation of the KIAA1199 gene is associated with cellular mortality. Cancer Lett. 2006, 239, 71–77. [Google Scholar] [CrossRef]
- Yoshida, H.; Nagaoka, A.; Nakamura, S.; Sugiyama, Y.; Okada, Y.; Inoue, S. Murine homologue of the human KIAA1199 is implicated in hyaluronan binding and depolymerization. FEBS Open Bio 2013, 3, 352–356. [Google Scholar] [CrossRef]
- Guo, J.; Cheng, H.; Zhao, S.; Yu, L. GG: A domain involved in phage LTF apparatus and implicated in human MEB and non-syndromic hearing loss diseases. FEBS Lett. 2006, 580, 581–584. [Google Scholar] [CrossRef]
- He, Q.Y.; Liu, X.H.; Li, Q.; Studholme, D.J.; Li, X.W.; Liang, S.P. G8: A novel domain associated with polycystic kidney disease and non-syndromic hearing loss. Bioinformatics 2006, 22, 2189–2191. [Google Scholar] [CrossRef]
- Birkenkamp-Demtroder, K.; Maghnouj, A.; Mansilla, F.; Thorsen, K.; Andersen, C.L.; Oster, B.; Hahn, S.; Orntoft, T.F. Repression of KIAA1199 attenuates Wnt-signalling and decreases the proliferation of colon cancer cells. Br. J. Cancer 2011, 105, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Yoshida, H.; Mizuno, S.; Hirozane, T.; Horiuchi, K.; Yoshino, Y.; Hara, H.; Kanai, Y.; Inoue, S.; Ishijima, M.; et al. Hyaluronan-binding protein involved in hyaluronan depolymerization dontrols endochondral ossification through hyaluronan metabolism. Am. J. Pathol. 2017, 187, 1162–1176. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Ishisaka, M.; Tsuruma, K.; Shimazawa, M.; Yoshida, H.; Inoue, S.; Shimoda, M.; Okada, Y.; Hara, H. Distribution and function of hyaluronan binding protein involved in hyaluronan depolymerization (HYBID, KIAA1199) in the mouse central nervous system. Neuroscience 2017, 347, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Shimazawa, M.; Nakamura, S.; Inoue, S.; Yoshida, H.; Shimoda, M.; Okada, Y.; Hara, H. Targeted deletion of HYBID (hyaluronan binding protein involved in hyaluronan depolymerization/ KIAA1199/CEMIP) decreases dendritic spine density in the dentate gyrus through hyaluronan accumulation. Biochem. Biophys. Res. Commun. 2018, 503, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Ellis, I.; Banyard, J.; Schor, S.L. Differential response of fetal and adult fibroblasts to cytokines: Cell migration and hyaluronan synthesis. Development 1997, 124, 1593–1600. [Google Scholar] [PubMed]
- Yamada, Y.; Itano, N.; Hata, K.; Ueda, M.; Kimata, K. Differential regulation by IL-1b and EGF of expression of three different hyaluronan synthases in oral mucosal epithelial cells and fibroblasts and dermal fibroblasts: Quantitative analysis using real-time RT-PCR. J. Investig. Dermatol. 2004, 122, 631–639. [Google Scholar] [CrossRef]
- Li, L.; Asteriou, T.; Bernert, B.; Heldin, C.H.; Heldin, P. Growth factor regulation of hyaluronan synthesis and degradation in human dermal fibroblasts: Importance of hyaluronan for the mitogenic response of PDGF-BB. Biochem. J. 2007, 404, 327–336. [Google Scholar] [CrossRef]
- Stern, R.; Asari, A.A.; Sugahara, K.N. Hyaluronan fragments: An information-rich system. Eur. J. Cell Biol. 2006, 85, 699–715. [Google Scholar] [CrossRef]
- West, D.C.; Hampson, I.N.; Arnold, F.; Kumar, S. Angiogenesis induced by degradation products of hyaluronic acid. Science 1985, 228, 1324–1326. [Google Scholar] [CrossRef]
- Rooney, P.; Kumar, S.; Ponting, J.; Wang, M. The role of hyaluronan in tumour neovascularization (review). Int. J. Cancer 1995, 60, 632–636. [Google Scholar] [CrossRef]
- Sugahara, K.N.; Murai, T.; Nishinakamura, H.; Kawashima, H.; Saya, H.; Miyasaka, M. Hyaluronan oligosaccharides induce CD44 cleavage and promote cell migration in CD44-expressing tumor cells. J. Biol. Chem. 2003, 278, 32259–32265. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Ishikawa, O.; Okada, K.; Kojima, Y.; Igarashi, Y.; Miyachi, Y. Disaccharide analysis of human skin glycosaminoglycans in sun-exposed and sun-protected skin of aged people. J. Dermatol. Sci. 1996, 11, 129–133. [Google Scholar] [CrossRef]
- Tzellos, T.G.; Klagas, I.; Vahtsevanos, K.; Triaridis, S.; Printza, A.; Kyrgidis, A.; Karakiulakis, G.; Zouboulis, C.C.; Papakonstantinou, E. Extrinsic ageing in the human skin is associated with alterations in the expression of hyaluronic acid and its metabolizing enzymes. Exp. Dermatol. 2009, 18, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, M.; Nakamura, M.; Luo, Y.; Morohashi, M. Ultrastructural characterization of microvasculature in photoaging. J. Dermatol. Sci. 2001, 27 (Suppl. 1), 32–41. [Google Scholar] [CrossRef]
- Trompezinski, S.; Pernet, I.; Schmitt, D.; Viac, J. UV radiation and prostaglandin E2 up-regulate vascular endothelial growth factor (VEGF) in cultured human fibroblasts. Inflamm. Res. 2001, 50, 422–427. [Google Scholar] [CrossRef]
- Manuskiatti, W.; Maibach, H.I. Hyaluronic acid and skin: Wound healing and aging. Int. J. Dermatol. 1996, 35, 539–544. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinol 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Stern, R.; Kogan, G.; Jedrzejas, M.J.; Soltes, L. The many ways to cleave hyaluronan. Biotechnol. Adv. 2007, 25, 537–557. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Tsukahara, K.; Fujimura, T.; Yoshida, Y.; Kitahara, T.; Hotta, M.; Moriwaki, S.; Witt, P.S.; Simion, F.A.; Takema, Y. Comparison of age-related changes in wrinkling and sagging of the skin in Caucasian females and in Japanese females. J. Cosmet. Sci. 2004, 55, 351–371. [Google Scholar] [CrossRef]
- Nouveau-Richard, S.; Yang, Z.; Mac-Mary, S.; Li, L.; Bastien, P.; Tardy, I.; Bouillon, C.; Humbert, P.; de Lacharriere, O. Skin ageing: A comparison between Chinese and European populations. A pilot study. J. Dermatol. Sci. 2005, 40, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.H. Photoaging in Asians. Photodermatol. Photoimmunol. Photomed. 2003, 19, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.V. Ethnic skin types: Are there differences in skin structure and function? Int. J. Cosmet. Sci. 2006, 28, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A cutaneous perspective on its production, metabolism, and functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kleszczynski, K.; Semak, I.; Janjetovic, Z.; Zmijewski, M.A.; Kim, T.K.; Slominski, R.M.; Reiter, R.J.; Fischer, T.W. Local melatoninergic system as the protector of skin integrity. Int. J. Mol. Sci. 2014, 15, 17705–17732. [Google Scholar] [CrossRef]
- Chaiprasongsuk, A.; Janjetovic, Z.; Kim, T.K.; Jarrett, S.G.; D’Orazio, J.A.; Holick, M.F.; Tang, E.K.Y.; Tuckey, R.C.; Panich, U.; Li, W.; et al. Protective effects of novel derivatives of vitamin D3 and lumisterol against UVB-induced damage in human keratinocytes involve activation of Nrf2 and p53 defense mechanisms. Redox Biol. 2019, 24, 101206. [Google Scholar] [CrossRef]
- Slominski, A.T.; Janjetovic, Z.; Kim, T.K.; Wasilewski, P.; Rosas, S.; Hanna, S.; Sayre, R.M.; Dowdy, J.C.; Li, W.; Tuckey, R.C. Novel non-calcemic secosteroids that are produced by human epidermal keratinocytes protect against solar radiation. J. Steroid Biochem. Mol. Biol. 2015, 148, 52–63. [Google Scholar] [CrossRef]
- Galamb, O.; Spisak, S.; Sipos, F.; Toth, K.; Solymosi, N.; Wichmann, B.; Krenacs, T.; Valcz, G.; Tulassay, Z.; Molnar, B. Reversal of gene expression changes in the colorectal normal-adenoma pathway by NS398 selective COX2 inhibitor. Br. J. Cancer 2010, 102, 765–773. [Google Scholar] [CrossRef]
- Yoshida, H.; Yamazaki, K.; Komiya, A.; Aoki, M.; Kasamatsu, S.; Murata, T.; Sayo, T.; Cilek, M.Z.; Okada, Y.; Takahashi, Y. Inhibitory effects of Sanguisorba officinalis root extract on HYBID (KIAA1199)-mediated hyaluronan degradation and skin wrinkling. Int. J. Cosmet. Sci. 2019, 41, 12–20. [Google Scholar] [CrossRef]
- Yoshida, H.; Yamazaki, K.; Komiya, A.; Aoki, M.; Nakamura, T.; Kasamatsu, S.; Murata, T.; Sayo, T.; Okada, Y.; Takahashi, Y. Inhibition of HYBID (KIAA1199)-mediated hyaluronan degradation and anti-wrinkle effect of Geranium thunbergii extract. J. Cosmet. Dermatol. 2019, 18, 1052–1060. [Google Scholar] [CrossRef]
- Fujimura, T.; Haketa, K.; Hotta, M.; Kitahara, T. Loss of skin elasticity precedes to rapid increase of wrinkle levels. J. Dermatol. Sci. 2007, 47, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kwon, S.H.; Huh, C.H.; Park, K.C.; Youn, S.W. The influences of skin visco-elasticity, hydration level and aging on the formation of wrinkles: A comprehensive and objective approach. Skin Res. Technol. 2013, 19, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Akazaki, S.; Nakagawa, H.; Kazama, H.; Osanai, O.; Kawai, M.; Takema, Y.; Imokawa, G. Age-related changes in skin wrinkles assessed by a novel three-dimensional morphometric analysis. Br. J. Dermatol. 2002, 147, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Averbeck, M.; Gebhardt, C.; Anderegg, U.; Simon, J.C. Suppression of hyaluronan synthase 2 expression reflects the atrophogenic potential of glucocorticoids. Exp. Dermatol. 2010, 19, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, C.; Averbeck, M.; Diedenhofen, N.; Willenberg, A.; Anderegg, U.; Sleeman, J.P.; Simon, J.C. Dermal hyaluronan is rapidly reduced by topical treatment with glucocorticoids. J. Investig. Dermatol. 2010, 130, 141–149. [Google Scholar] [CrossRef]
- Norsgaard, H.; Kurdykowski, S.; Descargues, P.; Gonzalez, T.; Marstrand, T.; Dunstl, G.; Ropke, M. Calcipotriol counteracts betamethasone-induced decrease in extracellular matrix components related to skin atrophy. Arch. Dermatol. Res. 2014, 306, 719–729. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, H.; Okada, Y. Role of HYBID (Hyaluronan Binding Protein Involved in Hyaluronan Depolymerization), Alias KIAA1199/CEMIP, in Hyaluronan Degradation in Normal and Photoaged Skin. Int. J. Mol. Sci. 2019, 20, 5804. https://doi.org/10.3390/ijms20225804
Yoshida H, Okada Y. Role of HYBID (Hyaluronan Binding Protein Involved in Hyaluronan Depolymerization), Alias KIAA1199/CEMIP, in Hyaluronan Degradation in Normal and Photoaged Skin. International Journal of Molecular Sciences. 2019; 20(22):5804. https://doi.org/10.3390/ijms20225804
Chicago/Turabian StyleYoshida, Hiroyuki, and Yasunori Okada. 2019. "Role of HYBID (Hyaluronan Binding Protein Involved in Hyaluronan Depolymerization), Alias KIAA1199/CEMIP, in Hyaluronan Degradation in Normal and Photoaged Skin" International Journal of Molecular Sciences 20, no. 22: 5804. https://doi.org/10.3390/ijms20225804
APA StyleYoshida, H., & Okada, Y. (2019). Role of HYBID (Hyaluronan Binding Protein Involved in Hyaluronan Depolymerization), Alias KIAA1199/CEMIP, in Hyaluronan Degradation in Normal and Photoaged Skin. International Journal of Molecular Sciences, 20(22), 5804. https://doi.org/10.3390/ijms20225804