Continentalic Acid Rather Than Kaurenoic Acid Is Responsible for the Anti-Arthritic Activity of Manchurian Spikenard In Vitro and In Vivo


 ,
,  ,
,
Abstract

1. Introduction
2. Results and Discussion
2.1. Optimization of the Ethanol Content of the Extraction Solvents in RAW264.7 Cells
2.2. The 50% Ethanolic Extract of Manchurian Spikenard Inhibits Interleukin (IL)-1β-Stimulated Production of Inflammatory Mediators in Human Chondrocytes
2.3. The 50% Ethanolic Extract of Manchurian Spikenard Inhibits IL-1β-Stimulated Expression of Matrix Metalloproteinases (MMPs) in Human Chondrocytes
2.4. Continentalic Acid, Not Kaurenoic Acid, is Responsible for the In Vitro Anti-Arthritic Activity of Manchurian Spikenard
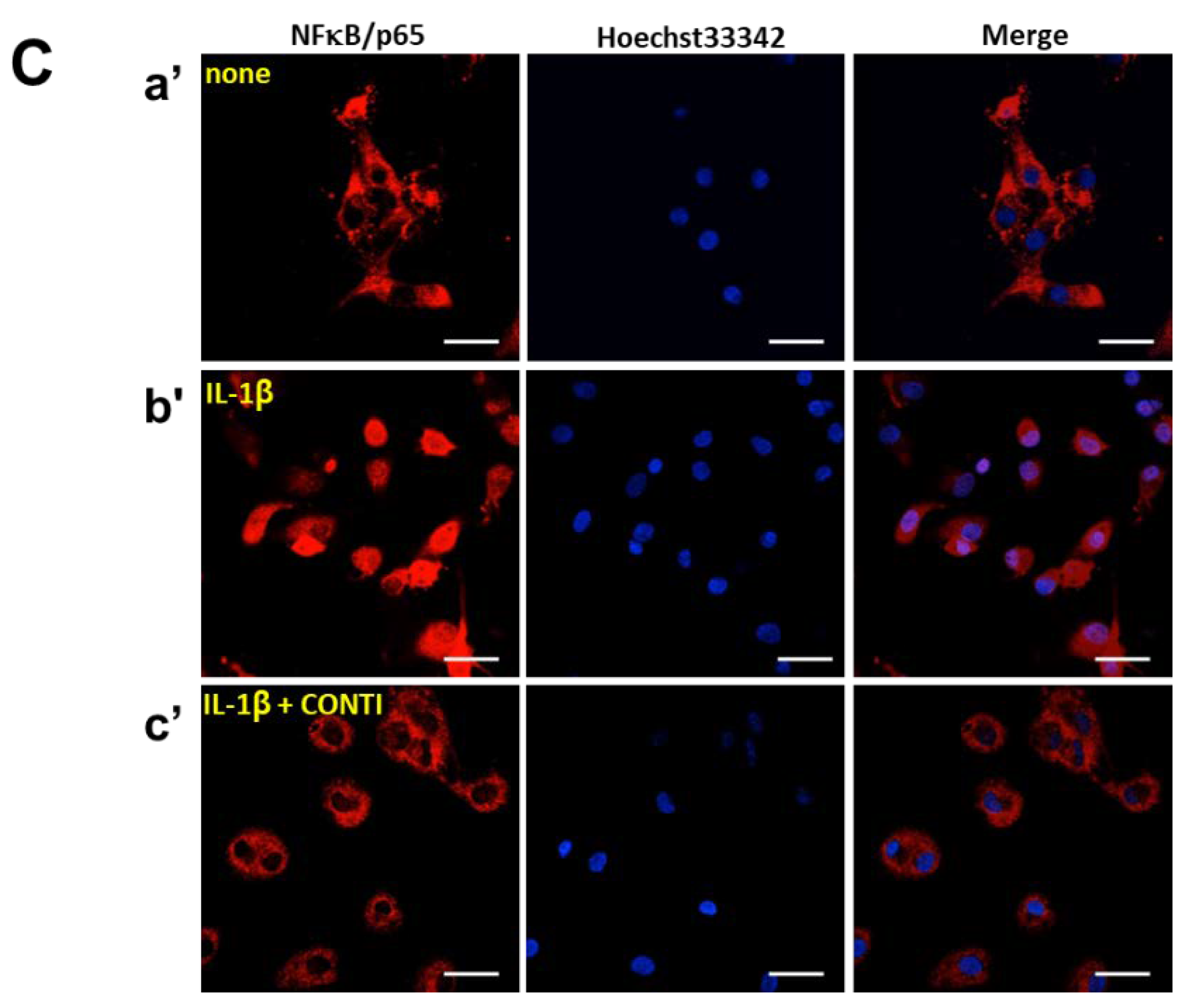
2.5. Continentalic Acid in the Spikenard Extract Inhibits IL-1β-Induced Phosphorylation of Extracellular Signal-Regulated Kinase/ Jun Amino-Terminal Kinase/p38 Mitogen-Activated Protein (ERK/JNK/p38 MAP) Kinases and Nuclear Translocation of the NF-κB/p65 Subunit
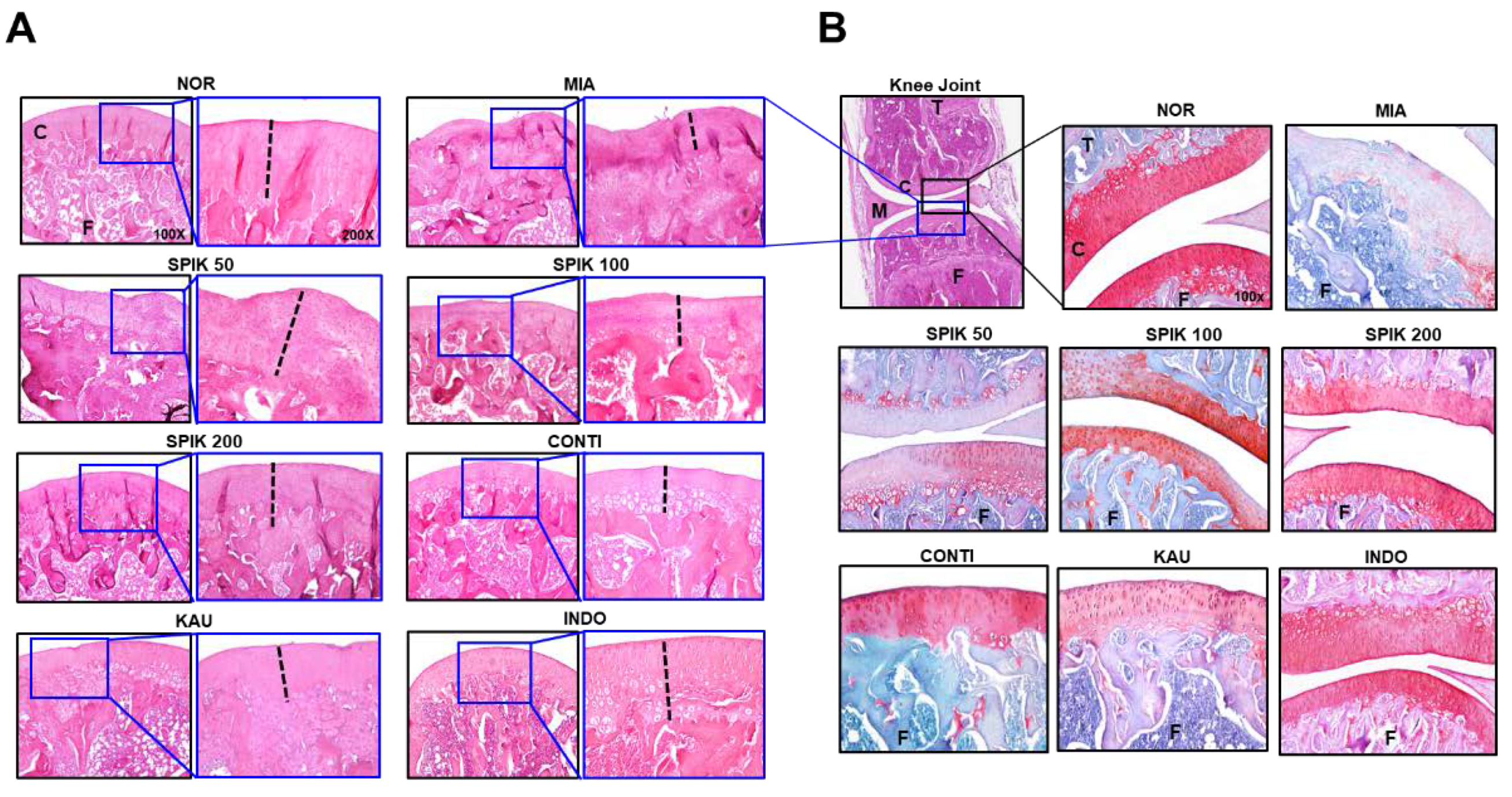
2.6. The Spikenard Extract and Continentalic Acid Alleviates Monoiodoacetate (MIA)-Induced Osteoarthritis in Rats
3. Materials and Methods
3.1. Cell Experiments
3.1.1. Plant Material and Chemicals
3.1.2. Cell Culture
3.1.3. Reverse Transcription Polymerase Chain Reaction (RT−PCR) and Enzyme-Linked Immunosorbent Assay (ELISA)
3.1.4. Western Hybridization
3.1.5. Immunofluorescence Microscopy
3.2. Animal Experiments
3.2.1. Animals and Groups
3.2.2. Monoiodoacetate(MIA)-Induced Knee Arthritis
3.2.3. Evaluation of Arthritic Symptoms
3.2.4. Histology
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldring:, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Herbalogy; Young Lim Sa: Seoul, Korea, 2000. [Google Scholar]
- Huh, J. Dong Ui Bo Gam: The precious mirror of Oriental Medicine, 2nd ed.; Beob In Mun Hwa Sa: Seoul, Korea, 2012. [Google Scholar]
- Okuyama, E.; Nishimura, S.; Yamazaki, M. Analgesic principles from Aralia cordata Thunb. Chem. Pharm. Bull. 1991, 39, 405–407. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lim, H.; Jung, H.A.; Choi, J.S.; Kim, Y.S.; Kang, S.S.; Kim, H.P. Anti-inflammatory activity of the constituents of the roots of Aralia continentalis. Arch. Pharm. Res. 2009, 32, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Hung, T.M.; Min, B.S.; Lee, I.S.; Na, M.K.; Woo, M.H.; Son, J.K.; Kim, Y.H.; Choi, J.S.; Bae, K.H. Quantitative determination of diterpenoids from the roots of Aralia cordata. Nat. Prod. Sci. 2009, 15, 50–54. [Google Scholar]
- Hong, R.; Sur, B.; Yeom, M.; Lee, B.; Kim, K.S.; Rodriguez, J.P.; Lee, S.; Kang, K.S.; Huh, C.K.; Lee, S.C.; et al. Anti-inflammatory and anti-arthritic effects of the ethanolic extract of Aralia continentalis Kitag. in IL-1β-stimulated human fibroblast-like synoviocytes and rodent models of polyarthritis and nociception. Phytomedicine 2018, 38, 45–56. [Google Scholar] [PubMed]
- Dunham, J.; Hoedt-Schmidt, S.; Kalbhen, D.A. Prolonged effect of iodoacetate on articular cartilage and its modification by an anti-rheumatic drug. Int. J. Exp. Pathol. 1993, 74, 283–289. [Google Scholar]
- Murphy, G.; Knäuper, V.; Atkinson, S.; Butler, G.; English, W.; Hutton, M.; Stracke, J.; Clark, I. Matrix metalloproteinases in arthritic disease. Arthritis Res. Ther. 2002, 4, S39–S49. [Google Scholar] [CrossRef]
- Jabłonska-Trypuc, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzyme Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef]
- Vincenti, M.P.; Brinckerhoff, C.E. Transcriptional regulation of collagenase (MMP-1, MMP-13) genes in arthritis: Integration of complex signaling pathways for the recruitment of gene-specific transcription factors. Arthritis Res. 2002, 4, 157–164. [Google Scholar] [CrossRef]
- Bang, J.S.; Oh, D.H.; Choi, H.M.; Sur, B.J.; Lim, S.J.; Kim, J.Y.; Yang, H.I.; Yoo, M.C.; Hahm, D.H.; et al. Anti-inflammatory and antiarthritic effects of piperine in human interleukin 1β-stimulated fibroblast-like synoviocytes and in rat arthritis models. Arthritis Res. Ther. 2009, 11, R49. [Google Scholar] [CrossRef]
- Kim, K.S.; Choi, H.M.; Oh, D.H.; Kim, C.; Jeong, J.S.; Yoo, M.C.; Yang, H.I. Effect of taurine chloramine on the production of matrix metalloproteinases (MMPs) in adiponectin- or IL-1β-stimulated fibroblast-like synoviocytes. J. Biomed. Sci. 2010, 17, S27. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Huang, J.F.; Du, W.X.; Tong, P.J. Expression and significance of MMP3 in synovium of knee joint at different stage in osteoarthritis patients. Asian Pac. J. Trop. Med. 2014, 7, 297–300. [Google Scholar] [CrossRef]
- Uemura, Y.; Hayashi, H.; Takahashi, T.; Saitho, T.; Umeda, R.; Ichise, Y.; Sendo, S.; Tsuji, G.; Kumagai, S. MMP-3 as a biomarker of disease activity of rheumatoid arthritis. Rinsho Byori. 2015, 63, 1357–1364. [Google Scholar] [PubMed]
- Green, M.J.; Gough, A.K.; Devlin, J.; Smith, J.; Astin, P.; Taylor, D.; Emery, P. Serum MMP-3 and MMP-1 and progression of joint damage in early rheumatoid arthritis. Rheumatology 2003, 42, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Peeters-Joris, C.; Hammani, K.; Singer, C.F. Differential regulation of MMP-13 (collagenase-3) and MMP-3 (stromelysin-1) in mouse calvariae. Biochim. Biophys. Acta. 1998, 1405, 14–28. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, E.B.; Kwon, Y.E.; Lee, J.J.; Cho, W.S.; Kim, H.A.; Song, Y.W. Effect of estrogen on the expression of matrix metalloproteinase (MMP)-1, MMP-3, and MMP-13 and tissue inhibitor of metalloproternase-1 in osteoarthritis chondrocytes. Rheumatol. Int. 2003, 23, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.H.; Huh, J.E.; Lee, J.D.; Choi, D.Y.; Park, D.S. Effect of Aralia cordata extracts on cartilage protection and apoptosis inhibition. Biol. Pharm. Bull. 2006, 29, 1423–1430. [Google Scholar] [CrossRef]
- Park, D.S.; Huh, J.E.; Baek, Y.H. Therapeutic effect of Aralia cordata extracts on cartilage protection in collagenase-induced inflammatory arthritis rabbit model. J. Ethnopharmacol. 2009, 125, 207–217. [Google Scholar] [CrossRef]
- Tsakiri, N.; Kimber, I.; Rothwell, N.J.; Pinteaux, E. Differential effects of interleukin-1α and β on interleukin-6 and chemokine synthesis in neurones. Mol. Cell Neurosci. 2008, 38, 259–265. [Google Scholar] [CrossRef]
- Perry, L.M. Medicinal plants of East and Southeast Asia: Attributed Properties and Uses; MIT Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Kim, K.H.; Sadikot, R.T.; Joo, M. Therapeutic effect of ent-kaur-16-en-19-oic acid on neutrophilic lung inflammation and sepsis is mediated by Nrf2. Biochem. Biophys. Res. Commun. 2016, 474, 534–540. [Google Scholar] [CrossRef]
- Lyu, J.H.; Lee, G.S.; Kim, K.H.; Kim, H.W.; Cho, S.I.; Jeong, S.I.; Kim, H.J.; Ju, Y.S.; Kim, H.K.; Sadikot, R.T.; et al. ent-kaur-16-en-19-oic Acid, isolated from the roots of Aralia continentalis, induces activation of Nrf2. J. Ethnopharmacol. 2011, 137, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.I.; Han, W.S.; Yun, Y.H.; Kim, K.J. Continentalic acid from Aralia continentalis shows activity against methicillin-resistant Staphylococcus aureus. Phytother. Res. 2006, 20, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.O.; Jeong, S.I.; Kwon, J.W.; Kim, Y.C.; Jang, S.I. Continentalic acid from Aralia continentalis induces growth inhibition and apoptosis in HepG2 cells. Arch. Pharm. Res. 2008, 31, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Arora, B.S.; Khajuria, A.; Sidiq, T.; Kishore, D.; Vishwakarma, R.A. Isolation of continentalic acid from Aralia cachemirica and its immune-biological evaluation. Int. J. Pharm. Sci. Res. 2011, 2, 2183–2189. [Google Scholar]
- Villa-Ruano, N.; Lozoya-Gloria, E.; Pacheco-Hernández, Y. Chapter 3—Kaurenoic acid: A diterpene with a wide range of biological activities. Stud. Nat. Products Chem. 2016, 51, 151–174. [Google Scholar]
- Loesera, R.F.; Ericksona, E.A.; Longa, D.L. Mitogen-activated protein kinases as therapeutic targets in osteoarthritis. Curr. Opin. Rheumatol. 2008, 20, 581–586. [Google Scholar] [CrossRef]
- Guzman, R.E.; Evans, M.G.; Bove, S.; Morenko, B.; Kilgore, K. Monoiodoacetate-induced histologic changes in subchondral bone and articular cartilage of rat femorotibial joints: An animal model of osteoarthritis. Toxicol. Pathol. 2003, 31, 619–624. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (bp) | Nucleotide Sequence | Annealing Temp. (°C)/Cycles | ||
|---|---|---|---|---|
| human | ||||
| GAPDH (579) | sense | 5′-ATC CCA TCA CCA TCT TCC AG-3′ | 58/30 | |
| antisense | 5′-CCT GCT TCA CCA CCT TCT TG-3′ | |||
| IL-6 (150) | sense | 5′-AGT TGC CTT CTT GGG ACT GA -3′ | 55/31 | |
| antisense | 5′-TCC ACG ATT TCC CAG AGA AC -3′ | |||
| IL-8 (166) | sense | 5′-GTT TTG CCA AGG AGT GCT AA -3′ | 55/31 | |
| antisense | 5′-CCA GAC AGA GCT CTC TTC CA -3′ | |||
| COX-2 (158) | sense | 5′-TGA GCA TCT ACG GTT TGC TG -3′ | 55/31 | |
| antisense | 5′-TGC TTG TCT GGA ACA ACT GC -3′ | |||
| MMP-1 (388) | sense | 5′-CCT AGC TAC ACC TTC AGT GG-3′ | 57/29 | |
| antisense | 5′-GCC CAG TAC TTA TTC CCT TT-3′ | |||
| MMP-3 (365) | sense | 5′-TCC CCC TGA CTC CCC TGA-3′ | 57/29 | |
| antisense | 5′-TCC TCA CGG TTG GAG GGA AA-3′ ′ | |||
| MMP-13 (150) | sense | 5′-TGA CCC TTC CTT ATC CCT TG-3′ | 57/29 | |
| antisense | 5′-ATA CGG TTG GGA AGT TCT GG-3′ | |||
| iNOS (320) | sense | 5′-GCA TGT ACC CTC GGT TCT GT-3′ | 58/30 | |
| antisense | 5′-CAT GGT GAA CAC GTT CTT GG-3′ | |||
| mouse | ||||
| GAPDH (223) | sense | 5′-AAC TTT GGC ATT GTG GAA GG-3′ | 58/30 | |
| antisense | 5′-ACA CAT TGG GGG TAG GAA CA-3′ | |||
| IL-6 (159) | sense | 5′-AGT TGC CTT CTT GGG ACT GA-3′ | 52/27 | |
| antisense | 5′-TCC ACG ATT TCC CAG AGA AC-3′ | |||
| COX-2 (194) | sense | 5′-AGA AGG AAA TGG CTG CAGAA 3′ | 55/28 | |
| antisense | 5′-GCT CGG CTT CCA GTA TTG AG -3′ | |||
| iNOS (199) | sense | 5′-CCT CCT CCA CCC TAC CAA GT-3′ | 57/28 | |
| antisense | 5′-CAC CCA AAG TGC TTC AGT CA-3′ | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, R.; Kim, K.S.; Choi, G.M.; Yeom, M.; Lee, B.; Lee, S.; Kang, K.S.; Lee, H.S.; Park, H.-J.; Hahm, D.-H. Continentalic Acid Rather Than Kaurenoic Acid Is Responsible for the Anti-Arthritic Activity of Manchurian Spikenard In Vitro and In Vivo. Int. J. Mol. Sci. 2019, 20, 5488. https://doi.org/10.3390/ijms20215488
Hong R, Kim KS, Choi GM, Yeom M, Lee B, Lee S, Kang KS, Lee HS, Park H-J, Hahm D-H. Continentalic Acid Rather Than Kaurenoic Acid Is Responsible for the Anti-Arthritic Activity of Manchurian Spikenard In Vitro and In Vivo. International Journal of Molecular Sciences. 2019; 20(21):5488. https://doi.org/10.3390/ijms20215488
Chicago/Turabian StyleHong, Riwon, Kyoung Soo Kim, Gwang Muk Choi, Mijung Yeom, Bombi Lee, Sanghyun Lee, Ki Sung Kang, Hyang Sook Lee, Hi-Joon Park, and Dae-Hyun Hahm. 2019. "Continentalic Acid Rather Than Kaurenoic Acid Is Responsible for the Anti-Arthritic Activity of Manchurian Spikenard In Vitro and In Vivo" International Journal of Molecular Sciences 20, no. 21: 5488. https://doi.org/10.3390/ijms20215488
APA StyleHong, R., Kim, K. S., Choi, G. M., Yeom, M., Lee, B., Lee, S., Kang, K. S., Lee, H. S., Park, H.-J., & Hahm, D.-H. (2019). Continentalic Acid Rather Than Kaurenoic Acid Is Responsible for the Anti-Arthritic Activity of Manchurian Spikenard In Vitro and In Vivo. International Journal of Molecular Sciences, 20(21), 5488. https://doi.org/10.3390/ijms20215488