Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber)


 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
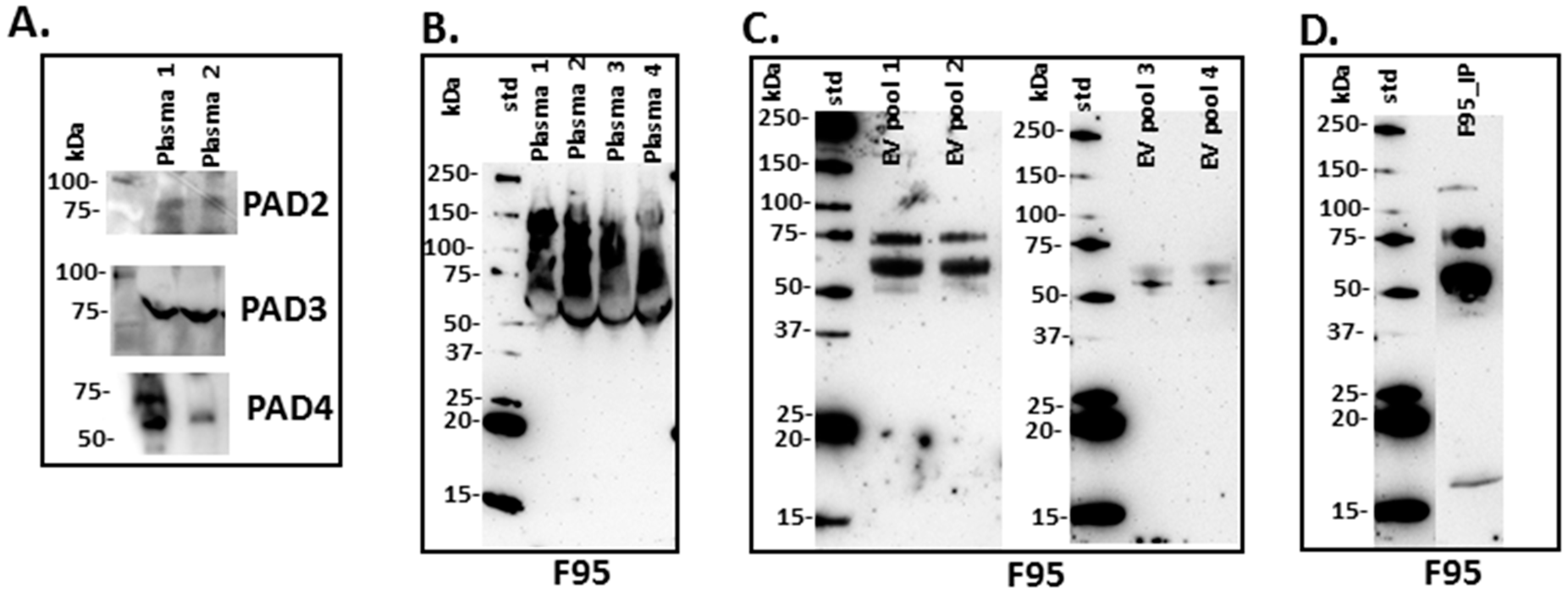
2.1. PAD Homologues in Naked Mole-Rat Plasma
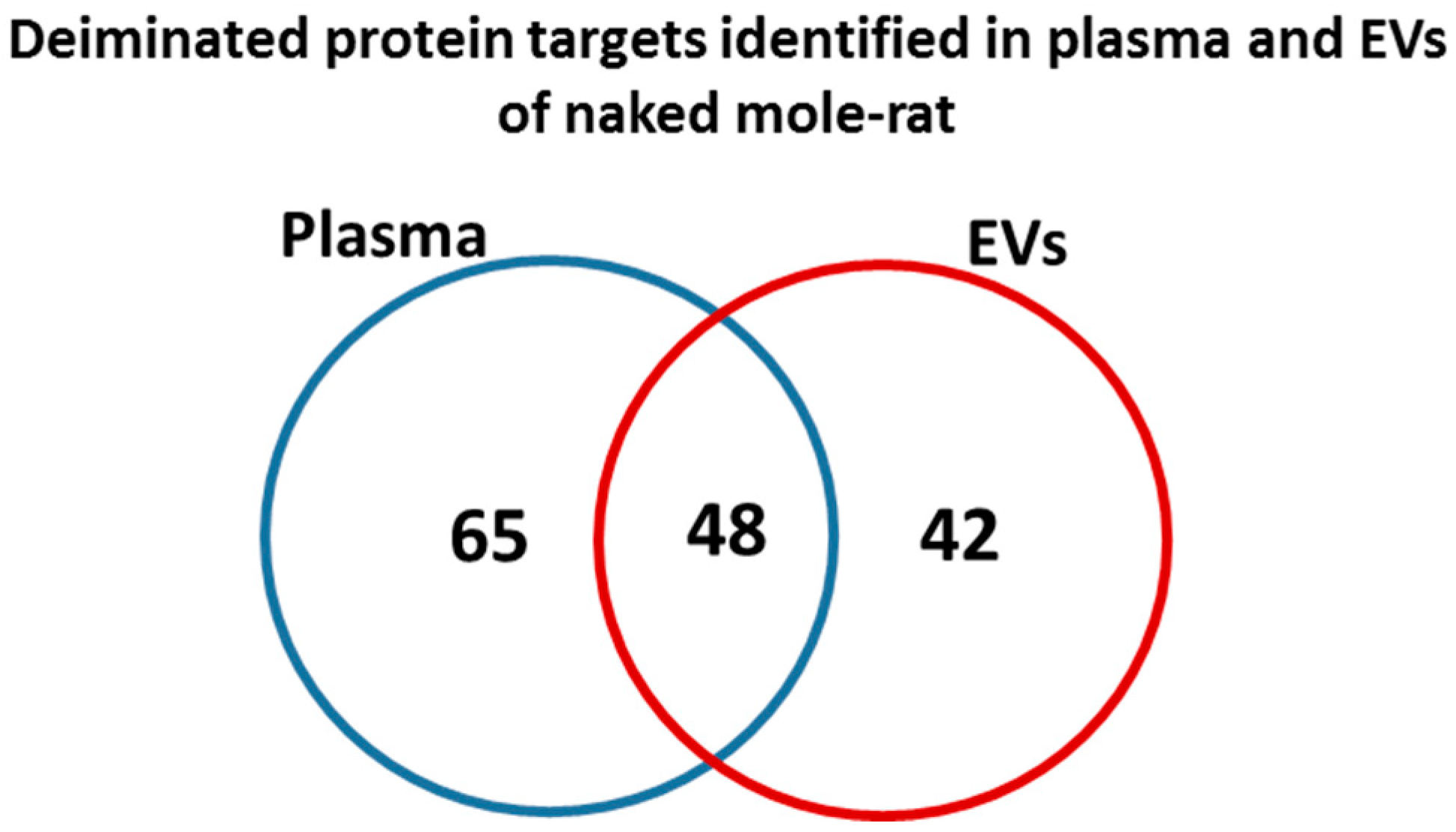
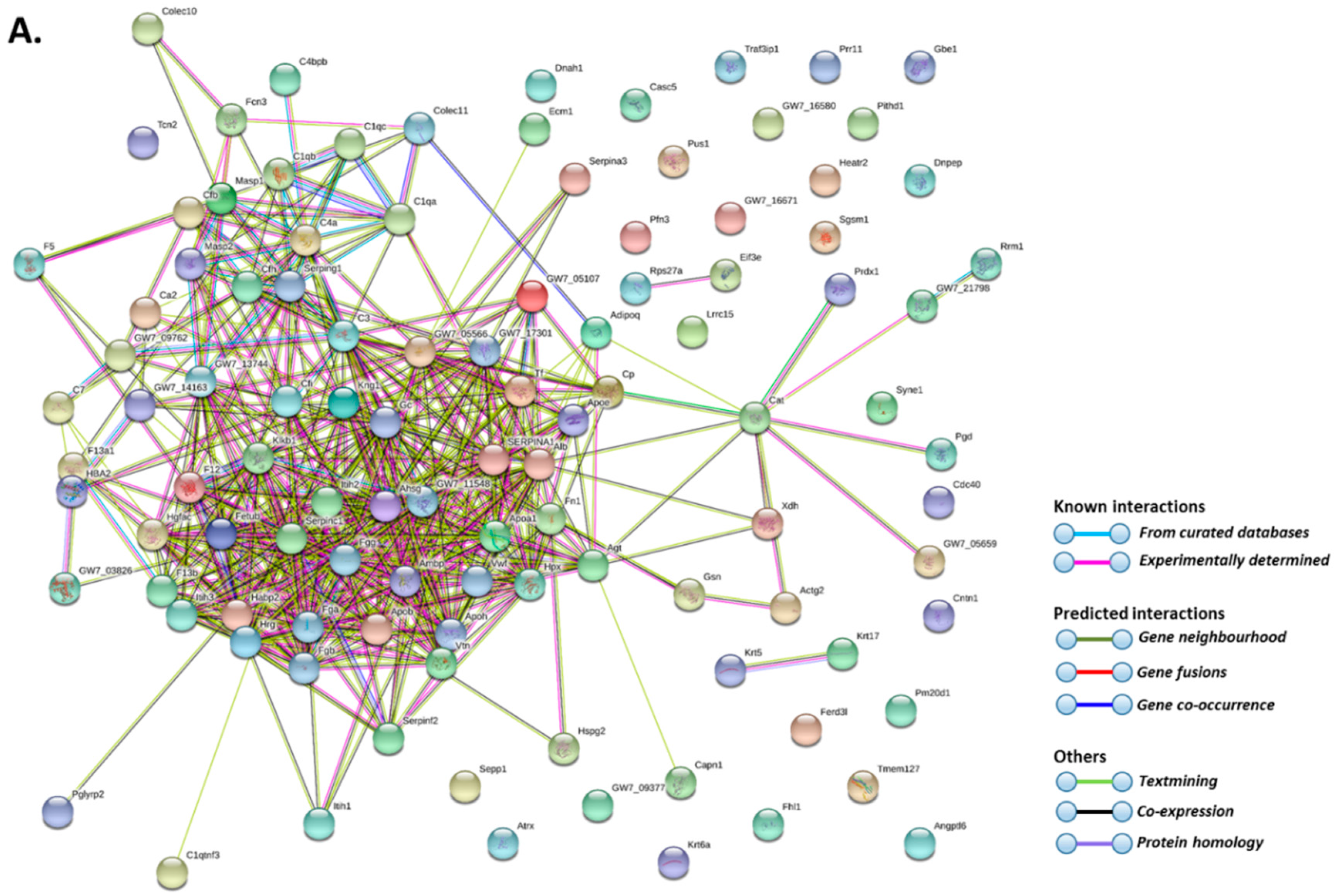
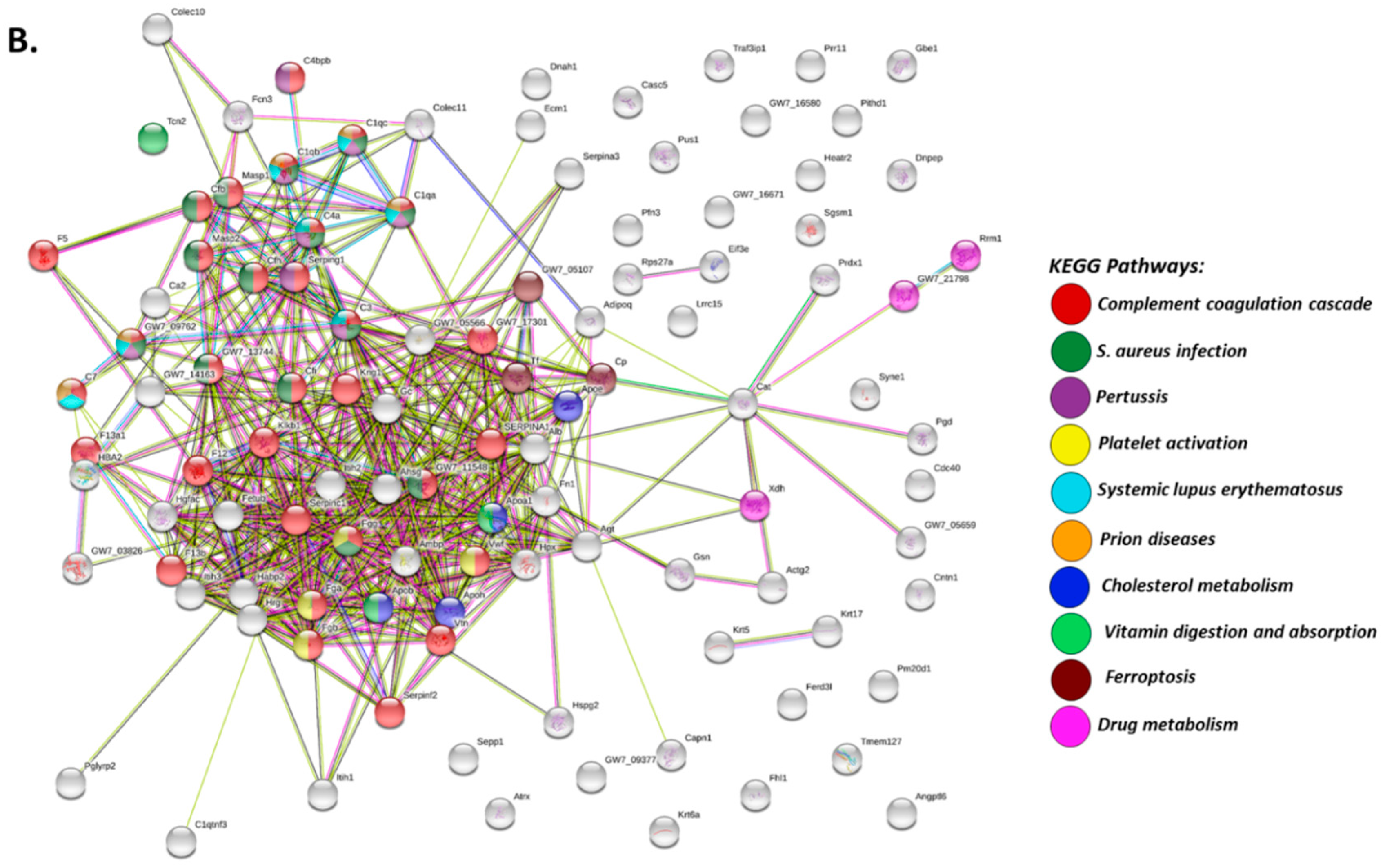
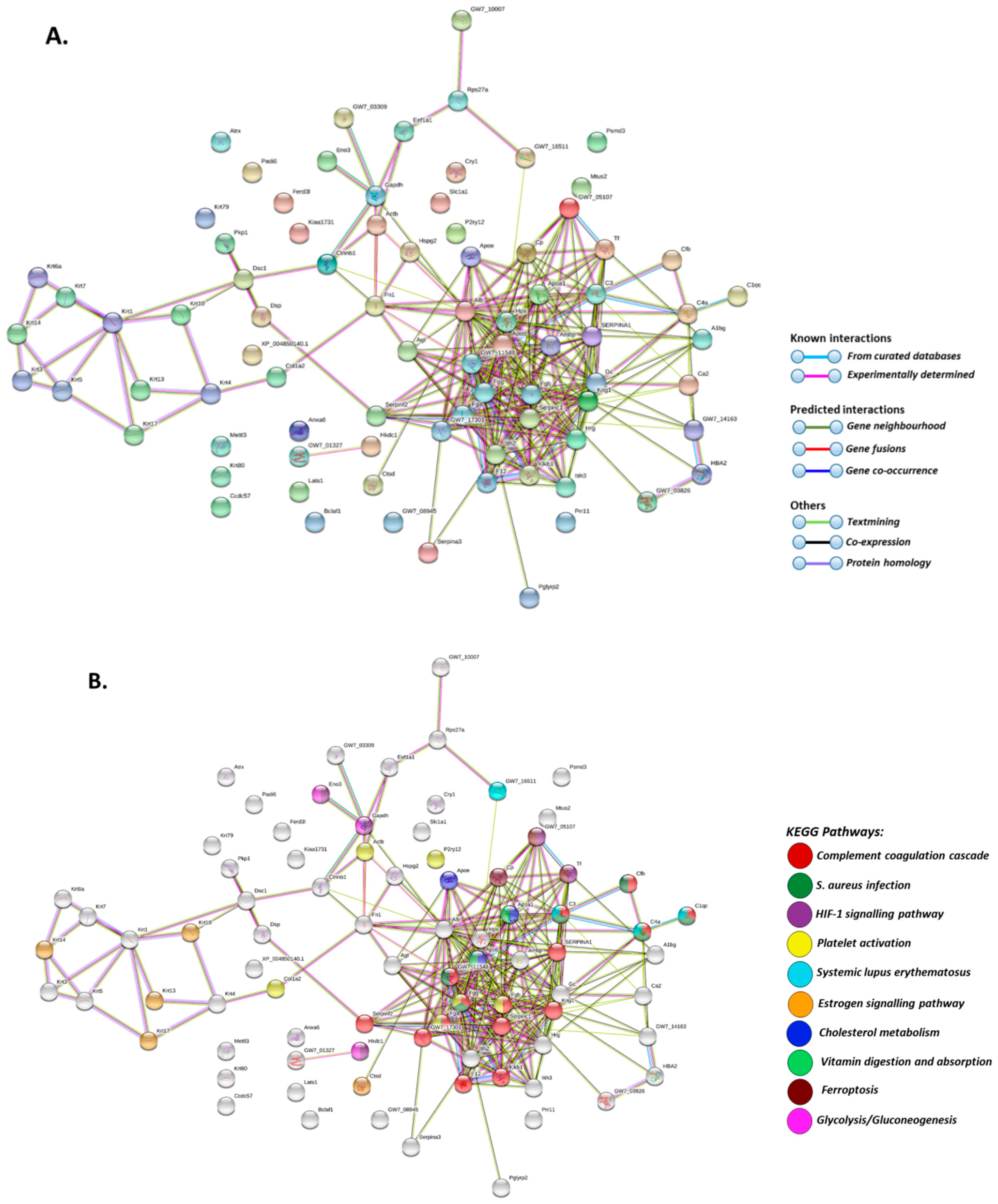
2.2. Deiminated Protein Profiles of Naked Mole-Rat Plasma and Plasma-Derived EVs
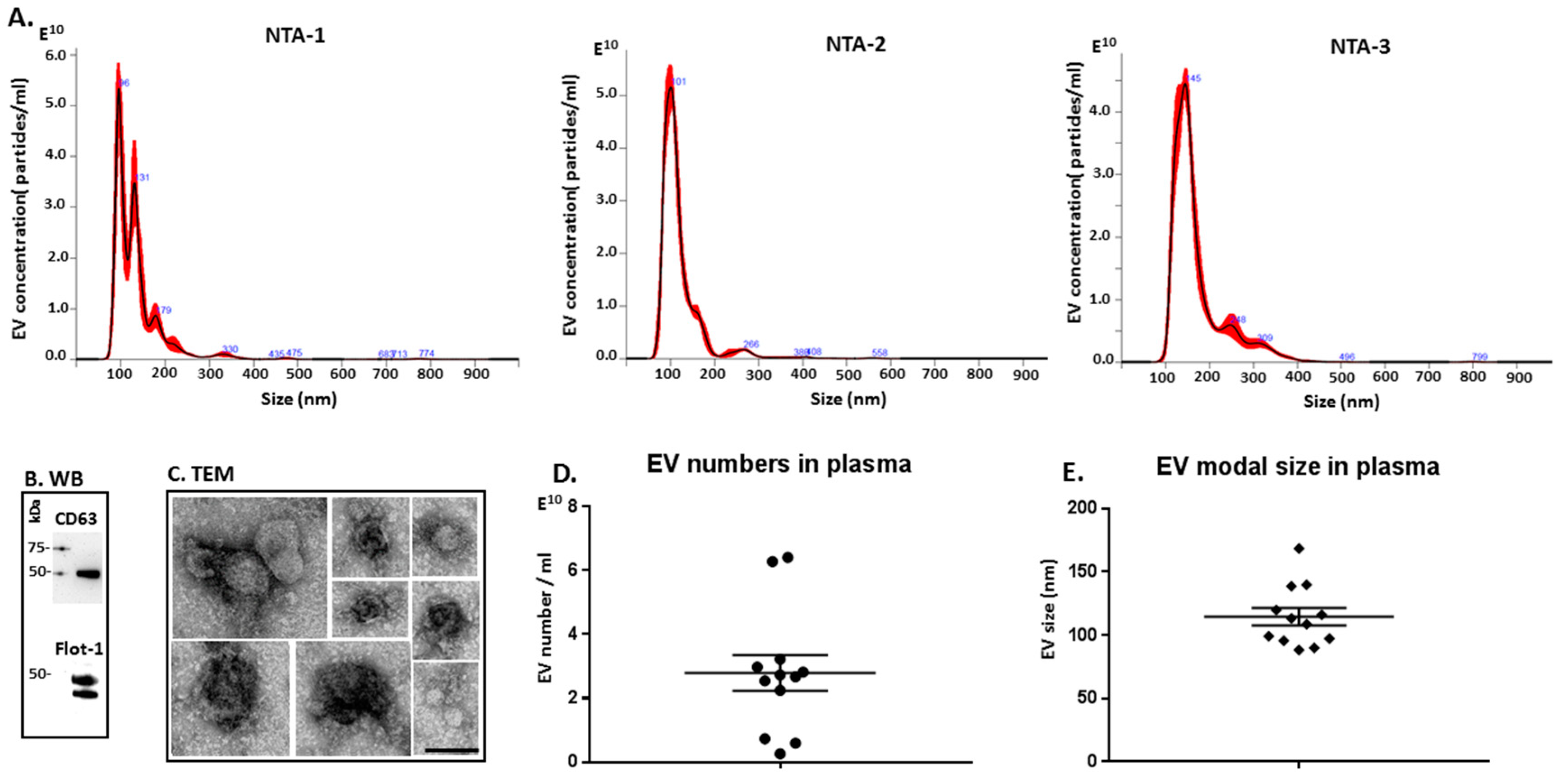
2.3. Characterisation of Extracellular Vesicles in Naked Mole-Rat Plasma
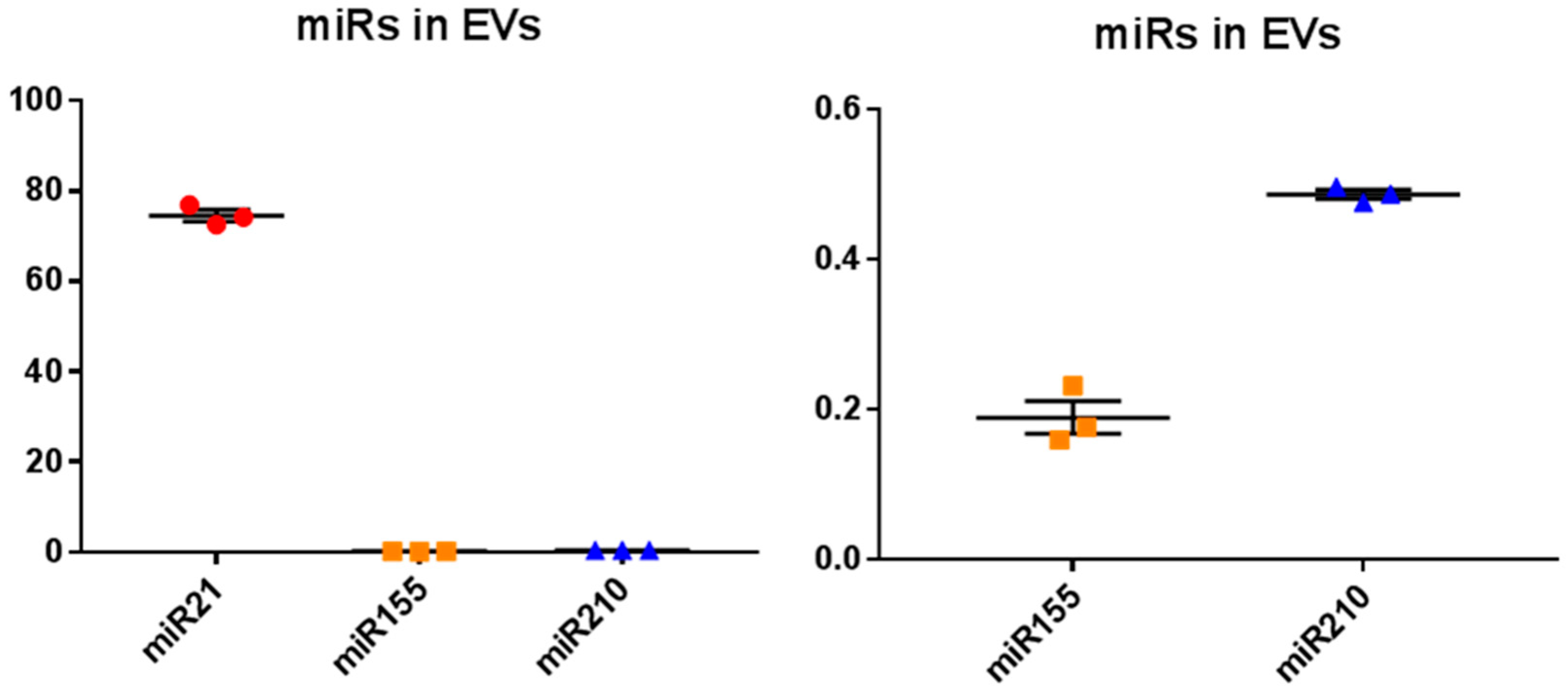
2.4. MicroRNA Analysis of Naked Mole-Rat EVs
3. Discussion
4. Materials and Methods
4.1. Sampling of Naked Mole-Rat Plasma
4.2. Extracellular Vesicle Isolation and Nanoparticle Tracking Analysis (NTA)
4.3. Transmission Electron Microscopy (TEM)
4.4. Western Blotting Analysis
4.5. Immunoprecipitation and Protein Identification
4.6. MicroRNA Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine Serum Albumin |
| CD63 | CD63 antigen; granulophysin; lysosomal-associated membrane protein 3 |
| CNS | Central Nervous System |
| ECL | Enhanced Chemiluminescence |
| EVs | Extracellular Vesicles |
| F95 | Pan-deimination/citrullination antibody |
| FBS | Foetal Bovine Serum |
| Flot-1 | Flotillin-1 |
| HIF | Hypoxia-inducible Factor |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| kDa | Kilodalton |
| LC-MS/MS | Liquid Chromatography Mass Spectrometry |
| miR | microRNA |
| NTA | Nanoparticle Tracking Analysis |
| PAD | Peptidylarginine Deiminase |
| SDS-PAGE | Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis |
| TBS | Tris Buffered Saline |
| TEM | Transmission Electron Microscopy |
| WB | Western Blotting |
References
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. Bioessays 2003, 25, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Toth, E.; Tarcsa, E.; Falus, A.; Buzas, E.I. Citrullination: A posttranslational modification in health and disease. Int. J. Biochem. Cell Biol. 2006, 38, 1662–1677. [Google Scholar] [CrossRef] [PubMed]
- Bicker, K.L.; Thompson, P.R. The protein arginine deiminases: Structure, function, inhibition, and disease. Biopolymers 2013, 99, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, Y. Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein Arginine Deiminases and Associated Citrullination: Physiological Functions and Diseases Associated with Dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Gallagher, M.; Kholia, S.; Kosgodage, U.S.; Hristova, M.; Hardy, J.; Inal, J.M. Peptidylarginine Deiminases-Roles in Cancer and Neurodegeneration and Possible Avenues for Therapeutic Intervention via Modulation of Exosome and Microvesicle (EMV) Release? Int. J. Mol. Sci. 2017, 18, 1196. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Martin, A.C. Protein moonlighting: A new factor in biology and medicine. Biochem. Soc. Trans. 2014, 42, 1671–1678. [Google Scholar] [CrossRef]
- Jeffrey, C.J. Protein moonlighting: What is it, and why is it important? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160523. [Google Scholar] [CrossRef]
- Mohanan, S.; Cherrington, B.D.; Horibata, S.; McElwee, J.L.; Thompson, P.R.; Coonrod, S.A. Potential role of peptidylarginine deiminase enzymes and protein citrullination in cancer pathogenesis. Biochem. Res. Int. 2012. [Google Scholar] [CrossRef]
- Wong, S.L.; Wagner, D.D. Peptidylarginine deiminase 4: A nuclear button triggering neutrophil extracellular traps in inflammatory diseases and aging. FASEB J. 2018, 32, 6358–6370. [Google Scholar] [CrossRef]
- Lange, S.; Gögel, S.; Leung, K.Y.; Vernay, B.; Nicholas, A.P.; Causey, C.P.; Thompson, P.R.; Greene, N.D.; Ferretti, P. Protein deiminases: New players in the developmentally regulated loss of neural regenerative ability. Dev. Biol. 2011, 355, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Rocha-Ferreira, E.; Thei, L.; Mawjee, P.; Bennett, K.; Thompson, P.R.; Subramanian, V.; Nicholas, A.P.; Peebles, D.; Hristova, M.; et al. Peptidylarginine deiminases: Novel drug targets for prevention of neuronal damage following hypoxic ischemic insult (HI) in neonates. J. Neurochem. 2014, 130, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Lange, S. Peptidylarginine Deiminases as Drug Targets in Neonatal Hypoxic-Ischemic Encephalopathy. Front. Neurol. 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Sase, T.; Arito, M.; Onodera, H.; Omoteyama, K.; Kurokawa, M.S.; Kagami, Y.; Ishigami, A.; Tanaka, Y.; Kato, T. Hypoxia-induced production of peptidylarginine deiminases and citrullinated proteins in malignant glioma cells. Biochem. Biophys. Res. Commun. 2017, 482, 50–56. [Google Scholar] [CrossRef]
- Yu, R.; Li, C.; Sun, L.; Jian, L.; Ma, Z.; Zhao, J.; Liu, X. Hypoxia induces production of citrullinated proteins in human fibroblast-like synoviocytes through regulating HIF1α. Scand. J. Immunol. 2018, 87, e12654. [Google Scholar] [CrossRef]
- Magnadottir, B.; Hayes, P.; Hristova, M.; Bragason, B.Þ.; Nicholas, A.P.; Dodds, A.W.; Gudmundsdottir, S.; Lange, S. Post-translational Protein Deimination in Cod (Gadus morhua L.) Ontogeny—Novel Roles in Tissue Remodelling and Mucosal Immune Defences? Dev. Comp. Immunol. 2018, 87, 157–170. [Google Scholar] [CrossRef]
- Magnadottir, B.; Hayes, P.; Gísladóttir, B.; Bragason, B.Þ.; Hristova, M.; Nicholas, A.P.; Guðmundsdóttir, S.; Lange, S. Pentraxins CRP-I and CRP-II are post-translationally deiminated and differ in tissue specificity in cod (Gadus morhua L.) ontogeny. Dev. Comp. Immunol. 2018, 87, 1–11. [Google Scholar] [CrossRef]
- Magnadottir, B.; Bragason, B.T.; Bricknell, I.R.; Bowden, T.; Nicholas, A.P.; Hristova, M.; Gudmundsdottir, S.; Dodds, A.W.; Lange, S. Peptidylarginine Deiminase and Deiminated Proteins are detected throughout Early Halibut Ontogeny—Complement Components C3 and C4 are post-translationally Deiminated in Halibut (Hippoglossus hippoglossus L.). Dev. Comp Immunol. 2019, 92, 1–19. [Google Scholar] [CrossRef]
- Kholia, S.; Jorfi, S.; Thompson, P.R.; Causey, C.P.; Nicholas, A.P.; Inal, J.; Lange, S. A Novel Role for Peptidylarginine Deiminases (PADs) in Microvesicle Release: A Therapeutic Potential for PAD Inhibitors to Sensitize Prostate Cancer Cells to Chemotherapy. J. Extracell. Vesicles 2015, 4, 26192. [Google Scholar] [CrossRef]
- Kosgodage, U.S.; Trindade, R.P.; Thompson, P.T.; Inal, J.M.; Lange, S. Chloramidine/Bisindolylmaleimide-I-Mediated Inhibition of Exosome and Microvesicle Release and Enhanced Efficacy of Cancer Chemotherapy. Int. J. Mol. Sci. 2017, 18, 1007. [Google Scholar] [CrossRef]
- Kosgodage, U.S.; Onganer, P.U.; Maclatchy, A.; Nicholas, A.P.; Inal, J.M.; Lange, S. Peptidylarginine Deiminases Post-translationally Deiminate Prohibitin and Modulate Extracellular Vesicle Release and miRNAs 21 and 126 in Glioblastoma Multiforme. Int. J. Mol. Sci. 2018, 20, 103. [Google Scholar] [CrossRef] [PubMed]
- Kosgodage, U.S.; Matewele, P.; Mastroianni, G.; Kraev, I.; Brotherton, D.; Awamaria, B.; Nicholas, A.P.; Lange, S.; Inal, J.M. Peptidylarginine Deiminase Inhibitors Reduce Bacterial Membrane Vesicle Release and Sensitize Bacteria to Antibiotic Treatment. Front. Cell. Infect. Microbiol. 2019, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Inal, J.M.; Ansa-Addo, E.A.; Lange, S. Interplay of host-pathogen microvesicles and their role in infectious disease. Biochem. Soc. Trans. 2013, 41, 258–262. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Drapkina, O.; Tonevitsky, A. Transcriptome of Extracellular Vesicles: State-of-the-Art. Front. Immunol. 2019, 10, 202. [Google Scholar] [CrossRef]
- Vagner, T.; Chin, A.; Mariscal, J.; Bannykh, S.; Engman, D.M.; Di Vizio, D. Protein Composition Reflects Extracellular Vesicle Heterogeneity. Proteomics 2019, 19, 1800167. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Ramirez, S.H.; Andrews, A.M.; Paul, D.; Pachter, J.S. Extracellular vesicles: Mediators and biomarkers of pathology along CNS barriers. Fluids Barriers CNS. 2018, 15, 19. [Google Scholar] [CrossRef]
- Iliev, D.; Strandskog, G.; Nepal, A.; Aspar, A.; Olsen, R.; Jørgensen, J.; Wolfson, D.; Ahluwalia, B.S.; Handzhiyski, J.; Mironova, R. Stimulation of exosome release by extracellular DNA is conserved across multiple cell types. FEBS J. 2018, 285, 3114–3133. [Google Scholar] [CrossRef]
- Magnadóttir, B.; Kraev, I.; Guðmundsdóttir, S.; Dodds, A.W.; Lange, S. Extracellular vesicles from cod (Gadus morhua L.) mucus contain innate immune factors and deiminated protein cargo. Dev. Comp. Immunol. 2019, 99, 103397. [Google Scholar] [CrossRef]
- Criscitiello, M.F.; Kraev, I.; Lange, S. Deiminated proteins in extracellular vesicles and plasma of nurse shark (Ginglymostoma cirratum)—Novel insights into shark immunity. Fish Shellfish Immunol. 2019, 92, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Kraev, I.; Magnadóttir, B.; Dodds, A.W. Complement component C4-like protein in Atlantic cod (Gadus morhua L.)—Detection in ontogeny and identification of post-translational deimination in serum and extracellular vesicles. Dev. Comp. Immunol. 2019, 101, 103437. [Google Scholar] [CrossRef] [PubMed]
- Rebl, A.; Köllner, B.; Anders, E.; Wimmers, K.; Goldammer, T. Peptidylarginine deiminase gene is differentially expressed in freshwater and brackish water rainbow trout. Mol. Biol. Rep. 2010, 37, 2333–2339. [Google Scholar] [CrossRef] [PubMed]
- Gavinho, B.; Rossi, I.V.; Evans-Osses, I.; Lange, S.; Ramirez, M.I. Peptidylarginine deiminase inhibition abolishes the production of large extracellular vesicles from Giardia intestinalis, affecting host-pathogen interactions by hindering adhesion to host cells. bioRxiv 2019. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZaid, A.A.; Yassin, A.M.; Ali, G.S.; Sitohy, M.Z. Biochemical characterization of peptidylarginine deiminase-like orthologs from thermotolerant Emericella dentata and Aspergillus nidulans. Enzyme Microb. Technol. 2019, 124, 41–53. [Google Scholar] [CrossRef]
- Woods, C.A.; Kilpatrick, C.W. Infraorder Hystricognathi. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; p. 1542. ISBN 978-0-8018-8221-0. [Google Scholar]
- O’Riain, M.J.; Faulkes, C.G. African mole rats: Eusociality, relatedness and ecological constraints. In Ecology of Social Evolution; Korb., J., Heinze., J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 207–223. [Google Scholar]
- Chung, D.; Dzal, Y.A.; Seow, A.; Milsom, W.K.; Pamenter, M.E. Naked mole rats exhibit metabolic but not ventilatory plasticity following chronic sustained hypoxia. Proc. Biol. Sci. 2016, 283, 20160216. [Google Scholar] [CrossRef]
- Pamenter, M.E.; Dzal, Y.A.; Milsom, W.K. Adenosine receptors mediate the hypoxic ventilatory response but not the hypoxic metabolic response in the naked mole rat during acute hypoxia. Proc. Biol. Sci. 2015, 282, 20141722. [Google Scholar] [CrossRef]
- Pamenter, M.E.; Lau, G.Y.; Richards, J.G.; Milsom, W.K. Naked mole rat brain mitochondria electron transport system flux and H+ leak are reduced during acute hypoxia. J. Exp. Biol. 2018, 221, jeb171397. [Google Scholar] [CrossRef]
- Park, T.J.; Reznick, J.; Peterson, B.L.; Blass, G.; Omerbasic, D.; Bennett, N.C.; Kuich, P.H.J.L.; Zasada, C.; Browe, B.M.; Hamann, W.; et al. Fructose-driven glycolysis supports anoxia resistance in the naked mole-rat. Science 2017, 356, 305–308. [Google Scholar] [CrossRef]
- Hochachka, P.W. Defense strategies against hypoxia and hypothermia. Science 1986, 231, 234–241. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Buck, L.T.; Doll, C.J.; Land, S.C. Unifying theory of hypoxia tolerance: Molecular/metabolic defense and rescue mechanisms for surviving oxygen lack. Proc. Natl. Acad. Sci. USA 1996, 93, 9493–9498. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.T.; Pamenter, M.E. Adaptive responses of vertebrate neurons to anoxia—Matching supply to demand. Respir. Physiol. Neurobiol. 2006, 154, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.T.; Pamenter, M.E. The hypoxia-tolerant vertebrate brain: Arresting synaptic activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2018, 224, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Ilacqua, A.N.; Kirby, A.M.; Pamenter, M.E. Behavioural responses of naked mole rats to acute hypoxia and anoxia. Biol. Lett. 2017, 13, 20170545. [Google Scholar] [CrossRef]
- Houlahan, C.R.; Kirby, A.M.; Dzal, Y.A.; Fairman, G.D.; Pamenter, M.E. Divergent behavioural responses to acute hypoxia between individuals and groups of naked mole rats. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2018, 224, 38–44. [Google Scholar] [CrossRef]
- Kirby, A.M.; Fairman, G.D.; Pamenter, M.E. Atypical behavioural, metabolic and thermoregulatory responses to hypoxia in the naked mole rat (Heterocephalus glaber). J. Zool. 2018, 305, 106–115. [Google Scholar] [CrossRef]
- Park, T.J.; Lu, Y.; Juttner, R.; Smith, E.S.; Hu, J.; Brand, A.; Wetzel, C.; Milenkovic, N.; Erdmann, B.; Heppenstall, P.A.; et al. Selective inflammatory pain insensitivity in the African naked mole-rat (Heterocephalus glaber). PLoS Biol. 2008, 6, e13. [Google Scholar] [CrossRef]
- Branigan, T.; Elkhalifa, S.; Pamenter, M.E. Behavioural responses to environmental hypercapnia in two eusocial species of African mole rats. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2018, 204, 811–819. [Google Scholar] [CrossRef]
- Seluanov, A.; Hine, C.; Azpurua, J.; Feigenson, M.; Bozzella, M.; Mao, Z.; Catania, K.C.; Gorbunova, V. Hypersensitivity to contact inhibition provides a clue to cancer resistance of naked mole-rat. Proc. Natl. Acad. Sci. USA 2009, 106, 19352–19357. [Google Scholar] [CrossRef]
- Tian, X.; Azpurua, J.; Hine, C.; Vaidya, A.; Myakishev-Rempel, M.; Ablaeva, J.; Mao, Z.; Nevo, E.; Gorbunova, V.; Seluanov, A. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature 2013, 499, 346–349. [Google Scholar] [CrossRef]
- Woodley, R.; Buffenstein, R. Thermogenic changes with chronic cold exposure in the naked mole-rat (Heterocephalus glaber). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 827–834. [Google Scholar] [CrossRef]
- Vandewint, A.L.; Zhu-Pawlowsky, A.J.; Kirby, A.; Tattersall, G.J.; Pamenter, M.E. Evaporative cooling and vasodilation mediate thermoregulation in naked mole-rats during normoxia but not hypoxia. J. Therm. Biol. 2019, 84, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A.; Chávez-Valencia, V. Learning of nature: The curious case of the naked mole rat. Mech. Ageing Dev. 2017, 164, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Munro, D.; Baldy, C.; Pamenter, M.E.; Treberg, J.R. The exceptional longevity of the naked mole-rat may be explained by mitochondrial antioxidant defenses. Aging Cell 2019, 18, e12916. [Google Scholar] [CrossRef]
- Munro, D.; Pamenter, M.E. Comparative studies of mitochondrial reactive oxygen species in animal longevity: Technical pitfalls and possibilities. Aging Cell 2019, 18, e13009. [Google Scholar] [CrossRef]
- Viltard, M.; Durand, S.; Pérez-Lanzón, M.; Aprahamian, F.; Lefevre, D.; Leroy, C.; Madeo, F.; Kroemer, G.; Friedlander, G. The metabolomic signature of extreme longevity: Naked mole rats versus mice. Aging 2019, 11, 4783–4800. [Google Scholar] [CrossRef]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of cancer resistance in long-lived mammals. Nat. Rev. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef]
- Ruby, J.G.; Smith, M.; Buffenstein, R. Naked Mole-Rat mortality rates defy gompertzian laws by not increasing with age. Elife 2018, 7, e31157. [Google Scholar] [CrossRef]
- Stenvinkel, P.; Shiels, P.G. Long-lived animals with negligible senescence: Clues for ageing research. Biochem. Soc. Trans. 2019, 47, 1157–1164. [Google Scholar] [CrossRef]
- Kim, E.B.; Fang, X.; Fushan, A.A.; Huang, Z.; Lobanov, A.V.; Han, L.; Marino, S.M.; Sun, X.; Turanov, A.A.; Yang, P.; et al. Genome sequencing reveals insights into physiology and longevity of the naked mole rat. Nature 2011, 479, 223–227. [Google Scholar] [CrossRef]
- Tan, L.; Ke, Z.; Tombline, G.; Macoretta, N.; Hayes, K.; Tian, X.; Lv, R.; Ablaeva, J.; Gilbert, M.; Bhanu, N.V.; et al. Naked Mole Rat Cells Have a Stable Epigenome that Resists iPSC Reprogramming. Stem Cell Rep. 2017, 9, 1721–1734. [Google Scholar] [CrossRef] [PubMed]
- Hilton, H.G.; Rubinstein, H.G.; Janki, N.D.; Ireland, P.; Bernstein, A.T.; Wright, N.; Finkle, K.M.; Martin-McNulty, D.; Roy, M.; et al. Single-cell transcriptomics of the naked mole-rat reveals unexpected features of mammalian immunity. bioRxiv 2019, 597195. [Google Scholar] [CrossRef]
- Frankenberg, A.D.V.; Reis, A.F.; Gerchman, F. Relationships between adiponectin levels, the metabolic syndrome, and type 2 diabetes: A literature review. Arch. Endocrinol. Metab. 2017, 61, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Spracklen, C.N.; Karaderi, T.; Yaghootkar, H.; Schurmann, C.; Fine, R.S.; Kutalik, Z.; Preuss, M.H.; Lu, Y.; Wittemans, L.B.L.; Adair, L.S.; et al. Exome-Derived Adiponectin-Associated Variants Implicate Obesity and Lipid Biology. Am. J. Hum. Genet. 2019, 105, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tao, J.; Zhao, P.J.; Tang, W.; Xu, J.P.; Zhang, K.Q.; Zou, C.G. Adiponectin receptor PAQR-2 signaling senses low temperature to promote C. elegans longevity by regulating autophagy. Nat. Commun. 2019, 10, 2602. [Google Scholar] [CrossRef]
- Fiaschi, T. Mechanisms of Adiponectin Action. Int. J. Mol. Sci. 2019, 20, 2894. [Google Scholar] [CrossRef]
- Fiaschi, T.; Magherini, F.; Gamberi, T.; Modesti, P.A.; Modesti, A. Adiponectin as a tissue regenerating hormone: More than a metabolic function. Cell. Mol. Life Sci. 2014, 71, 1917–1925. [Google Scholar] [CrossRef]
- Parida, S.; Siddharth, S.; Sharma, D. Adiponectin, obesity, and cancer: Clash of the bigwigs in health and disease. Int. J. Mol. Sci. 2019, 20, 2519. [Google Scholar] [CrossRef] [PubMed]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. MiRNA-210: A Current Overview. Anticancer Res. 2017, 37, 6511–6521. [Google Scholar] [PubMed]
- Tang, J.; Ning, R.; Zeng, B.; Li, Y. Molecular Evolution of PTEN Pseudogenes in Mammals. PLoS ONE 2016, 11, e0167851. [Google Scholar] [CrossRef] [PubMed]
- Altwasser, R.; Paz, A.; Korol, A.; Manov, I.; Avivi, A.; Shams, I. The transcriptome landscape of the carcinogenic treatment response in the blind mole rat: Insights into cancer resistance mechanisms. BMC Genom. 2019, 20, 17. [Google Scholar] [CrossRef]
- Musso, G.; Cassader, M.; Cohney, S.; De Michieli, F.; Pinach, S.; Saba, F.; Gambino, R. Fatty Liver and Chronic Kidney Disease: Novel Mechanistic Insights and Therapeutic Opportunities. Diabetes Care 2016, 39, 1830–1845. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A. microRNA dysregulation in neurodegenerative diseases: A systematic review. Prog. Neurobiol. 2019, 26, 101664. [Google Scholar] [CrossRef]
- Li, J.; Li, K.; Chen, X. Inflammation-regulatory microRNAs: Valuable targets for intracranial atherosclerosis. J. Neurosci. Res. 2019, 97, 1242–1252. [Google Scholar] [CrossRef]
- Xu, H.X.; Pan, W.; Qian, J.F.; Liu, F.; Dong, H.Q.; Liu, Q.J. MicroRNA-21 contributes to the puerarin-induced cardioprotection via suppression of apoptosis and oxidative stress in a cell model of ischemia/reperfusion injury. Mol. Med. Rep. 2019, 20, 719–727. [Google Scholar] [CrossRef]
- Xiaoyan, W.; Pais, E.M.; Lan, L.; Jingrui, C.; Lin, M.; Fordjour, P.A.; Guanwei, F. MicroRNA-155: A Novel Armamentarium Against Inflammatory Diseases. Inflammation 2017, 40, 708–716. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Zhang, H.; Huang, P.; Luthra, R. Hypoxia-regulated microRNA-210 modulates mitochondrial function and decreases ISCU and COX10 expression. Oncogene 2010, 29, 4362–4368. [Google Scholar] [CrossRef]
- Voloboueva, L.A.; Sun, X.; Xu, L.; Ouyang, Y.-B.; Giffard, R.G. Distinct effects of miR-210 reduction on neurogenesis: Increased neuronal survival of inflammation but reduced proliferation associated with mitochondrial enhancement. J. Neurosci. 2017, 37, 3072–3084. [Google Scholar] [CrossRef] [PubMed]
- Favaro, E.; Ramachandran, A.; McCormick, R.; Gee, H.; Blancher, C.; Crosby, M.; Devlin, C.; Blick, C.; Buffa, F.; Li, J.-L.; et al. MicroRNA-210 regulates mitochondrial free radical response to hypoxia and krebs cycle in cancer cells by targeting iron sulfur cluster protein ISCU. PLoS ONE 2010, 5, e10345. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Le, Q.-T.; Giaccia, A.J. MiR-210—Micromanager of the hypoxia pathway. Trends Mol. Med. 2010, 16, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Pamenter, M.E.; Dzal, Y.A.; Thompson, W.A.; Milsom, W.K. Do naked mole rats accumulate a metabolic acidosis or an oxygen debt in severe hypoxia? J. Exp. Biol. 2019, 222, jeb191197. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.M.; Goldman, B.D.; Goldman, S.L.; Seney, M.L.; Forger, N.G. Neuroendocrinology and sexual differentiation in eusocial mammals. Front. Neuroendocrinol. 2009, 30, 519–533. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Nicholas, A.P.; Whitaker, J.N. Preparation of a monoclonal antibody to citrullinated epitopes: Its characterization and some applications to immunohistochemistry in human brain. Glia 2002, 37, 328–336. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 −ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Symbol | Total Score (p < 0.05) # |
|---|---|---|
| Apolipoprotein B-100 | G5AZB7_HETGA | 6077 |
| Alpha-2-macroglobulin | G5BPM1_HETGA | 4974 |
| Complement C3 | G5C0N5_HETGA | 4300 |
| Serotransferrin | G5BQA9_HETGA | 4058 |
| Serum albumin | G5B5P2_HETGA | 3394 |
| Fibronectin | G5BHR4_HETGA | 3333 |
| Fibrinogen beta chain | G5BML3_HETGA | 2638 |
| Kininogen-1 | G5BT86_HETGA | 2276 |
| Fibrinogen alpha chain | G5BML2_HETGA | 2094 |
| Ceruloplasmin | G5BSL1_HETGA | 2022 |
| Histidine-rich glycoprotein | G5BT87_HETGA | 1851 |
| Complement C4-A | G5C3H6_HETGA | 1759 |
| Complement factor H | G5BM71_HETGA | 1646 |
| Plasminogen | G5BC53_HETGA | 1548 |
| Inter-alpha-trypsin inhibitor heavy chain H4 | G5BUN4_HETGA | 1455 |
| Fibrinogen gamma chain | G5BML1_HETGA | 1338 |
| Complement factor B | G5C3H1_HETGA | 1313 |
| Plasma kallikrein | G5BNV2_HETGA | 1167 |
| Antithrombin-III | G5ARS6_HETGA | 1093 |
| Coagulation factor V | G5CB46_HETGA | 1092 |
| Complement C5 | G5AXS5_HETGA | 1002 |
| Hemoglobin subunit beta | G5BS33_HETGA | 941 |
| Transcobalamin-2 isoform 1 | G5AVP0_HETGA | 913 |
| Hemopexin | G5BBR0_HETGA | 899 |
| Coagulation factor XIII B chain | G5BM72_HETGA | 821 |
| Protein AMBP | G5B1Y4_HETGA | 817 |
| N-acetylmuramoyl-L-alanine amidase | G5BYP3_HETGA | 812 |
| Coagulation factor XII | G5BQ09_HETGA | 802 |
| Apolipoprotein A-I | APOA1_HETGA | 798 |
| Hemoglobin subunit alpha | G5BXY1_HETGA | 784 |
| Coagulation factor XIII A chain | G5BAS8_HETGA | 779 |
| Hemoglobin subunit beta | G5BYJ8_HETGA | 759 |
| Alpha-1-antiproteinase S | G5B496_HETGA | 724 |
| Inter-alpha-trypsin inhibitor heavy chain H3 | G5BUN3_HETGA | 682 |
| Vitronectin | G5BVN8_HETGA | 652 |
| Complement factor I | G5AQM1_HETGA | 647 |
| Hemoglobin subunit epsilon-1 | G5BS35_HETGA | 642 |
| Inter-alpha-trypsin inhibitor heavy chain H2 | G5AXV8_HETGA | 584 |
| Apolipoprotein E | G5CBM7_HETGA | 581 |
| Inhibitor of carbonic anhydrase | G5BQB0_HETGA | 537 |
| Inter-alpha-trypsin inhibitor heavy chain H1 | G5BUN2_HETGA | 482 |
| Four and a half LIM domains protein 1 | G5CA61_HETGA | 469 |
| Haptoglobin | G5B5U6_HETGA | 464 |
| C4b-binding protein | G5BP10_HETGA | 438 |
| L-lactate dehydrogenase | G5AKA3_HETGA | 437 |
| Insulin-like growth factor-binding protein complex acid labile chain | G5BY64_HETGA | 403 |
| Catalase | G5AXV0_HETGA | 382 |
| Fetuin-B | G5BT88_HETGA | 347 |
| Alpha-2-HS-glycoprotein | G5BT89_HETGA | 337 |
| Selenoprotein P | G5APA7_HETGA | 328 |
| Vitamin D-binding protein | G5BE53_HETGA | 317 |
| Adiponectin | G5BT83_HETGA | 283 |
| von Willebrand factor | G5CAN6_HETGA | 274 |
| Beta-2-glycoprotein 1 | G5BGY7_HETGA | 268 |
| Basement membrane-specific heparan sulfate proteoglycan core protein | G5BI06_HETGA | 267 |
| Gelsolin | G5AXS0_HETGA | 259 |
| Aspartyl aminopeptidase | G5AKJ4_HETGA | 256 |
| Complement C1q subcomponent subunit A | G5BHZ8_HETGA | 246 |
| Ficolin-3 | G5AUT5_HETGA | 241 |
| Mannan-binding lectin serine protease 1 | G5BTD5_HETGA | 228 |
| Alpha-1-antichymotrypsin | G5B491_HETGA | 228 |
| Carboxypeptidase N catalytic chain | G5AYP3_HETGA | 210 |
| Complement C1q subcomponent subunit C | G5BHZ7_HETGA | 191 |
| Mannan-binding lectin serine protease 2 | G5C521_HETGA | 188 |
| Prothrombin | G5ATC4_HETGA | 177 |
| Complement C1q tumour necrosis factor-related protein 3 | G5BQ97_HETGA0 | 177 |
| Nucleoside diphosphate kinase | G5C4L3_HETGA | 168 |
| L-lactate dehydrogenase | G5BEG2_HETGA | 167 |
| Alpha-2-antiplasmin | G5BCV1_HETGA | 164 |
| Keratin, type II cytoskeletal 6B | G5ALS1_HETGA | 152 |
| Carbonic anhydrase 2 | G5ATW7_HETGA | 143 |
| Proprotein convertase subtilisin/kexin type 9 | G5APQ4_HETGA | 137 |
| Angiotensinogen | G5BQI5_HETGA | 121 |
| 6-phosphogluconate dehydrogenase, decarboxylating | G5C530_HETGA | 91 |
| Nesprin-1 | G5C0E1_HETGA | 87 |
| Complement C1q subcomponent subunit B | G5BHZ6_HETGA | 87 |
| Angiopoietin-related protein 6 | G5B341_HETGA | 85 |
| Hepatocyte growth factor activator | G5BZF5_HETGA | 84 |
| Calpain-1 catalytic subunit | G5B6L3_HETGA | 84 |
| Sulfhydryl oxidase | G5AYL7_HETGA | 80 |
| Keratin, type II cytoskeletal 5 | G5ALS3_HETGA | 76 |
| Profilin | G5BH50_HETGA | 75 |
| 40S ribosomal protein S27a | G5B8W4_HETGA | 68 |
| Xanthine dehydrogenase/oxidase | G5B3Z0_HETGA | 68 |
| Plasma protease C1 inhibitor | G5BLJ5_HETGA | 67 |
| Collectin-11 | G5C7L1_HETGA | 66 |
| Extracellular matrix protein 1 | G5BH40_HETGA | 64 |
| Actin, gamma-enteric smooth muscle | G5AXH0_HETGA | 61 |
| Collectin-10 | G5C9R8_HETGA | 61 |
| Keratin, type I cytoskeletal 17 | G5B0M4_HETGA | 58 |
| Contactin-1 | G5BGJ6_HETGA | 55 |
| Dynein heavy chain 1, axonemal | G5BUL8_HETGA | 50 |
| Leucine-rich repeat-containing protein 15 | G5ALE6_HETGA | 43 |
| Transcriptional regulator ATRX | G5C0I5_HETGA | 42 |
| Transmembrane protein 127 | G5BMW1_HETGA | 41 |
| TRAF3-interacting protein 1 | G5BHH8_HETGA | 41 |
| Mannose-binding protein A | G5C4H7_HETGA | 40 |
| Hyaluronan-binding protein 2 | G5BKD5_HETGA | 39 |
| Fer3-like protein | G5BZJ1_HETGA | 38 |
| Complement component C7 | G5C4R4_HETGA | 38 |
| PITH domain-containing protein | G5BHY3_HETGA | 38 |
| 1,4-alpha-glucan-branching enzyme | G5BA56_HETGA | 37 |
| tRNA pseudouridine synthase | G5BB54_HETGA | 37 |
| SRRM2-like protein | G5BG61_HETGA | 34 |
| Putative carboxypeptidase PM20D1 | G5BX50_HETGA | 34 |
| Olfactory receptor | G5C741_HETGA | 33 |
| Protein CASC5 | G5BLZ4_HETGA | 33 |
| Conserved oligomeric Golgi complex subunit 1 | G5B5M3_HETGA | 32 |
| Small G protein signalling modulator 1 | G5BGN5_HETGA | 32 |
| Eukaryotic translation initiation factor 3 subunit E | G5BD61_HETGA | 31 |
| HEAT repeat-containing protein 2 | G5AZ15_HETGA | 31 |
| Proline-rich protein 11 | G5AXY5_HETGA | 31 |
| Peroxiredoxin-1 | G5ARW1_HETGA | 30 |
| Pre-mRNA-processing factor 17 | G5B0Y2_HETGA | 30 |
| Ribonucleoside-diphosphate reductase | G5B9C2_HETGA | 30 |
| Protein Name | Symbol | Total Score (p < 0.05) # |
|---|---|---|
| Alpha-2-macroglobulin | G5BPM1_HETGA | 2978 |
| Serum albumin | G5B5P2_HETGA | 2570 |
| Serotransferrin | G5BQA9_HETGA | 1976 |
| Keratin, type II cytoskeletal 6B | G5ALS1_HETGA | 1459 |
| Complement C3 | G5C0N5_HETGA | 1440 |
| Keratin, type II cytoskeletal 5 | G5ALS3_HETGA | 1211 |
| Kininogen-1 | G5BT86_HETGA | 1196 |
| Histidine-rich glycoprotein | G5BT87_HETGA | 1186 |
| Keratin, type I cytoskeletal 10 | G5AX66_HETGA | 1130 |
| Keratin, type I cytoskeletal 14 | G5B0M6_HETGA | 1038 |
| Keratin, type II cytoskeletal 1 | G5ALS8_HETGA | 1011 |
| Fibrinogen beta chain | G5BML3_HETGA | 974 |
| Fibrinogen alpha chain | G5BML2_HETGA | 956 |
| Apolipoprotein B-100 | G5AZB7_HETGA | 834 |
| Keratin, type I cytoskeletal 13 | G5B0N0_HETGA | 701 |
| Keratin, type I cytoskeletal 17 | G5B0M4_HETGA | 673 |
| Desmoplakin | G5BAT4_HETGA | 670 |
| Hemoglobin subunit alpha | G5BXY1_HETGA | 591 |
| Hemoglobin subunit beta | G5BS33_HETGA | 581 |
| Complement C4-A | G5C3H6_HETGA | 571 |
| Hemoglobin subunit beta | G5BYJ8_HETGA | 568 |
| Apolipoprotein A-I | APOA1_HETGA | 499 |
| Keratin, type II cytoskeletal 79 | G5BJ37_HETGA | 468 |
| Actin, cytoplasmic 1 | G5BI78_HETGA | 444 |
| Keratin, type II cytoskeletal 4 | G5BJ36_HETGA | 443 |
| Fibrinogen gamma chain | G5BML1_HETGA | 382 |
| Ceruloplasmin | G5BSL1_HETGA | 350 |
| Apolipoprotein E | G5CBM7_HETGA | 338 |
| Inter-alpha-trypsin inhibitor heavy chain H4 | G5BUN4_HETGA | 332 |
| Protein AMBP | G5B1Y4_HETGA | 280 |
| Inhibitor of carbonic anhydrase | G5BQB0_HETGA | 244 |
| Hemopexin | G5BBR0_HETGA | 239 |
| Histone H4 | G5BKL3_HETGA | 238 |
| Plasminogen | G5BC53_HETGA | 232 |
| Alpha-1-antiproteinase S | G5B496_HETGA | 217 |
| Hemoglobin subunit epsilon-1 | G5BS35_HETGA | 213 |
| Junction plakoglobin | G5B0M0_HETGA | 202 |
| N-acetylmuramoyl-L-alanine amidase | G5BYP3_HETGA | 190 |
| Inter-alpha-trypsin inhibitor heavy chain H3 | G5BUN3_HETGA | 190 |
| Keratin, type II cytoskeletal 7 | G5BL96_HETGA | 182 |
| Fibronectin | G5BHR4_HETGA | 164 |
| Glyceraldehyde-3-phosphate dehydrogenase | G5CAP7_HETGA | 154 |
| Coagulation factor XII | G5BQ09_HETGA | 152 |
| Antithrombin-III | G5ARS6_HETGA | 150 |
| Catenin beta-1 | G5ALX2_HETGA | 126 |
| Plakophilin-1 | G5B3A4_HETGA | 120 |
| Annexin | G5AWC0_HETGA | 119 |
| Keratin, type II cytoskeletal 3 | G5ALT2_HETGA | 118 |
| 40S ribosomal protein S27a | G5B8W4_HETGA | 116 |
| Inter-alpha-trypsin inhibitor heavy chain H2 | G5AXV8_HETGA | 106 |
| Alpha-2-antiplasmin | G5BCV1_HETGA | 86 |
| Complement factor B | G5C3H1_HETGA | 81 |
| Elongation factor 1-alpha | G5ALK7_HETGA | 78 |
| Angiotensinogen | G5BQI5_HETGA | 77 |
| Histone H2B | G5BH20_HETGA | 77 |
| Plasma kallikrein | G5BNV2_HETGA | 72 |
| Beta-enolase | G5BW96_HETGA | 70 |
| Vitamin D-binding protein | G5BE53_HETGA | 57 |
| Alpha-1-antichymotrypsin | G5B491_HETGA | 55 |
| Keratin, type II cytoskeletal 80 | G5BL95_HETGA | 53 |
| Heat shock cognate 71 kDa protein | G5B170_HETGA | 51 |
| Leucine-rich repeat-containing protein KIAA1731 | G5C3Y1_HETGA | 50 |
| Cathepsin D | G5C2G1_HETGA | 49 |
| Basement membrane-specific heparan sulfate proteoglycan core protein | G5BI06_HETGA | 47 |
| Skin-specific protein 32 | G5BUY9_HETGA | 46 |
| Desmocollin-1 | G5C312_HETGA | 46 |
| Protein-arginine deiminase type-6 | G5BZN1_HETGA | 46 |
| Cryptochrome-1 | G5C454_HETGA | 45 |
| Complement C1q subcomponent subunit C | G5BHZ7_HETGA | 45 |
| p2Y purinoceptor 12 | G5C102_HETGA | 44 |
| Alpha-1B-glycoprotein | G5B7K8_HETGA | 44 |
| Exportin-1 | G5C1Y9_HETGA | 44 |
| Putative hexokinase HKDC1 | G5B183_HETGA | 43 |
| Transcriptional regulator ATRX | G5C0I5_HETGA | 42 |
| Coiled-coil domain-containing protein 57 | G5BJ98_HETGA | 42 |
| Microtubule plus-end tracking protein TIP150 | G5ATU1_HETGA | 40 |
| Tubulin alpha-1C chain | G5AQ00_HETGA | 38 |
| Fer3-like protein | G5BZJ1_HETGA | 38 |
| Amino acid transporter | G5BYQ9_HETGA | 36 |
| Ventricular zone-expressed PH domain-containing protein-like protein 1 | G5B253_HETGA | 35 |
| SRRM2-like protein | G5BG61_HETGA | 35 |
| N6-adenosine-methyltransferase 70 kDa subunit | G5BFU9_HETGA | 33 |
| Proline-rich protein 11 | G5AXY5_HETGA | 33 |
| Carbonic anhydrase 2 | G5ATW7_HETGA | 32 |
| 26S proteasome non-ATPase regulatory subunit 3 | G5BRY1_HETGA | 31 |
| Serine/threonine-protein kinase LATS1 | G5BMD1_HETGA | 31 |
| Transmembrane gamma-carboxyglutamic acid protein 1 | G5C418_HETGA | 31 |
| Collagen alpha-2(I) chain | G5ANK8_HETGA | 31 |
| Bcl-2-associated transcription factor 1 | G5C1A7_HETGA | 30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pamenter, M.E.; Uysal-Onganer, P.; Huynh, K.W.; Kraev, I.; Lange, S. Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber). Int. J. Mol. Sci. 2019, 20, 5378. https://doi.org/10.3390/ijms20215378
Pamenter ME, Uysal-Onganer P, Huynh KW, Kraev I, Lange S. Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber). International Journal of Molecular Sciences. 2019; 20(21):5378. https://doi.org/10.3390/ijms20215378
Chicago/Turabian StylePamenter, Matthew E., Pinar Uysal-Onganer, Kenny W. Huynh, Igor Kraev, and Sigrun Lange. 2019. "Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber)" International Journal of Molecular Sciences 20, no. 21: 5378. https://doi.org/10.3390/ijms20215378
APA StylePamenter, M. E., Uysal-Onganer, P., Huynh, K. W., Kraev, I., & Lange, S. (2019). Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber). International Journal of Molecular Sciences, 20(21), 5378. https://doi.org/10.3390/ijms20215378