Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Patients
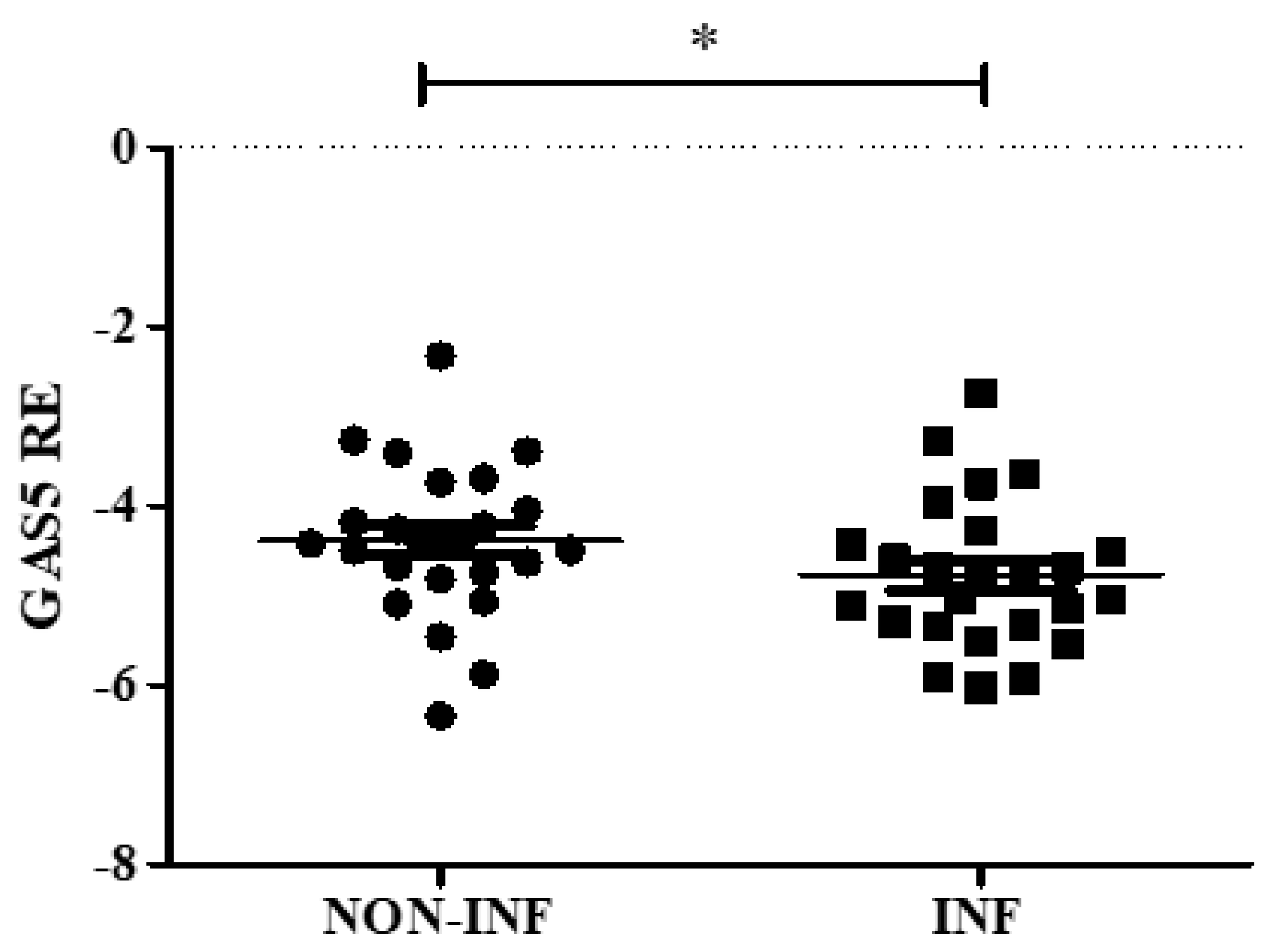
2.2. GAS5 Expression in Mucosal Biopsies of Pediatric IBD Patients
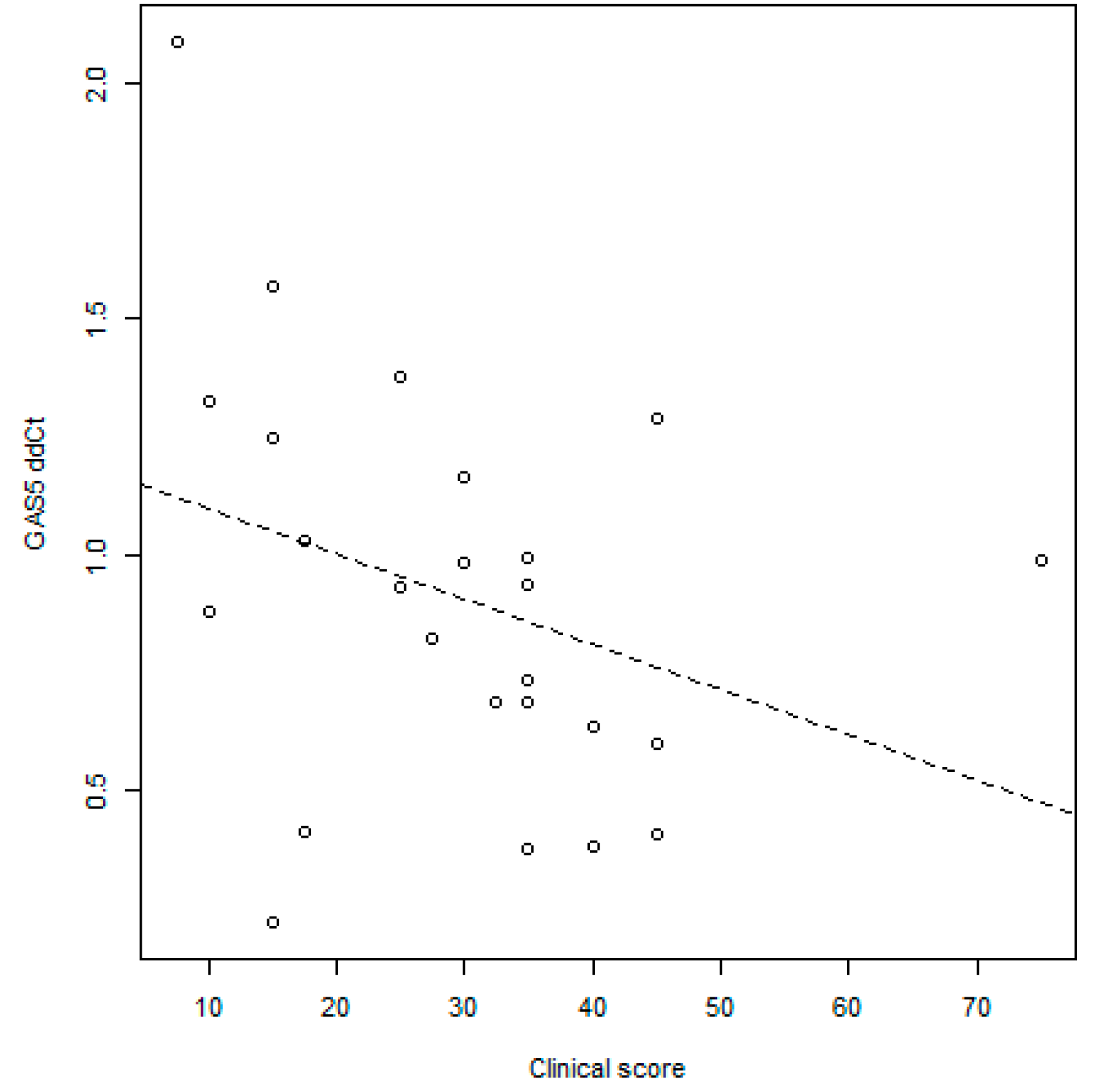
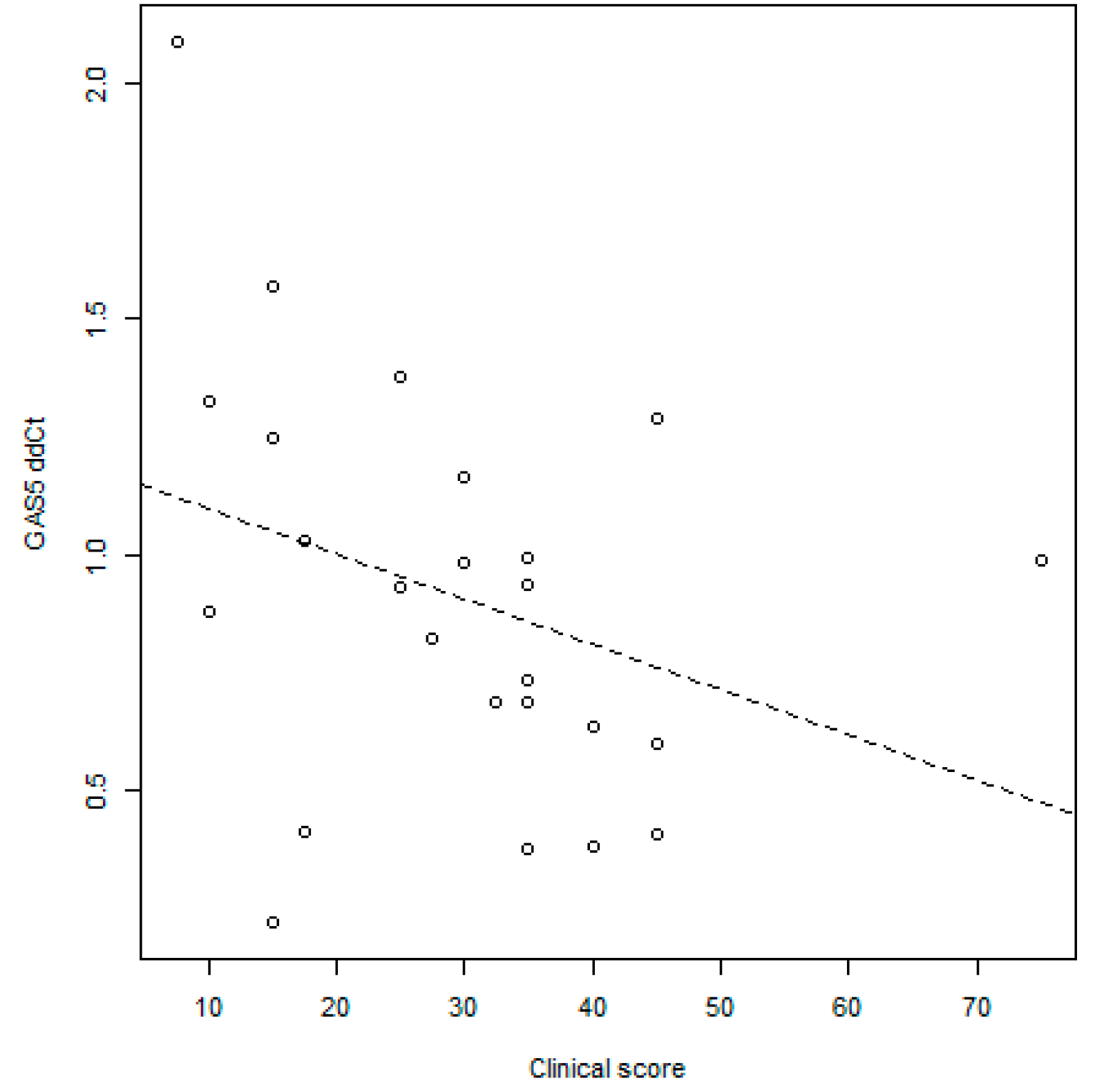
GAS5 Correlation with Disease Activity Scores
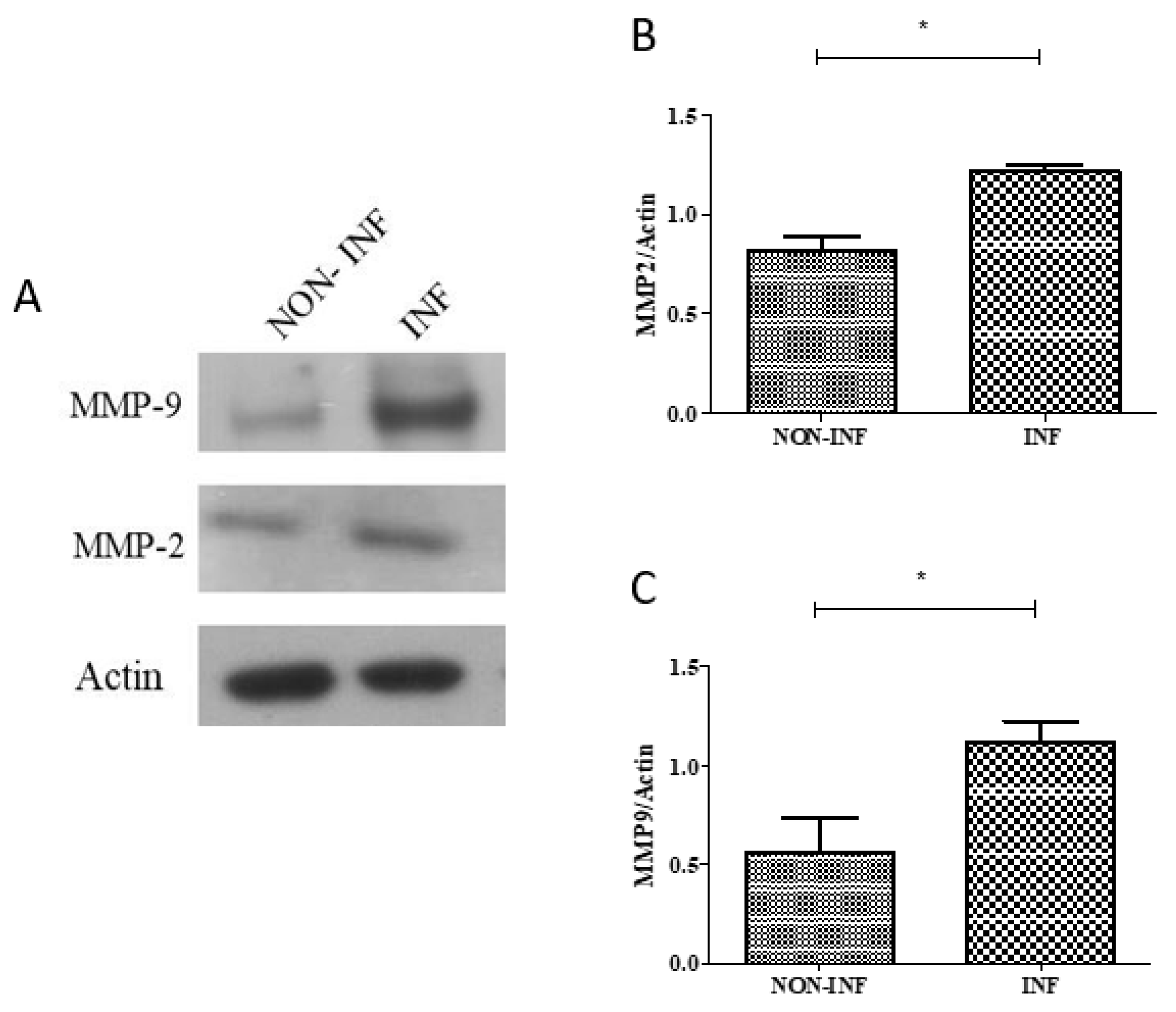
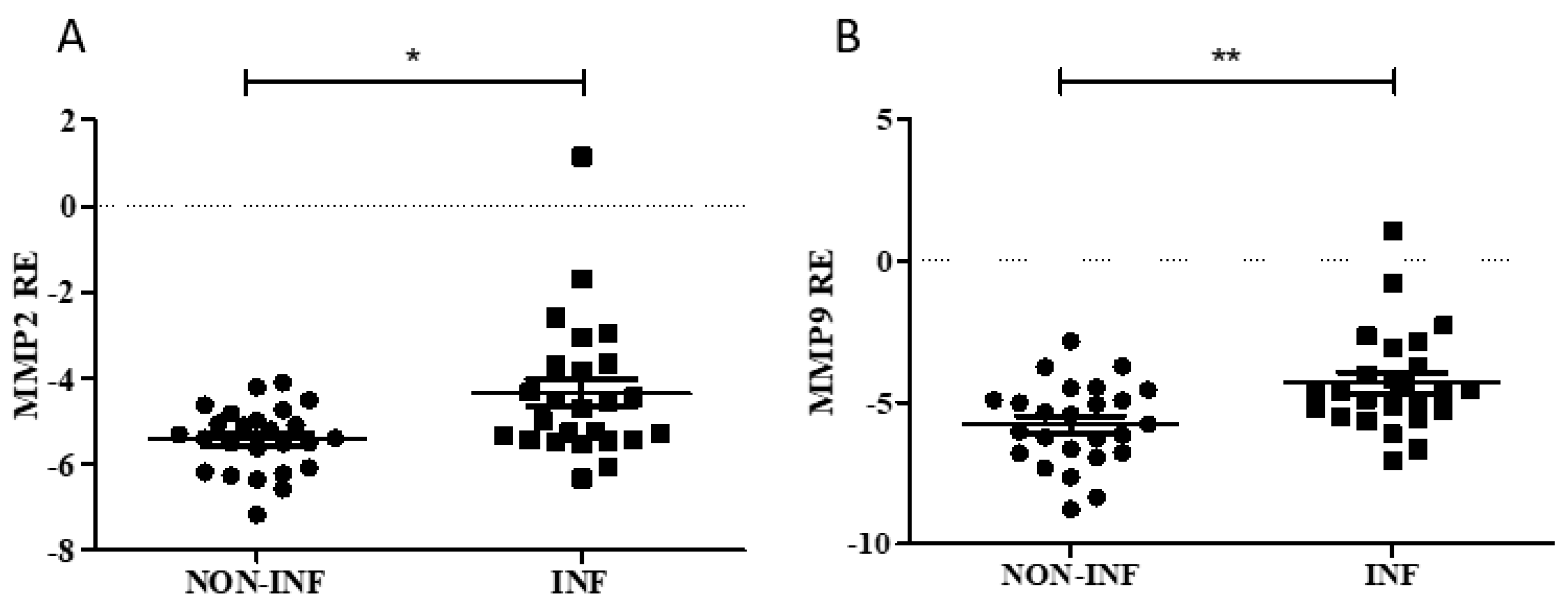
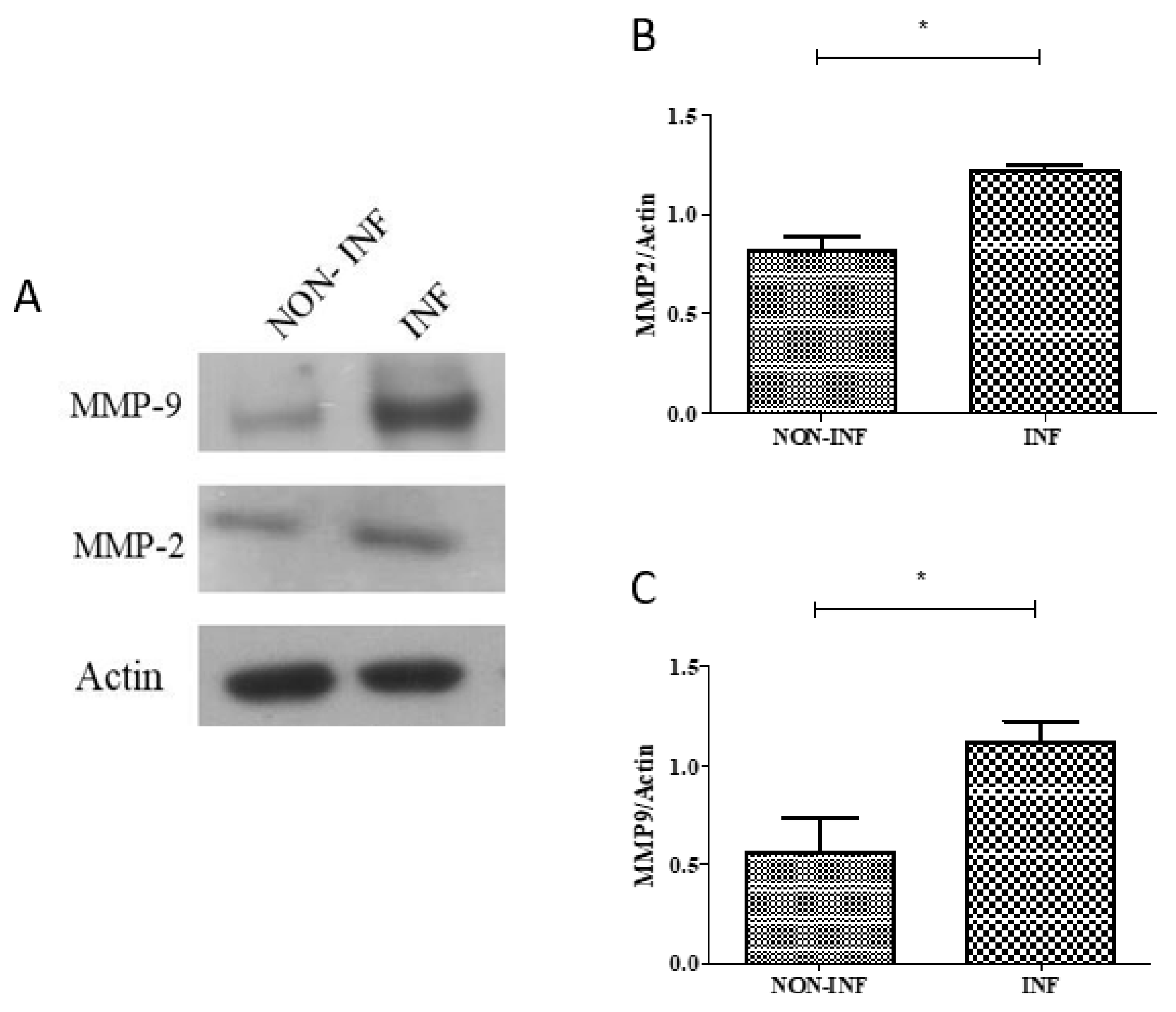
2.3. MMP2 and MMP9 Gene and Protein Expression in Colon Biopsies of Pediatric IBD Patients
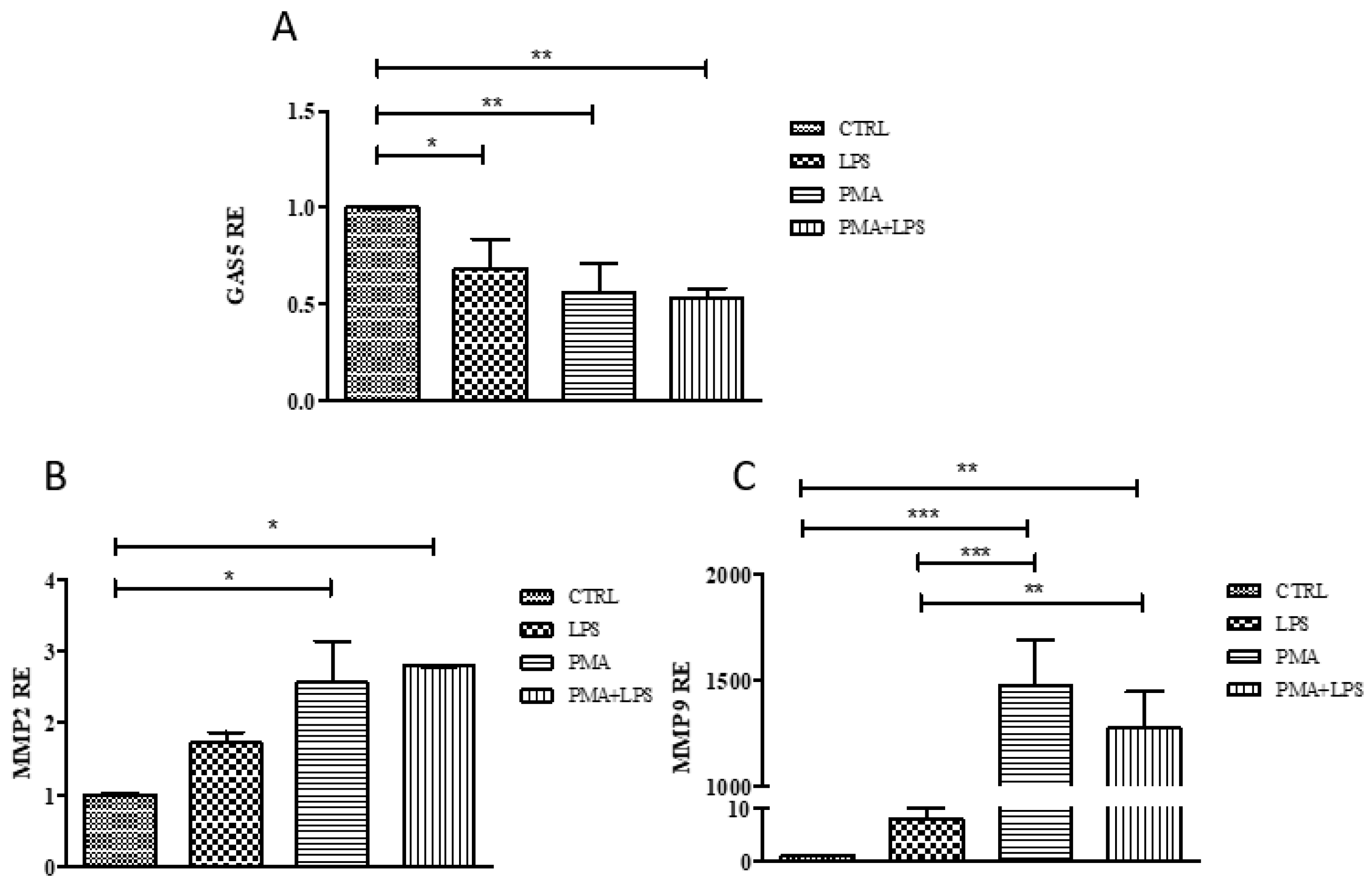
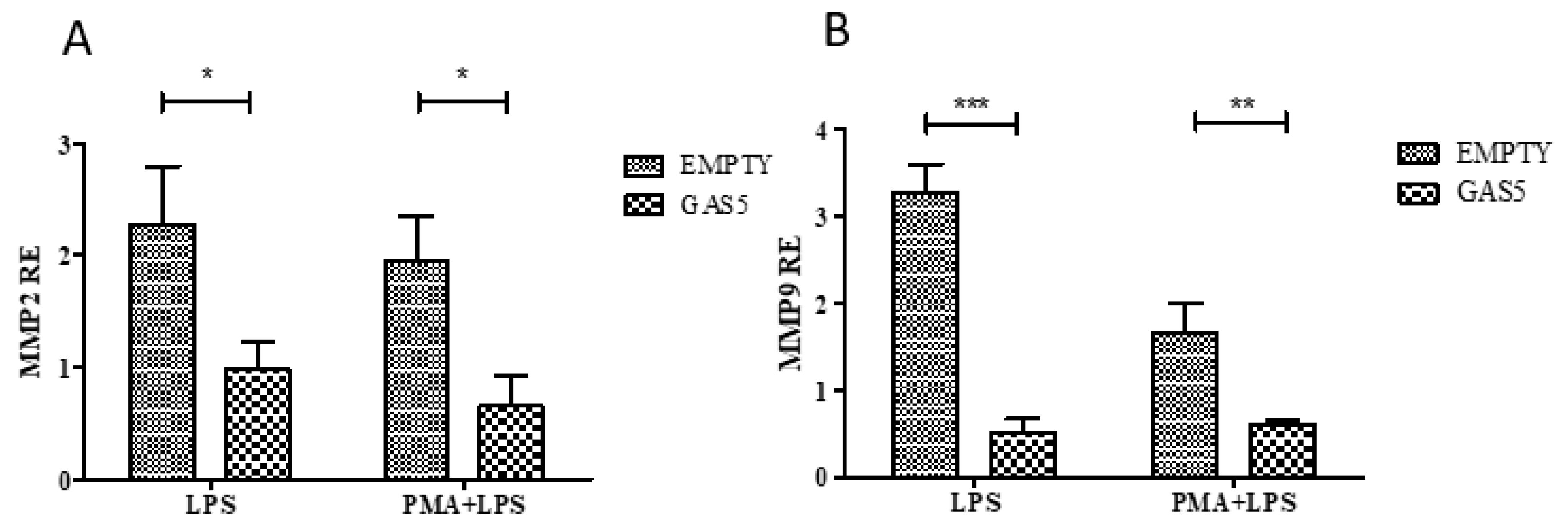
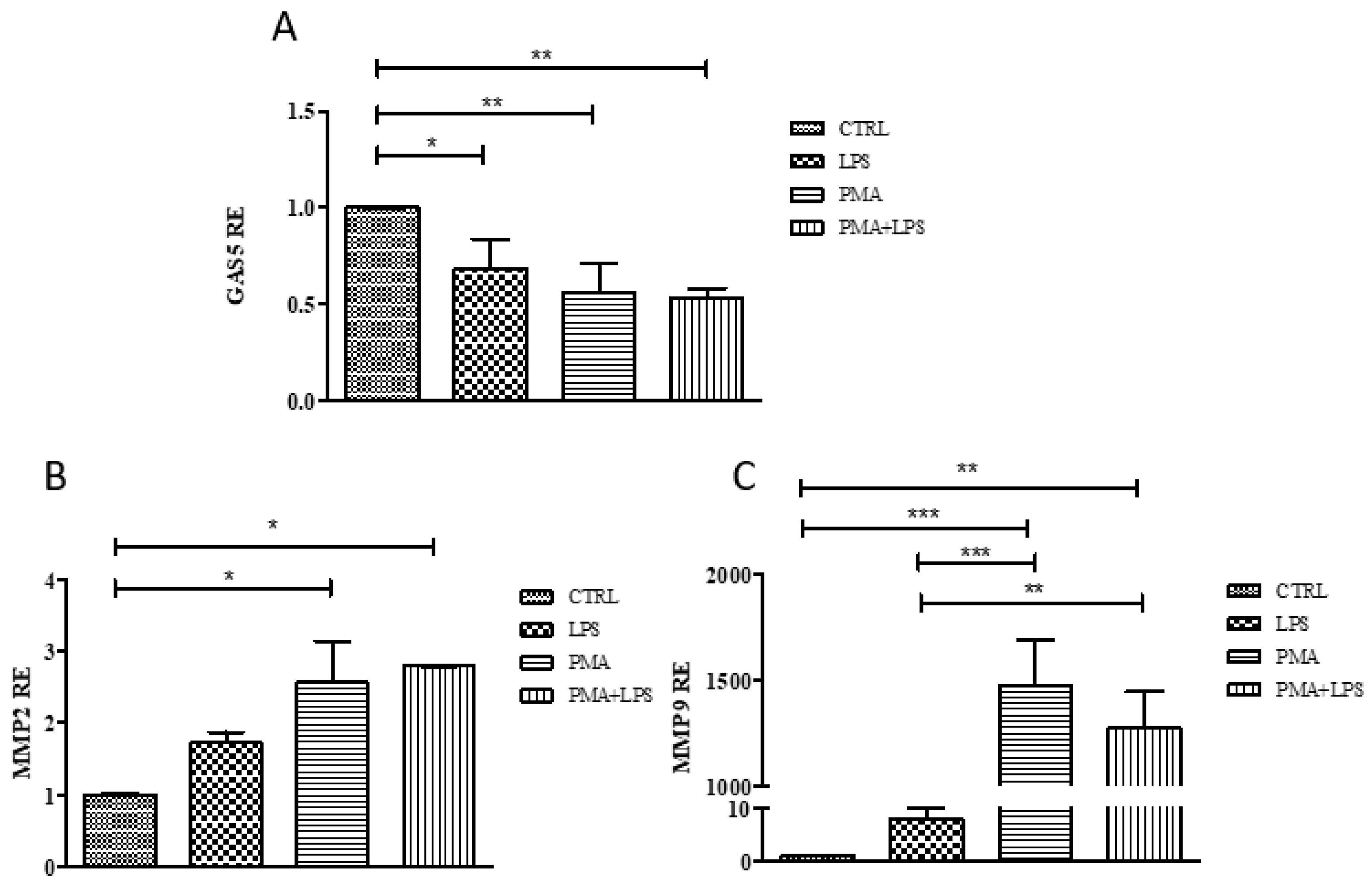
2.4. GAS5, MMP2, and MMP9 Gene Expression in the THP1 Cell Line
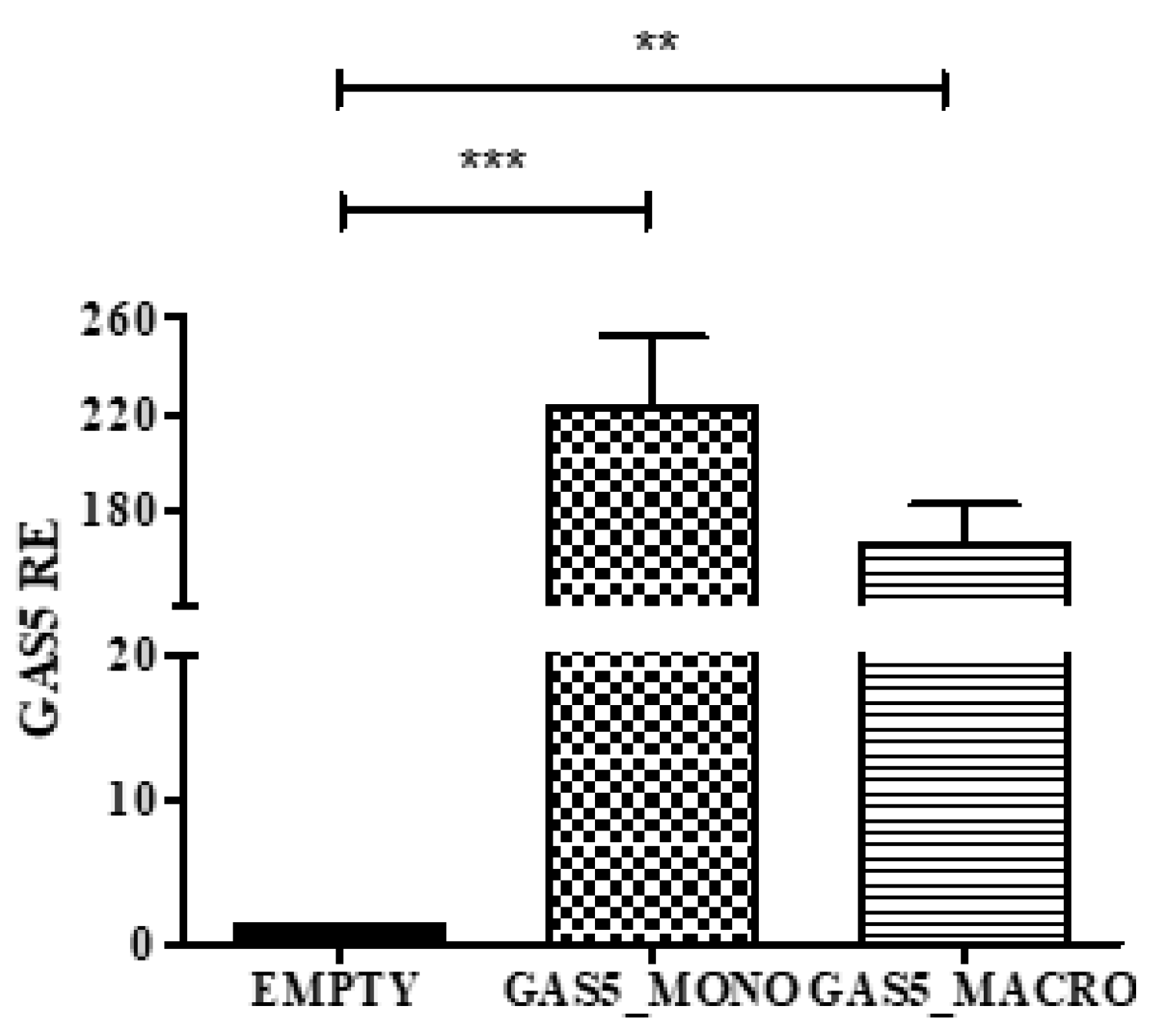
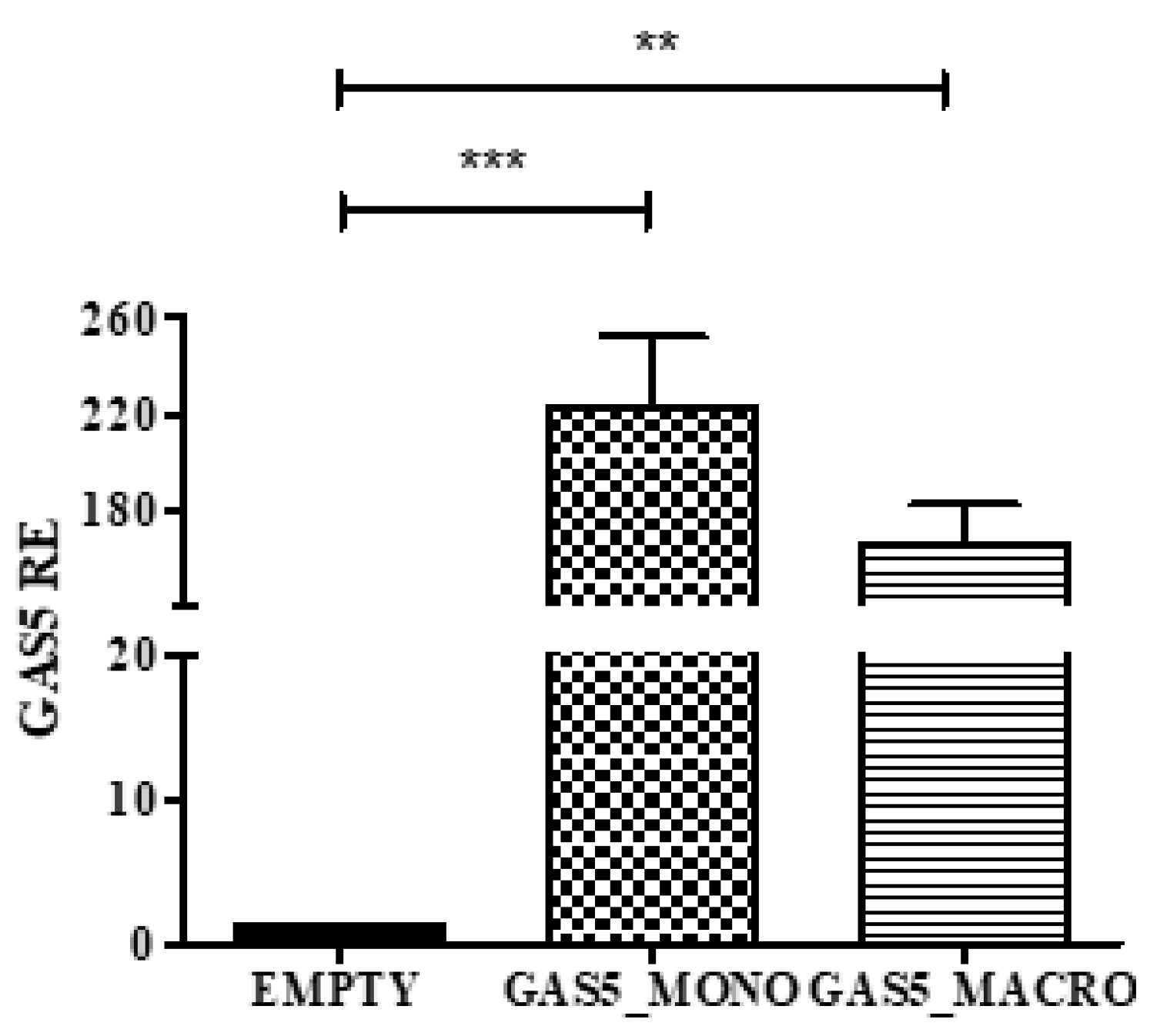
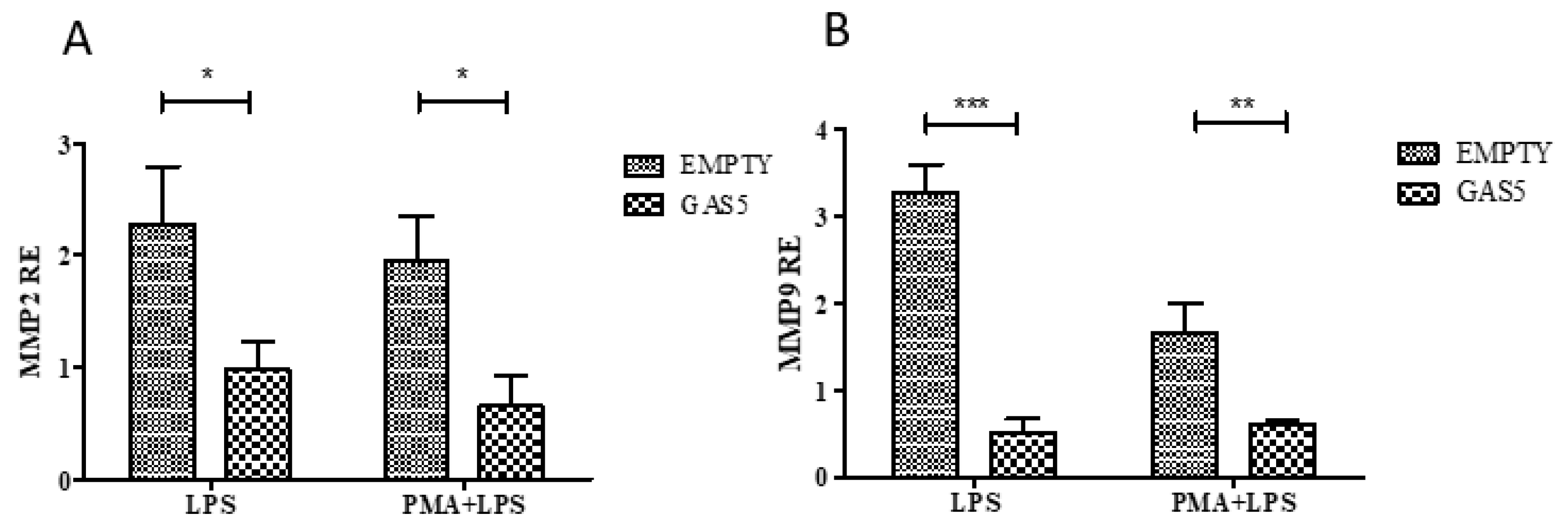
2.5. MMP2 and MMP9 Gene Expression in Cells Transfected with the pcDNA3.1-GAS5 Plasmid
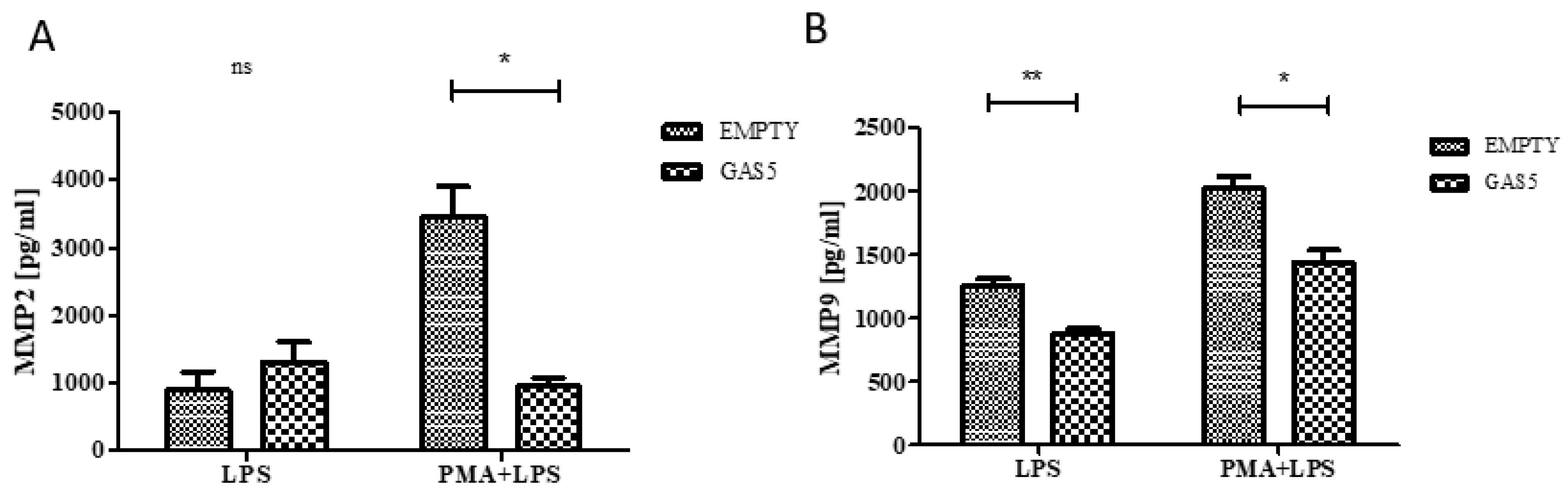
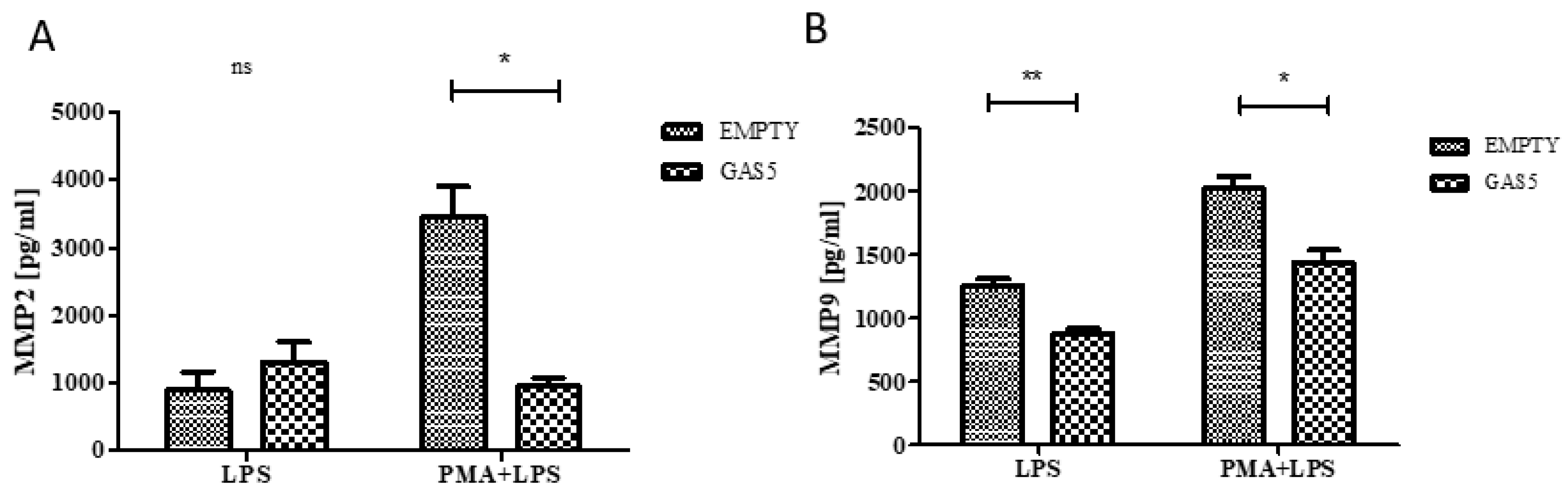
2.6. MMP2 and MMP9 Proteins Released by THP1 Cells Overexpressing GAS5
3. Discussion
4. Materials and Methods
4.1. Clinical Samples and Histologic Evaluation
- active inflammation (neutrophils aggressive toward the glandular structures in the lamina propria) (range from 0 to 3);
- crypt abscesses (range from 0 to 1);
- erosions and ulcerations (i.e., presence of granulation tissue) (range from 0 to 1).
- glandular crypts alteration (range from 0 to 3);
- basal plasmacytosis (range from 0 to 1);
- epithelioid granulomas (range from 0 to 1).
4.2. Ethical Considerations
4.3. Total RNA Isolation
4.4. Quantitative Real-Time PCR
4.5. Immunoblotting
4.6. Cell Line and Stimulation
4.7. Transient Transfection
4.8. MMP2 and MMP9 Quantification
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| lncRNA | Long non-coding RNA |
| GAS5 | Growth arrest–specific transcript 5 |
| MMPs | Matrix metalloproteinases |
| IBD | Inflammatory bowel disease |
| CD | Crohn’s disease |
| UC | Ulcerative colitis |
| LPS | Lipopolysaccharide |
| PMA | Phorbol-12-myristate 13-acetate |
| ECM | Extracellular matrix components |
| PCDAI | Pediatric Crohn’s Disease Activity Index |
| PUCAI | Pediatric Ulcerative Colitis Activity Index |
References
- Kim, D.H.; Cheon, J.H. Pathogenesis of inflammatory bowel disease and recent advances in biologic therapies. Immune Netw. 2017, 17, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Petrey, A.C.; Motte, C.A. The extracellular matrix in IBD: A dynamic mediator of inflammation. Curr. Opin. Gastroenterol. 2017, 33, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, G.; Hritz, I.; Varga, M.Z.; Juhász, M.; Miheller, P.; Cierny, G.; Tulassay, Z.; Herszényi, L. The impact of matrix metalloproteinases and their tissue inhibitors in inflammatory bowel diseases. Dig. Dis. 2012, 30, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Vijay-Kumar, M.; Wang, L.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Matrix metalloproteinase-9-mediated tissue injury overrides the protective effect of matrix metalloproteinase-2 during colitis. Am. J. Physiol. Gastrointest. Liver. Physiol. 2009, 296, G175–G184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stallmach, A.; Chan, C.C.; Ecker, K.W.; Feifel, G.; Herbst, H.; Schuppan, D.; Zeitz, M. Comparable expression of matrix metalloproteinases 1 and 2 in pouchitis and ulcerative colitis. Gut 2000, 47, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matusiewicz, M.; Neubauer, K.; Mierzchala-Pasierb, M.; Gamian, A.; Krzystek-Korpacka, M. Matrix metalloproteinase-9: Its interplay with angiogenic factors in inflammatory bowel diseases. Dis. Markers 2014, 2014, 643645. [Google Scholar] [CrossRef]
- Bailey, C.J.; Hembry, R.M.; Alexander, A.; Irving, M.H.; Grant, M.E.; Shuttleworth, C.A. Distribution of the matrix metalloproteinases stromelysin, gelatinases A and B, and collagenase in Crohn’s disease and normal intestine. J. Clin. Pathol. 1994, 47, 113–116. [Google Scholar] [CrossRef]
- Meijer, M.J.; Mieremet-Ooms, M.A.; van der Zon, A.M.; van Duijn, W.; van Hogezand, R.A.; Sier, C.F.; Hommes, D.W.; Lamers, C.B.; Verspaget, H.W. Increased mucosal matrix metalloproteinase-1, -2, -3 and -9 activity in patients with inflammatory bowel disease and the relation with Crohn’s disease phenotype. Dig. Liver. Dis. 2007, 39, 733–739. [Google Scholar] [CrossRef]
- Lucafò, M.; Stankovic, B.; Kotur, N.; Di Silvestre, A.; Martelossi, S.; Ventura, A.; Zukic, B.; Pavlovic, S.; Decorti, G. Pharmacotranscriptomic biomarkers in glucocorticoid treatment of pediatric inflammatory bowel disease. Curr. Med. Chem. 2018, 25, 2855–2871. [Google Scholar] [CrossRef]
- Xu, D.; Jiang, Y.; Yang, L.; Hou, X.; Wang, J.; Gu, W.; Wang, X.; Liu, L.; Zhang, J.; Lu, H. Long noncoding RNAs expression profile and functional networks in rheumatoid arthritis. Oncotarget 2017, 8, 95280–95292. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.C.; Pan, H.F.; Leng, R.X.; Wang, D.G.; Li, X.P.; Li, X.M.; Ye, D.Q. Emerging role of long noncoding RNAs in autoimmune diseases. Autoimmun Rev. 2015, 14, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Wang, C.; Wang, D.; Yuan, H. LncRBA GSA5, up-regulated by ox-LDL, aggravates inflammatory response and MMP expression in THP-1 macrophages by acting like a sponge for miR-221. Exp. Cell Res. 2018, 369, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.R.; Schuliga, M.J.; Stewart, A.G. Pro-inflammatory mediators increase levels of the noncoding RNA GAS5 in airway smooth muscle and epithelial cells. Can. J. Physiol. Pharmacol. 2015, 93, 203–206. [Google Scholar] [CrossRef]
- Ito, I.; Asai, A.; Suzuki, S.; Kobayashi, M.; Suzuki, F. M2b macrophage polarization accompanied with reduction of long noncoding RNA GAS5. Biochem. Biophys. Res. Commun. 2017, 493, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, H.; Xiao, Y.; Tang, X.; Li, Y.; Han, Q.; Fu, J.; Yang, Y.; Zhu, Y. LncRNA GAS5 is a critical regulator of metastasis phenotype of melanoma cells and inhibits tumor growth in vivo. Oncol. Targets Ther. 2016, 9, 4075–4087. [Google Scholar] [CrossRef]
- Chen, L.; Yang, H.; Xiao, Y.; Tang, X.; Li, Y.; Han, Q.; Fu, J.; Yang, Y.; Zhu, Y. Lentiviral-mediated overexpression of long non-coding RNA GAS5 reduces invasion by mediating MMP2 expression and activity in human melanoma cells. Int. J. Oncol. 2016, 48, 1509–1518. [Google Scholar] [CrossRef]
- Koelink, P.J.; Overbeek, S.A.; Braber, S.; Morgan, M.E.; Henricks, P.A.; Abdul Roda, M.; Verspaget, H.W.; Wolfkamp, S.C.; te Velde, A.A.; Jones, C.W.; et al. Collagen degradation and neutrophilic infiltration: A vicious circle in inflammatory bowel disease. Gut 2014, 63, 578–587. [Google Scholar] [CrossRef]
- Kim, C.S.; Kang, J.H.; Cho, H.R.; Blankenship, T.N.; Erickson, K.L.; Kawada, T.; Yu, R. Potential involvement of CCL23 in atherosclerotic lesion formation/progression by the enhancement of chemotaxis, adhesion molecule expression, and MMP-2 release from monocytes. Inflamm Res. 2011, 60, 889–895. [Google Scholar] [CrossRef]
- Silva, F.A.; Rodrigues, B.L.; Ayrizono, M.L.; Leal, R.F. The immunological basis of inflammatory bowel disease. Gastroenterol. Res. Pract. 2016, 2016, 2097274. [Google Scholar] [CrossRef]
- Van Lint, P.; Libert, C. Chemokine and cytokine processing by matrix metalloproteinases and its effect on leukocyte migration and inflammation. J. Leukoc. Biol. 2007, 82, 1375–1381. [Google Scholar] [CrossRef] [Green Version]
- Buisson, A.; Vazeille, E.; Minet-Quinard, R.; Goutte, M.; Bouvier, D.; Goutorbe, F.; Pereira, B.; Barnich, N.; Bommelaer, G. Fecal matrix metalloprotease-9 and lipocalin-2 as biomarkers in detecting endoscopic activity in patients with inflammatory bowel diseases. J. Clin. Gastroenterol. 2018, 52, e53–e62. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, M.A.; Zurakowski, D.; Rufo, P.A.; Walker, T.R.; Fox, V.L.; Moses, M.A. Increased incidence of urinary matrix metalloproteinases as predictors of disease in pediatric patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Kofla-Dłubacz, A.; Matusiewicz, M.; Krzesiek, E.; Noga, L.; Iwańczak, B. Metalloproteinase-3 and -9 as novel markers in the evaluation of ulcerative colitis activity in children. Adv. Clin. Exp. Med. 2014, 23, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Yarani, R.; Mirza, A.H.; Kaur, S.; Pociot, F. The emerging role of lncRNAs in inflammatory bowel disease. Exp. Mol. Med. 2018, 50, 161. [Google Scholar] [CrossRef]
- Mayama, T.; Marr, A.K.; Kino, T. Differential expression of glucocorticoid receptor noncoding RNA repressor Gas5 in autoimmune and inflammatory diseases. Horm. Metab. Res. 2016, 48, 550–557. [Google Scholar] [CrossRef]
- Wu, G.C.; Hu, Y.; Guan, S.Y.; Ye, D.Q.; Pan, H.F. Differential plasma expression profiles of long non-Coding RNAs reveal potential biomarkers for systemic lupus erythematosus. Biomolecules 2019, 9, 206. [Google Scholar] [CrossRef]
- Lucafò, M.; Di Silvestre, A.; Romano, M.; Avian, A.; Antonelli, R.; Martelossi, S.; Naviglio, S.; Tommasini, A.; Stocco, G.; Ventura, A.; et al. Role of the long non-coding RNA growth arrest-specific 5 in Glucocorticoid response in children with inflammatory bowel disease. Basic. Clin. Pharmacol. Toxicol. 2018, 122, 87–93. [Google Scholar] [CrossRef]
- Turner, D.; Otley, A.R.; Mack, D.; Hyams, J.; de Bruijne, J.; Uusoue, K.; Walters, T.D.; Zachos, M.; Mamula, P.; Beaton, D.E.; et al. Development, validation, and evaluation of a pediatric ulcerative colitis activity index: A prospective multicenter study. Gastroenterology 2007, 133, 423–432. [Google Scholar] [CrossRef]
- Haberman, Y.; BenShoshan, M.; Di Segni, A.; Dexheimer, P.J.; Braun, T.; Weiss, B.; Walters, T.D.; Baldassano, R.N.; Noe, J.D.; Markowitz, J.; et al. Long ncRNA Landscape in the ileum of treatment-naive early-onset crohn disease. Inflamm. Bowel. Dis. 2018, 24, 346–360. [Google Scholar] [CrossRef]
- Carman, N.; Tomalty, D.; Church, P.C.; Mack, D.R.; Benchimol, E.I.; Otley, A.R.; Jacobson, K.; Huynh, H.Q.; DeBruyn, J.C.; El-Matary, W.; et al. Clinical disease activity and endoscopic severity correlate poorly in children newly diagnosed with Crohn’s disease. Gastrointest. Endosc. 2019, 89, 364–372. [Google Scholar] [CrossRef]
- Angrand, P.O.; Vennin, C.; Le Bourhis, X.; Adriaenssens, E. The role of long non-coding RNAs in genome formatting and expression. Front. Genet. 2015, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yu, Z.; Fang, X.; Liu, M.; Pu, Y.; Shao, Q.; Wang, D.; Zhao, X.; Huang, A.; Xiang, Z.; et al. LncRNA GAS5 inhibits microglial M2 polarization and exacerbates demyelination. EMBO Rep. 2017, 18, 1801–1816. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, J.; Huang, S.; Su, G.; Pi, G. LncRNA GAS5 overexpression reverses LPS-induced inflammatory injury and apoptosis through up-regulating KLF2 expression in ATDC5 chondrocytes. Cell Physiol. Biochem. 2018, 45, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Magro, F.; Langner, C.; Driessen, A.; Ensari, A.; Geboes, K.; Mantzaris, G.J.; Villanacci, V.; Becheanu, G.; Nunes, P.B.; Cathomas, G.; et al. European consensus on the histopathology of inflammatory bowel disease. J. Crohns Colitis. 2013, 7, 827–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzerini, M.; Villanacci, V.; Pellegrin, M.C.; Martelossi, S.; Magazzù, G.; Pellegrino, S.; Lucanto, M.C.; Barabino, A.; Calvi, A.; Arrigo, S.; et al. Endoscopic and Histologic Healing in Children With Inflammatory Bowel Diseases Treated With Thalidomide. Clin Gastroenterol. Hepatol. 2017, 15, 1382–1389. [Google Scholar] [CrossRef]
- Krzystek-Korpacka, M.; Diakowska, D.; Bania, J.; Gamian, A. Expression stability of common housekeeping genes is differently affected by bowel inflammation and cancer: Implications for finding suitable normalizers for inflammatory bowel disease studies. Inflamm. Bowel. Dis. 2014, 20, 1147–1156. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n) | 25 |
| Age (mean, range) | 12.5, 6.2–18 |
| Male (%) | 13 (52%) |
| CD (%) | 13 (52%) |
| UC (%) | 12 (48%) |
| Clinical score: | |
| PCDAI 1 (mean, range) | 30.6, 7.5–55 |
| PUCAI 2 (mean, range) | 30.1, 10–75 |
| Histologic findings: | |
| Inflammation score | Score (% of patients) |
| 0 (0%) | |
| 1 (4%) | |
| 2 (24%) | |
| 3 (36%) | |
| 4 (28%) | |
| 5 (8%) | |
| Architectural score | Score (% of patients) |
| 0 (0%) | |
| 1 (8%) | |
| 2 (28%) | |
| 3 (52%) | |
| 4 (12%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucafò, M.; Pugnetti, L.; Bramuzzo, M.; Curci, D.; Di Silvestre, A.; Marcuzzi, A.; Bergamo, A.; Martelossi, S.; Villanacci, V.; Bozzola, A.; et al. Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD. Int. J. Mol. Sci. 2019, 20, 5280. https://doi.org/10.3390/ijms20215280
Lucafò M, Pugnetti L, Bramuzzo M, Curci D, Di Silvestre A, Marcuzzi A, Bergamo A, Martelossi S, Villanacci V, Bozzola A, et al. Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD. International Journal of Molecular Sciences. 2019; 20(21):5280. https://doi.org/10.3390/ijms20215280
Chicago/Turabian StyleLucafò, Marianna, Letizia Pugnetti, Matteo Bramuzzo, Debora Curci, Alessia Di Silvestre, Annalisa Marcuzzi, Alberta Bergamo, Stefano Martelossi, Vincenzo Villanacci, Anna Bozzola, and et al. 2019. "Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD" International Journal of Molecular Sciences 20, no. 21: 5280. https://doi.org/10.3390/ijms20215280
APA StyleLucafò, M., Pugnetti, L., Bramuzzo, M., Curci, D., Di Silvestre, A., Marcuzzi, A., Bergamo, A., Martelossi, S., Villanacci, V., Bozzola, A., Cadei, M., De Iudicibus, S., Decorti, G., & Stocco, G. (2019). Long Non-Coding RNA GAS5 and Intestinal MMP2 and MMP9 Expression: A Translational Study in Pediatric Patients with IBD. International Journal of Molecular Sciences, 20(21), 5280. https://doi.org/10.3390/ijms20215280