Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives



Abstract
1. Introduction
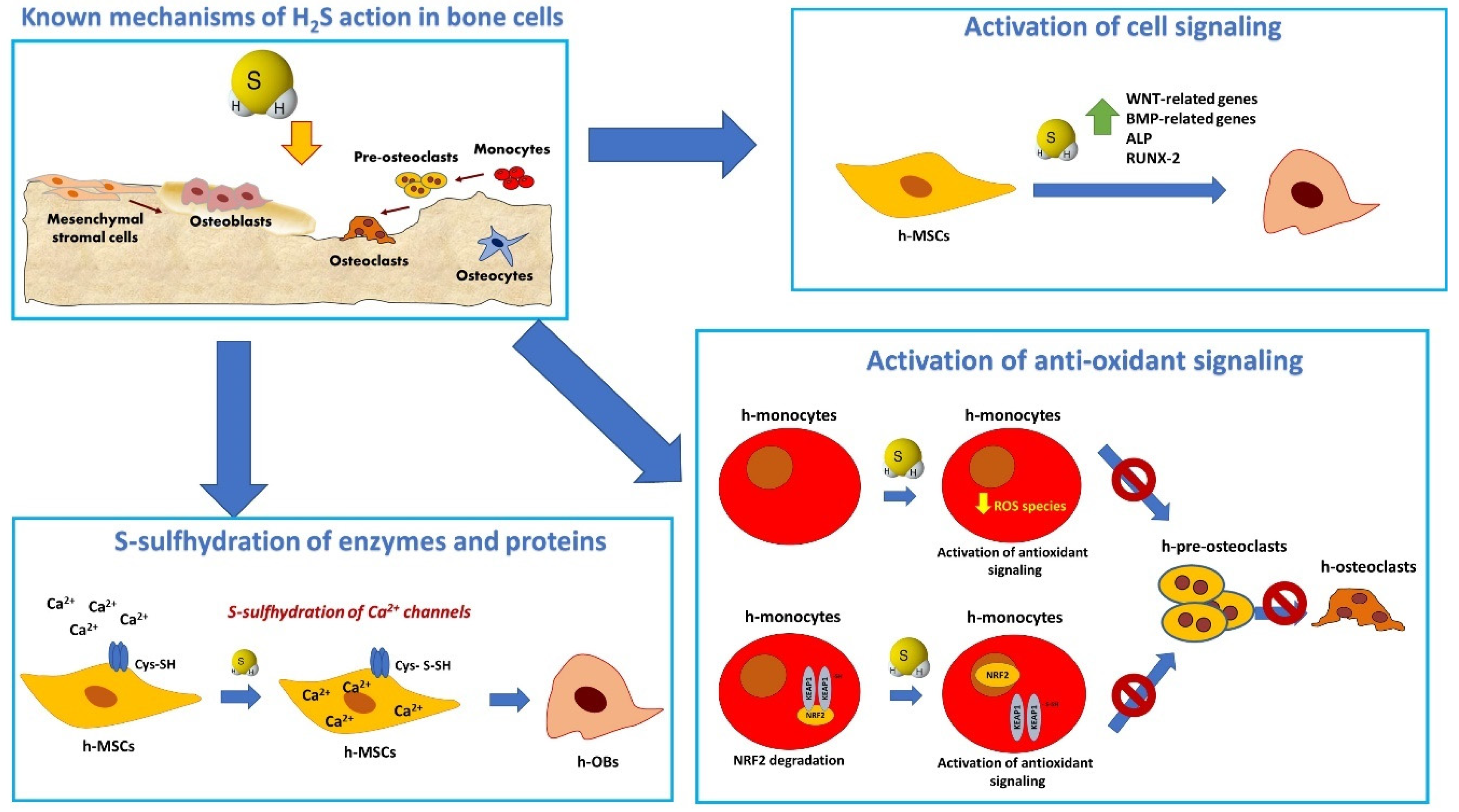
2. Bone Tissue Repair: H2S as a Suitable Biological Cue for Scaffold Functionalization
2.1. H2S Regulates Inflammation in Bone Tissue
2.2. H2S Regulates Migration and Survival of Cells Involved in Bone Repair
2.3. H2S Promotes the Expression of Angiogenic Factors in Bone Tissue
2.4. H2S Stimulates Bone Formation
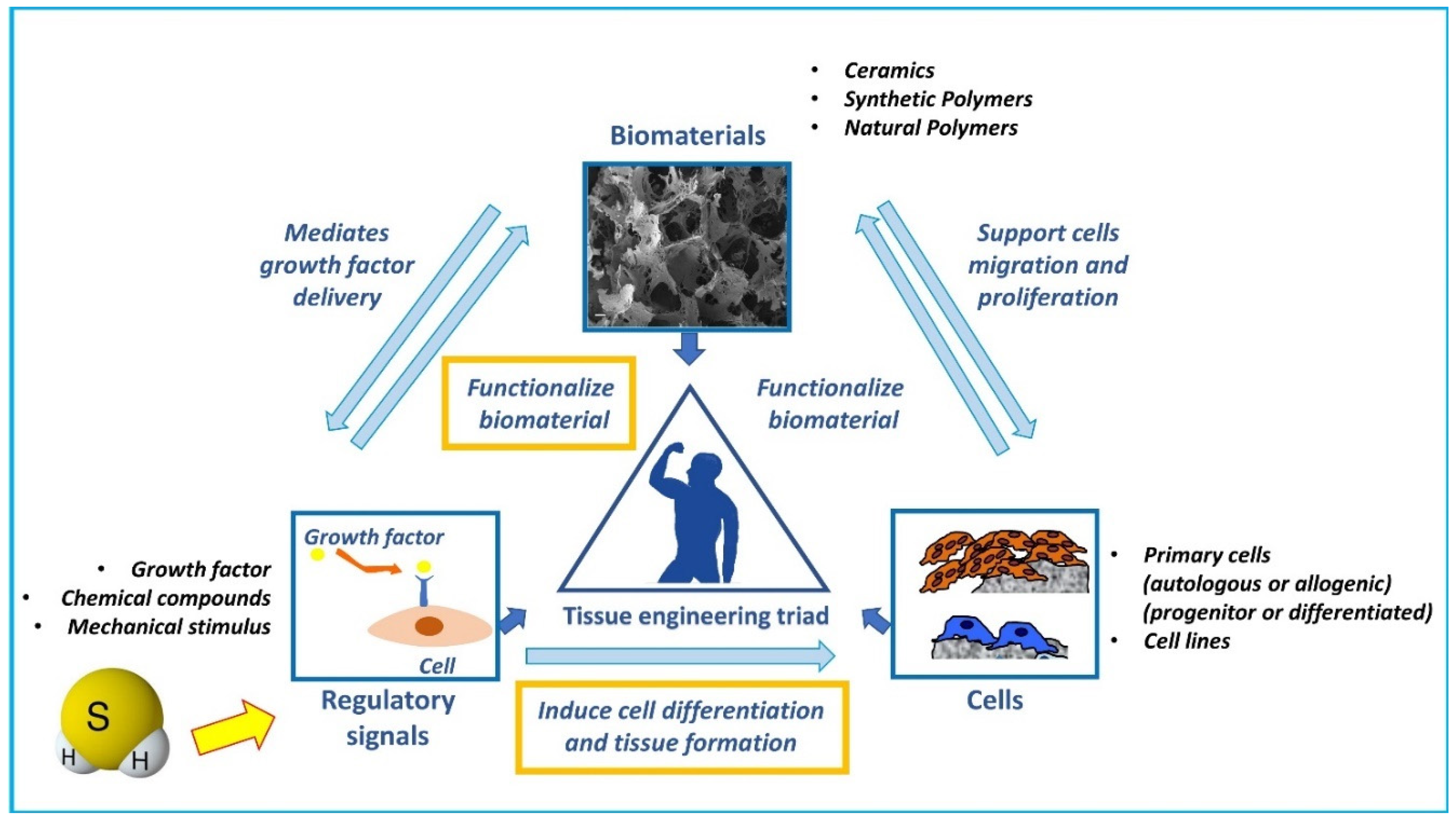
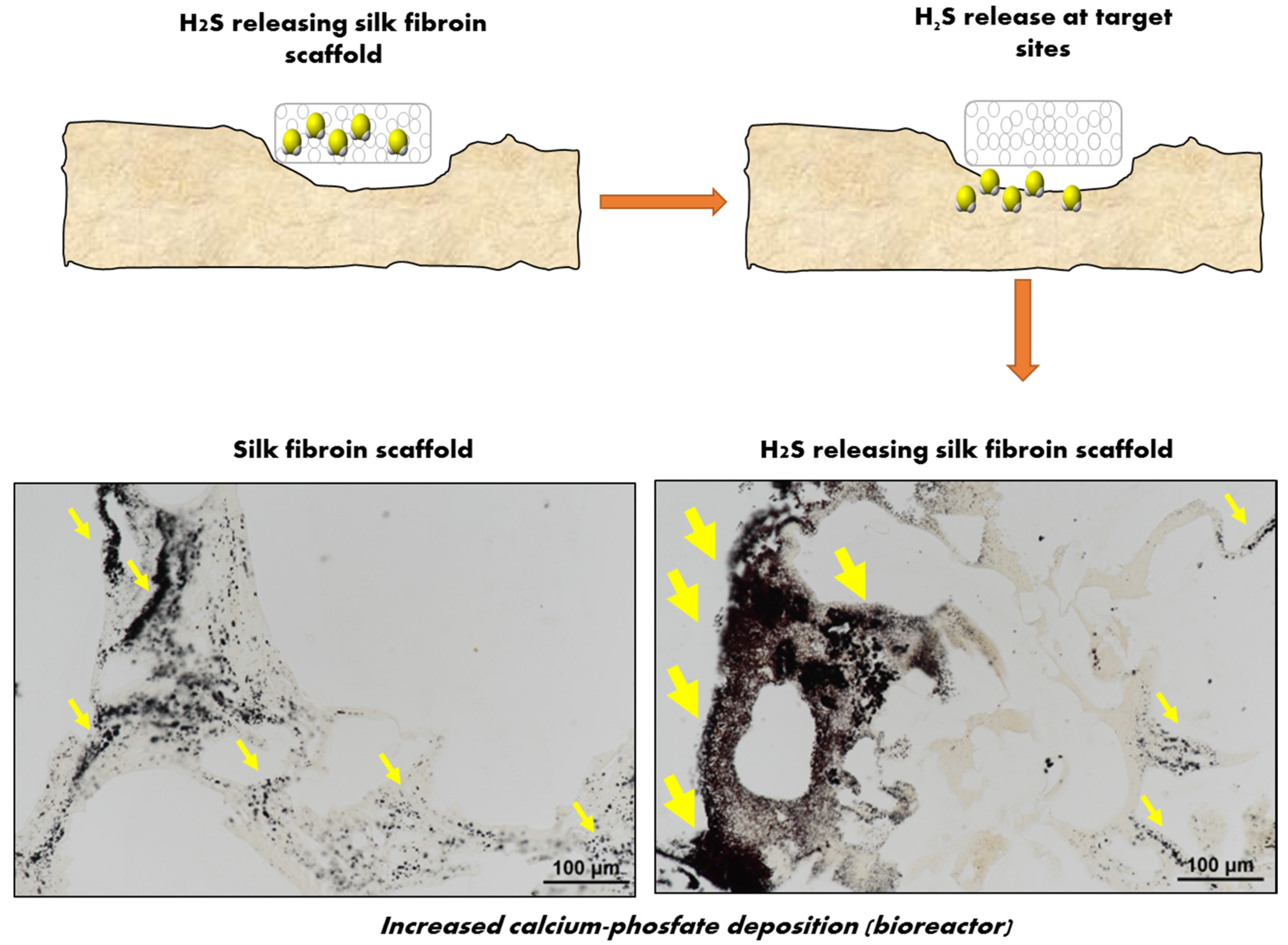
3. H2S-Releasing Scaffolds for Regenerative Medicine
4. Perspectives for H2S-Loaded Scaffolds for BTE
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Williamson, M.B.; Fromm, H.J. The Incorporation of Sulfur Amino Acids into the Proteins of Regenerating Wound Tissue. J. Biol. Chem. 1955, 212, 705–712. [Google Scholar]
- Abe, K.; Kimura, H. The Possible Role of Hydrogen Sulfide as an Endogenous Neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the Transsulfuration Pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Papapetropoulos, A. International Union of Basic and Clinical Pharmacology. CII: Pharmacological Modulation of H2S Levels: H2S Donors and H2S Biosynthesis Inhibitors. Pharmacol. Rev. 2017, 69, 497–564. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.T.; Chen, L.; Xu, S.; Day, J.J.; Li, X.; Xian, M. Recent Development of Hydrogen Sulfide Releasing/Stimulating Reagents and Their Potential Applications in Cancer and Glycometabolic Disorders. Front. Pharmacol. 2017, 8, 664. [Google Scholar] [CrossRef]
- Grabowski, P. Physiology of Bone. Endocr. Dev. 2015, 28, 33–55. [Google Scholar] [PubMed]
- Guntur, A.R.; Rosen, C.J. Bone as an Endocrine Organ. Endocr. Pract. 2012, 18, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone Remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Seeman, E. Bone Remodelling: Its Local Regulation and the Emergence of Bone Fragility. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 701–722. [Google Scholar] [CrossRef]
- Sommerfeldt, D.W.; Rubin, C.T. Biology of Bone and How It Orchestrates the Form and Function of the Skeleton. Eur. Spine J. 2001, 10 (Suppl. 2), S86–S95. [Google Scholar]
- Winkler, T.; Sass, F.A.; Duda, G.N.; Schmidt-Bleek, K. A Review of Biomaterials in Bone Defect Healing, Remaining Shortcomings and Future Opportunities for Bone Tissue Engineering: The Unsolved Challenge. Bone Jt. Res. 2018, 7, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Nauth, A.; Schemitsch, E.; Norris, B.; Nollin, Z.; Watson, J.T. Critical-Size Bone Defects: Is There a Consensus for Diagnosis and Treatment? J. Orthop. Trauma 2018, 32 (Suppl. 1), S7–S11. [Google Scholar] [CrossRef] [PubMed]
- Curry, A.S.; Pensa, N.W.; Barlow, A.M.; Bellis, S.L. Taking Cues from the Extracellular Matrix to Design Bone-Mimetic Regenerative Scaffolds. Matrix Biol. 2016, 52–54, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Grassi, F.; Tyagi, A.M.; Calvert, J.W.; Gambari, L.; Walker, L.D.; Yu, M.; Robinson, J.; Li, J.Y.; Lisignoli, G.; Vaccaro, C.; et al. Hydrogen Sulfide Is a Novel Regulator of Bone Formation Implicated in the Bone Loss Induced by Estrogen Deficiency. J. Bone Miner. Res. 2016, 31, 949–963. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, R.; Liu, X.; Zhou, Y.; Qu, C.; Kikuiri, T.; Wang, S.; Zandi, E.; Du, J.; Ambudkar, I.S.; et al. Hydrogen Sulfide Maintains Mesenchymal Stem Cell Function and Bone Homeostasis Via Regulation of Ca(2+) Channel Sulfhydration. Cell Stem Cell 2014, 15, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; Kelly, K.E.; Voor, M.J.; Metreveli, N.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Promotes Bone Homeostasis by Balancing Inflammatory Cytokine Signaling in Cbs-Deficient Mice through an Epigenetic Mechanism. Sci. Rep. 2018, 8, 15226. [Google Scholar] [CrossRef] [PubMed]
- Ozcivici, E.; Luu, Y.K.; Adler, B.; Qin, Y.X.; Rubin, J.; Judex, S.; Rubin, C.T. Mechanical Signals as Anabolic Agents in Bone. Nat. Rev. Rheumatol. 2010, 6, 50–59. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T.A. The Biology of Fracture Healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Colnot, C.; Thompson, Z.; Miclau, T.; Werb, Z.; Helms, J.A. Altered Fracture Repair in the Absence of Mmp9. Development 2003, 130, 4123–4133. [Google Scholar] [CrossRef]
- Perez, J.R.; Kouroupis, D.; Li, D.J.; Best, T.M.; Kaplan, L.; Correa, D. Tissue Engineering and Cell-Based Therapies for Fractures and Bone Defects. Front. Bioeng. Biotechnol. 2018, 6, 105. [Google Scholar] [CrossRef]
- Hak, D.J.; Fitzpatrick, D.; Bishop, J.A.; Marsh, J.L.; Tilp, S.; Schnettler, R.; Simpson, H.; Alt, V. Delayed Union and Nonunions: Epidemiology, Clinical Issues, and Financial Aspects. Injury 2014, 45 (Suppl. 2), S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Verrier, S.; Alini, M.; Alsberg, E.; Buchman, S.R.; Kelly, D.; Laschke, M.W.; Menger, M.D.; Murphy, W.L.; Stegemann, J.P.; Schutz, M.; et al. Tissue Engineering and Regenerative Approaches to Improving the Healing of Large Bone Defects. Eur. Cell Mater. 2016, 32, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G.; Moghaddam, A. Allograft Bone Matrix Versus Synthetic Bone Graft Substitutes. Injury 2011, 42 (Suppl. 2), S16–S21. [Google Scholar] [CrossRef]
- Baldwin, P.; Li, D.J.; Auston, D.A.; Mir, H.S.; Yoon, R.S.; Koval, K.J. Autograft, Allograft, and Bone Graft Substitutes: Clinical Evidence and Indications for Use in the Setting of Orthopaedic Trauma Surgery. J. Orthop. Trauma 2019, 33, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Calcei, J.G.; Rodeo, S.A. Orthobiologics for Bone Healing. Clin. Sports Med. 2019, 38, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, H.; Zhao, Y.; Luo, J.; Yang, L.; Liu, W.; He, Q. Functionalized Cell-Free Scaffolds for Bone Defect Repair Inspired by Self-Healing of Bone Fractures: A Review and New Perspectives. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 98, 1241–1251. [Google Scholar] [CrossRef]
- Lopes, D.; Martins-Cruz, C.; Oliveira, M.B.; Mano, J.F. Bone Physiology as Inspiration for Tissue Regenerative Therapies. Biomaterials 2018, 185, 240–275. [Google Scholar] [CrossRef]
- Bigham-Sadegh, A.; Oryan, A. Selection of Animal Models for Pre-Clinical Strategies in Evaluating the Fracture Healing, Bone Graft Substitutes and Bone Tissue Regeneration and Engineering. Connect. Tissue Res. 2015, 56, 175–194. [Google Scholar] [CrossRef]
- Peric, M.; Dumic-Cule, I.; Grcevic, D.; Matijasic, M.; Verbanac, D.; Paul, R.; Grgurevic, L.; Trkulja, V.; Bagi, C.M.; Vukicevic, S. The Rational Use of Animal Models in the Evaluation of Novel Bone Regenerative Therapies. Bone 2015, 70, 73–86. [Google Scholar] [CrossRef]
- Loi, F.; Cordova, L.A.; Pajarinen, J.; Lin, T.H.; Yao, Z.; Goodman, S.B. Inflammation, Fracture and Bone Repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Wullschleger, M.E.; Alexander, K.A.; Wu, A.C.; Millard, S.M.; Kaur, S.; Maugham, M.L.; Gregory, L.S.; Steck, R.; Pettit, A.R. Fracture Healing Via Periosteal Callus Formation Requires Macrophages for Both Initiation and Progression of Early Endochondral Ossification. Am. J. Pathol. 2014, 184, 3192–3204. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tan, J.; Martino, M.M.; Lui, K.O. Regulatory T-Cells: Potential Regulator of Tissue Repair and Regeneration. Front. Immunol. 2018, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, R.; Shi, S. Systemic Infusion of Mesenchymal Stem Cells Improves Cell-Based Bone Regeneration Via Upregulation of Regulatory T Cells. Tissue Eng. Part A 2015, 21, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Colburn, N.T.; Zaal, K.J.; Wang, F.; Tuan, R.S. A Role for Gamma/Delta T Cells in a Mouse Model of Fracture Healing. Arthritis Rheum. 2009, 60, 1694–1703. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Shi, S. The Role of Recipient T Cells in Mesenchymal Stem Cell-Based Tissue Regeneration. Int. J. Biochem. Cell Biol. 2012, 44, 2044–2050. [Google Scholar] [CrossRef]
- Sadtler, K.; Allen, B.W.; Estrellas, K.; Housseau, F.; Pardoll, D.M.; Elisseeff, J.H. The Scaffold Immune Microenvironment: Biomaterial-Mediated Immune Polarization in Traumatic and Nontraumatic Applications. Tissue Eng. Part A 2017, 23, 1044–1053. [Google Scholar] [CrossRef]
- Hegde, A.; Bhatia, M. Hydrogen Sulfide in Inflammation: Friend or Foe? Inflamm. Allergy Drug Targets 2011, 10, 118–122. [Google Scholar] [CrossRef]
- Fox, B.; Schantz, J.T.; Haigh, R.; Wood, M.E.; Moore, P.K.; Viner, N.; Spencer, J.P.; Winyard, P.G.; Whiteman, M. Inducible Hydrogen Sulfide Synthesis in Chondrocytes and Mesenchymal Progenitor Cells: Is H2S a Novel Cytoprotective Mediator in the Inflamed Joint? J. Cell Mol. Med. 2012, 16, 896–910. [Google Scholar] [CrossRef]
- Gambari, L.; Lisignoli, G.; Cattini, L.; Manferdini, C.; Facchini, A.; Grassi, F. Sodium Hydrosulfide Inhibits the Differentiation of Osteoclast Progenitor Cells Via Nrf2-Dependent Mechanism. Pharmacol. Res. 2014, 87, 99–112. [Google Scholar] [CrossRef]
- Ezeriņa, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-Acetyl Cysteine Functions as a Fast-Acting Antioxidant by Triggering Intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 2018, 25, 447–459. [Google Scholar] [CrossRef]
- Cao, S.; Zhu, P.; Yu, X.; Chen, J.; Li, J.; Yan, F.; Wang, L.; Yu, J.; Chen, G. Hydrogen Sulfide Attenuates Brain Edema in Early Brain Injury after Subarachnoid Hemorrhage in Rats: Possible Involvement of Mmp-9 Induced Blood-Brain Barrier Disruption and Aqp4 Expression. Neurosci. Lett. 2016, 621, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, R.C.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen Sulfide Is an Endogenous Modulator of Leukocyte-Mediated Inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Shen, X.; Whiteman, M.; Xin, H.; Shen, Y.; Xin, X.; Moore, P.K.; Zhu, Y.Z. Hydrogen Sulfide Mitigates Myocardial Infarction Via Promotion of Mitochondrial Biogenesis-Dependent M2 Polarization of Macrophages. Antioxid. Redox Signal. 2016, 25, 268–281. [Google Scholar] [CrossRef]
- Zhou, X.; Chu, X.; Xin, D.; Li, T.; Bai, X.; Qiu, J.; Yuan, H.; Liu, D.; Wang, D.; Wang, Z. L-Cysteine-Derived H2S Promotes Microglia M2 Polarization Via Activation of the Ampk Pathway in Hypoxia-Ischemic Neonatal Mice. Front. Mol. Neurosci. 2019, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, A.; Zhou, Y.; Zheng, S.; Yang, Y.; An, Y.; Xu, K.; Wang, Q. Novel H2S-Releasing Hydrogel for Wound Repair Via in Situ Polarization of M2 Macrophages. Biomaterials 2019, 222, 119398. [Google Scholar] [CrossRef]
- Brancaleone, V.; Mitidieri, E.; Flower, R.J.; Cirino, G.; Perretti, M. Annexin A1 Mediates Hydrogen Sulfide Properties in the Control of Inflammation. J. Pharmacol. Exp. Ther. 2014, 351, 96–104. [Google Scholar] [CrossRef]
- Herrera, B.S.; Coimbra, L.S.; da Silva, A.R.; Teixeira, S.A.; Costa, S.K.; Wallace, J.L.; Spolidorio, L.C.; Muscara, M.N. The H2S-Releasing Naproxen Derivative, Atb-346, Inhibits Alveolar Bone Loss and Inflammation in Rats with Ligature-Induced Periodontitis. Med. Gas. Res. 2015, 5, 4. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; D’Amico, R.; Militi, A.; Oteri, G.; Wallace, J.L.; Di Paola, R.; Cuzzocrea, S. Anti-Inflammatory Effect of Atb-352, a H2S -Releasing Ketoprofen Derivative, on Lipopolysaccharide-Induced Periodontitis in Rats. Pharmacol. Res. 2018, 132, 220–231. [Google Scholar] [CrossRef]
- Xiao, Z.; Bonnard, T.; Shakouri-Motlagh, A.; Wylie, R.A.L.; Collins, J.; White, J.; Heath, D.E.; Hagemeyer, C.E.; Connal, L.A. Triggered and Tunable Hydrogen Sulfide Release from Photogenerated Thiobenzaldehydes. Chemistry 2017, 23, 11294–11300. [Google Scholar] [CrossRef]
- Benedetti, F.; Curreli, S.; Krishnan, S.; Davinelli, S.; Cocchi, F.; Scapagnini, G.; Gallo, R.C.; Zella, D. Anti-Inflammatory Effects of H2S During Acute Bacterial Infection: A Review. J. Transl. Med. 2017, 15, 100. [Google Scholar] [CrossRef]
- Miller, T.W.; Wang, E.A.; Gould, S.; Stein, E.V.; Kaur, S.; Lim, L.; Amarnath, S.; Fowler, D.H.; Roberts, D.D. Hydrogen Sulfide Is an Endogenous Potentiator of T Cell Activation. J. Biol. Chem. 2012, 287, 4211–4221. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Liu, Y.; Shi, S. Hydrogen Sulfide Regulates Homeostasis of Mesenchymal Stem Cells and Regulatory T Cells. J. Dent. Res. 2016, 95, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Qu, C.; Zhou, Y.; Konkel, J.E.; Shi, S.; Liu, Y.; Chen, C.; Liu, S.; Liu, D.; Chen, Y.; et al. Hydrogen Sulfide Promotes Tet1- and Tet2-Mediated Foxp3 Demethylation to Drive Regulatory T Cell Differentiation and Maintain Immune Homeostasis. Immunity 2015, 43, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Akter, F. The Role of Hydrogen Sulfide in Burns. Burns 2016, 42, 519–525. [Google Scholar] [CrossRef]
- Zhao, H.; Lu, S.; Chai, J.; Zhang, Y.; Ma, X.; Chen, J.; Guan, Q.; Wan, M.; Liu, Y. Hydrogen Sulfide Improves Diabetic Wound Healing in Ob/Ob Mice Via Attenuating Inflammation. J. Diabetes Complicat. 2017, 31, 1363–1369. [Google Scholar] [CrossRef]
- Wang, G.; Li, W.; Chen, Q.; Jiang, Y.; Lu, X.; Zhao, X. Hydrogen Sulfide Accelerates Wound Healing in Diabetic Rats. Int. J. Clin. Exp. Pathol. 2015, 8, 5097–5104. [Google Scholar]
- Seebach, C.; Henrich, D.; Wilhelm, K.; Barker, J.H.; Marzi, I. Endothelial Progenitor Cells Improve Directly and Indirectly Early Vascularization of Mesenchymal Stem Cell-Driven Bone Regeneration in a Critical Bone Defect in Rats. Cell Transplant. 2012, 21, 1667–1677. [Google Scholar] [CrossRef]
- Briasoulis, A.; Tousoulis, D.; Antoniades, C.; Papageorgiou, N.; Stefanadis, C. The Role of Endothelial Progenitor Cells in Vascular Repair after Arterial Injury and Atherosclerotic Plaque Development. Cardiovasc. Ther. 2011, 29, 125–139. [Google Scholar] [CrossRef]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef]
- Pankajakshan, D.; Agrawal, D.K. Mesenchymal Stem Cell Paracrine Factors in Vascular Repair and Regeneration. J. Biomed. Technol. Res. 2014, 1. [Google Scholar] [CrossRef]
- Hu, Q.; Ke, X.; Zhang, T.; Chen, Y.; Huang, Q.; Deng, B.; Xie, S.; Wang, J.; Nie, R. Hydrogen Sulfide Improves Vascular Repair by Promoting Endothelial Nitric Oxide Synthase-Dependent Mobilization of Endothelial Progenitor Cells. J. Hypertens. 2019, 37, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Zou, J.; Hu, Q.; Wang, X.; Hu, C.; Yang, R.; Liang, J.; Shu, X.; Nie, R.; Peng, C. Hydrogen Sulfide-Preconditioning of Human Endothelial Progenitor Cells Transplantation Improves Re-Endothelialization in Nude Mice with Carotid Artery Injury. Cell Physiol. Biochem. 2017, 43, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Garikipati, V.N.; Nickoloff, E.; Wang, C.; Polhemus, D.J.; Zhou, J.; Benedict, C.; Khan, M.; Verma, S.K.; Rabinowitz, J.E.; et al. Restoration of Hydrogen Sulfide Production in Diabetic Mice Improves Reparative Function of Bone Marrow Cells. Circulation 2016, 134, 1467–1483. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hua, Y.; Qi, Y.; Meng, G.; Yang, S. Exogenous Hydrogen Sulphide Supplement Accelerates Skin Wound Healing Via Oxidative Stress Inhibition and Vascular Endothelial Growth Factor Enhancement. Exp. Dermatol. 2019, 28, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Xin, X.; Xin, H.; Shen, X.; Zhu, Y.Z. Hydrogen Sulfide Recruits Macrophage Migration by Integrin Beta1-Src-Fak/Pyk2-Rac Pathway in Myocardial Infarction. Sci. Rep. 2016, 6, 22363. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Li, C.S.; Wang, C.M.; Xie, Y.J.; Wang, A.L. Cse/H2S System Protects Mesenchymal Stem Cells from Hypoxia and Serum Deprivationinduced Apoptosis Via Mitochondrial Injury, Endoplasmic Reticulum Stress and Pi3k/Akt Activation Pathways. Mol. Med. Rep. 2015, 12, 2128–2134. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, H.; Kong, G.; Shim, W.; Zhang, G. Hydrogen Sulfide Augments the Proliferation and Survival of Human Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells through Inhibition of Bkca. Cytotherapy 2013, 15, 1395–1405. [Google Scholar] [CrossRef]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular Biology of Fracture Healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture Healing: Mechanisms and Interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef]
- Utvag, S.E.; Grundnes, O.; Rindal, D.B.; Reikeras, O. Influence of Extensive Muscle Injury on Fracture Healing in Rat Tibia. J. Orthop. Trauma 2003, 17, 430–435. [Google Scholar] [CrossRef]
- Hu, K.; Olsen, B.R. Osteoblast-Derived Vegf Regulates Osteoblast Differentiation and Bone Formation During Bone Repair. J. Clin. Investig. 2016, 126, 509–526. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Pyriochou, A.; Altaany, Z.; Yang, G.; Marazioti, A.; Zhou, Z.; Jeschke, M.G.; Branski, L.K.; Herndon, D.N.; Wang, R.; et al. Hydrogen Sulfide Is an Endogenous Stimulator of Angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21972–21977. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.J.; Wang, M.J.; Moore, P.K.; Jin, H.M.; Yao, T.; Zhu, Y.C. The Novel Proangiogenic Effect of Hydrogen Sulfide Is Dependent on Akt Phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Katsouda, A.; Bibli, S.I.; Pyriochou, A.; Szabo, C.; Papapetropoulos, A. Regulation and Role of Endogenously Produced Hydrogen Sulfide in Angiogenesis. Pharmacol. Res. 2016, 113, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Cai, W.J.; Zhu, Y.C. Mechanisms of Angiogenesis: Role of Hydrogen Sulphide. Clin. Exp. Pharmacol. Physiol. 2010, 37, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Cai, W.J.; Li, N.; Ding, Y.J.; Chen, Y.; Zhu, Y.C. The Hydrogen Sulfide Donor Nahs Promotes Angiogenesis in a Rat Model of Hind Limb Ischemia. Antioxid. Redox Signal. 2010, 12, 1065–1077. [Google Scholar] [CrossRef]
- Gambari, L.; Amore, E.; Raggio, R.; Bonani, W.; Barone, M.; Lisignoli, G.; Grigolo, B.; Motta, A.; Grassi, F. Hydrogen Sulfide-Releasing Silk Fibroin Scaffold for Bone Tissue Engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 102, 471–482. [Google Scholar] [CrossRef]
- Liu, X.; Pan, L.; Zhuo, Y.; Gong, Q.; Rose, P.; Zhu, Y. Hypoxia-Inducible Factor-1alpha Is Involved in the Pro-Angiogenic Effect of Hydrogen Sulfide under Hypoxic Stress. Biol. Pharm. Bull. 2010, 33, 1550–1554. [Google Scholar] [CrossRef]
- Majumder, A.; Singh, M.; George, A.K.; Behera, J.; Tyagi, N.; Tyagi, S.C. Hydrogen Sulfide Improves Postischemic Neoangiogenesis in the Hind Limb of Cystathionine-Beta-Synthase Mutant Mice Via Ppar-Gamma/Vegf Axis. Physiol. Rep. 2018, 6, e13858. [Google Scholar] [CrossRef]
- Gambari, L.; Lisignoli, G.; Gabusi, E.; Manferdini, C.; Paolella, F.; Piacentini, A.; Grassi, F. Distinctive Expression Pattern of Cystathionine-Beta-Synthase and Cystathionine-Gamma-Lyase Identifies Mesenchymal Stromal Cells Transition to Mineralizing Osteoblasts. J. Cell. Physiol. 2017, 232, 3574–3585. [Google Scholar] [CrossRef]
- Zheng, Y.; Liao, F.; Lin, X.; Zheng, F.; Fan, J.; Cui, Q.; Yang, J.; Geng, B.; Cai, J. Cystathionine Gamma-Lyase-Hydrogen Sulfide Induces Runt-Related Transcription Factor 2 Sulfhydration, Thereby Increasing Osteoblast Activity to Promote Bone Fracture Healing. Antioxid. Redox Signal. 2017, 27, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, E.; Penolazzi, L.; Angelozzi, M.; Grassi, F.; Gambari, L.; Lisignoli, G.; de Bonis, P.; Cavallo, M.; Piva, R. The Expression of Cystathionine Gamma-Lyase Is Regulated by Estrogen Receptor Alpha in Human Osteoblasts. Oncotarget 2017, 8, 101686–101696. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhai, Y.; Behera, J.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Attenuates Homocysteine-Induced Osteoblast Dysfunction by Inhibiting Mitochondrial Toxicity. J. Cell. Physiol. 2019, 234, 18602–18614. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Liu, Y.; Xiao, T.H.; Jiang, W.; Lin, B.W.; Zhang, X.M.; Lin, Y.M.; Xu, Z.S. Gyy4137 Stimulates Osteoblastic Cell Proliferation and Differentiation Via an Erk1/2-Dependent Anti-Oxidant Mechanism. Am. J. Transl. Res. 2017, 9, 1183–1192. [Google Scholar] [PubMed]
- Mo, S.; Hua, Y. Cystathionine Gamma Lyase-H2S Contributes to Osteoclastogenesis During Bone Remodeling Induced by Mechanical Loading. Biochem. Biophys. Res. Commun. 2018, 501, 471–477. [Google Scholar] [CrossRef]
- Liu, F.; Wen, F.; He, D.; Liu, D.; Yang, R.; Wang, X.; Yan, Y.; Liu, Y.; Kou, X.; Zhou, Y. Force-Induced H2S by Pdlscs Modifies Osteoclastic Activity During Tooth Movement. J. Dent. Res. 2017, 96, 694–702. [Google Scholar] [CrossRef]
- Lee, S.K.; Chung, J.H.; Choi, S.C.; Auh, Q.S.; Lee, Y.M.; Lee, S.I.; Kim, E.C. Sodium Hydrogen Sulfide Inhibits Nicotine and Lipopolysaccharide-Induced Osteoclastic Differentiation and Reversed Osteoblastic Differentiation in Human Periodontal Ligament Cells. J. Cell Biochem. 2013, 114, 1183–1193. [Google Scholar] [CrossRef]
- Cen, S.D.; Yu, W.B.; Ren, M.M.; Chen, L.J.; Sun, C.F.; Ye, Z.L.; Deng, H.; Hu, R.D. Endogenous Hydrogen Sulfide Is Involved in Osteogenic Differentiation in Human Periodontal Ligament Cells. Arch. Oral. Biol. 2016, 68, 1–8. [Google Scholar] [CrossRef]
- Jiang, Z.; Hua, Y. Hydrogen Sulfide Promotes Osteogenic Differentiation of Human Periodontal Ligament Cells Via P38-Mapk Signaling Pathway under Proper Tension Stimulation. Arch. Oral. Biol. 2016, 72, 8–13. [Google Scholar] [CrossRef]
- Pu, H.; Hua, Y. Hydrogen Sulfide Regulates Bone Remodeling and Promotes Orthodontic Tooth Movement. Mol. Med. Rep. 2017, 16, 9415–9422. [Google Scholar] [CrossRef]
- Xu, Z.S.; Dai, F.; Chen, J.; Lv, M.; Cheng, J.W.; Zhang, X.M.; Lin, B.W. Experimental Research into the Potential Therapeutic Effect of Gyy4137 on Ovariectomy-Induced Osteoporosis. Cell. Mol. Biol. Lett. 2018, 23, 47. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; George, A.K.; Voor, M.J.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Epigenetically Mitigates Bone Loss through Opg/Rankl Regulation During Hyperhomocysteinemia in Mice. Bone 2018, 114, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Lewis, J.L.; Lin, H.S.; Kuo, L.T.; Mao, S.W.; Tai, Y.S.; Chang, M.S.; Ables, G.P.; Perrone, C.E.; Yang, R.S. A Methionine-Restricted Diet and Endurance Exercise Decrease Bone Mass and Extrinsic Strength but Increase Intrinsic Strength in Growing Male Rats. J. Nutr. 2014, 144, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hao, D.; Zhang, H.; Liu, B.; Yang, M.; He, B. Treatment with Hydrogen Sulfide Attenuates Sublesional Skeletal Deterioration Following Motor Complete Spinal Cord Injury in Rats. Osteoporos. Int. 2017, 28, 687–695. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, Y.; Lu, K.; Zhang, H.; Fan, X. Gyy4137 Promotes Bone Formation in a Rabbit Distraction Osteogenesis Model: A Preliminary Report. J. Oral Maxillofac. Surg. 2015, 73, 732. [Google Scholar] [CrossRef]
- Jiang, X.W.; Zhang, Y.; Cheng, Y.Z.; Fan, X.S.; Deng, X.; Peng, H.Y. The Expression of Endogenous Hydrogen Sulfide Signal During Distraction Osteogenesis in a Rabbit Model. Int. J. Oral Maxillofac. Surg. 2018, 47, 262–267. [Google Scholar] [CrossRef]
- Hu, K.; Olsen, B.R. The Roles of Vascular Endothelial Growth Factor in Bone Repair and Regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef]
- Miclau, K.R.; Brazina, S.A.; Bahney, C.S.; Hankenson, K.D.; Hunt, T.K.; Marcucio, R.S.; Miclau, T. Stimulating Fracture Healing in Ischemic Environments: Does Oxygen Direct Stem Cell Fate During Fracture Healing? Front. Cell Dev. Biol. 2017, 5, 45. [Google Scholar] [CrossRef]
- Runyan, C.M.; Gabrick, K.S. Biology of Bone Formation, Fracture Healing, and Distraction Osteogenesis. J. Craniofac. Surg. 2017, 28, 1380–1389. [Google Scholar] [CrossRef]
- Coates, B.A.; McKenzie, J.A.; Buettmann, E.G.; Liu, X.; Gontarz, P.M.; Zhang, B.; Silva, M.J. Transcriptional Profiling of Intramembranous and Endochondral Ossification after Fracture in Mice. Bone 2019, 127, 577–591. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cullinane, D.M.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Fracture Healing as a Post-Natal Developmental Process: Molecular, Spatial, and Temporal Aspects of Its Regulation. J. Cell Biochem. 2003, 88, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.K.; Kim, H.N.; Bhang, S.H.; Shin, J.Y.; Han, J.; La, W.G.; Jeong, G.J.; Kang, S.; Lee, J.R.; Oh, J.; et al. Enhanced Bone Repair by Guided Osteoblast Recruitment Using Topographically Defined Implant. Tissue Eng. Part A 2016, 22, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Lee, D.H.; Jeon, J.; Oh, S.H.; Yang, H.S. Topographically Defined, Biodegradable Nanopatterned Patches to Regulate Cell Fate and Acceleration of Bone Regeneration. ACS Appl. Mater. Interfaces 2018, 10, 38780–38790. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Ammann, P.; Boivin, G.; Rey, C. Mechanisms of Action and Therapeutic Potential of Strontium in Bone. Calcified Tissue Int. 2001, 69, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Zreiqat, H.; Howlett, C.R.; Zannettino, A.; Evans, P.; Schulze-Tanzil, G.; Knabe, C.; Shakibaei, M. Mechanisms of Magnesium-Stimulated Adhesion of Osteoblastic Cells to Commonly Used Orthopaedic Implants. J. Biomed. Mater. Res. 2002, 62, 175–184. [Google Scholar] [CrossRef]
- Quinlan, E.; Partap, S.; Azevedo, M.M.; Jell, G.; Stevens, M.M.; O’Brien, F.J. Hypoxia-Mimicking Bioactive Glass/Collagen Glycosaminoglycan Composite Scaffolds to Enhance Angiogenesis and Bone Repair. Biomaterials 2015, 52, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, E.M. Silicon: A Requirement in Bone Formation Independent of Vitamin D1. Calcified Tissue Int. 1981, 33, 27–34. [Google Scholar] [CrossRef]
- Wu, J.; Li, Y.; He, C.; Kang, J.; Ye, J.; Xiao, Z.; Zhu, J.; Chen, A.; Feng, S.; Li, X.; et al. Novel H2S Releasing Nanofibrous Coating for In Vivo Dermal Wound Regeneration. ACS Appl. Mater. Interfaces 2016, 8, 27474–27481. [Google Scholar] [CrossRef]
- Goren, I.; Kohler, Y.; Aglan, A.; Pfeilschifter, J.; Beck, K.F.; Frank, S. Increase of Cystathionine-Gamma-Lyase (Cse) During Late Wound Repair: Hydrogen Sulfide Triggers Cytokeratin 10 Expression in Keratinocytes. Nitric Oxide 2019, 87, 31–42. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Surmiak, M.; Sliwowski, Z.; Wierdak, M.; Kwiecien, S.; Chmura, A.; Brzozowski, T. Cross-Talk between Hydrogen Sulfide and Carbon Monoxide in the Mechanism of Experimental Gastric Ulcers Healing, Regulation of Gastric Blood Flow and Accompanying Inflammation. Biochem. Pharmacol. 2018, 149, 131–142. [Google Scholar] [CrossRef]
- Andreu, V.; Mendoza, G.; Arruebo, M.; Irusta, S. Smart Dressings Based on Nanostructured Fibers Containing Natural Origin Antimicrobial, Anti-Inflammatory, and Regenerative Compounds. Materials 2015, 8, 5154–5193. [Google Scholar] [CrossRef] [PubMed]
- Cacciotti, I.; Ciocci, M.; di Giovanni, E.; Nanni, F.; Melino, S. Hydrogen Sulfide-Releasing Fibrous Membranes: Potential Patches for Stimulating Human Stem Cells Proliferation and Viability under Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 2368. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Zhao, Y.; Xian, M.; Wang, Q. Biological Thiols-Triggered Hydrogen Sulfide Releasing Microfibers for Tissue Engineering Applications. Acta Biomater. 2015, 27, 205–213. [Google Scholar] [CrossRef]
- Mauretti, A.; Neri, A.; Kossover, O.; Seliktar, D.; Nardo, P.D.; Melino, S. Design of a Novel Composite H2S-Releasing Hydrogel for Cardiac Tissue Repair. Macromol. Biosci. 2016, 16, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Abdelmonem, M.; Shahin, N.N.; Rashed, L.A.; Amin, H.A.A.; Shamaa, A.A.; Shaheen, A.A. Hydrogen Sulfide Enhances the Effectiveness of Mesenchymal Stem Cell Therapy in Rats with Heart Failure: In Vitro Preconditioning Versus In Vivo Co-Delivery. Biomed. Pharmacother. 2019, 112, 108584. [Google Scholar] [CrossRef]
- Liang, W.; Chen, J.; Li, L.; Li, M.; Wei, X.; Tan, B.; Shang, Y.; Fan, G.; Wang, W.; Liu, W. Conductive Hydrogen Sulfide-Releasing Hydrogel Encapsulating Adscs for Myocardial Infarction Treatment. ACS Appl. Mater. Interfaces 2019, 11, 14619–14629. [Google Scholar] [CrossRef]
- Carter, J.M.; Qian, Y.; Foster, J.C.; Matson, J.B. Peptide-Based Hydrogen Sulphide-Releasing Gels. Chem. Commun. 2015, 51, 13131–13134. [Google Scholar] [CrossRef]
- Longchamp, A.; Kaur, K.; Macabrey, D.; Dubuis, C.; Corpataux, J.S.; Deglise, M.; Matson, J.B.; Allagnat, F. Hydrogen Sulfide-Releasing Peptide Hydrogel Limits the Development of Intimal Hyperplasia in Human Vein Segments. Acta Biomater. 2019, 97, 374–384. [Google Scholar] [CrossRef]
- Shitole, A.A.; Raut, P.W.; Sharma, N.; Giram, P.; Khandwekar, A.P.; Garnaik, B. Electrospun Polycaprolactone/Hydroxyapatite/Zno Nanofibers as Potential Biomaterials for Bone Tissue Regeneration. J. Mater. Sci. Mater. Med. 2019, 30, 51. [Google Scholar] [CrossRef]
- Wu, Y.A.; Chiu, Y.C.; Lin, Y.H.; Ho, C.C.; Shie, M.Y.; Chen, Y.W. 3d-Printed Bioactive Calcium Silicate/Poly-Epsilon-Caprolactone Bioscaffolds Modified with Biomimetic Extracellular Matrices for Bone Regeneration. Int. J. Mol. Sci. 2019, 20, 942. [Google Scholar] [CrossRef]
- Fairag, R.; Rosenzweig, D.H.; Ramirez-Garcialuna, J.L.; Weber, M.H.; Haglund, L. Three-Dimensional Printed Polylactic Acid Scaffolds Promote Bone-Like Matrix Deposition In Vitro. ACS Appl. Mater. Interfaces 2019, 11, 15306–15315. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; You, M.L.; Ding, Z.Q.; Ye, W.B. A Review of Emerging Bone Tissue Engineering Via Peg Conjugated Biodegradable Amphiphilic Copolymers. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 97, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; An, G.; Liang, M.; Liu, Y.; Zhang, B.; Wang, Y. A Composited Peg-Silk Hydrogel Combining with Polymeric Particles Delivering Rhbmp-2 for Bone Regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 65, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Li, Z.; Organ, C.L.; Park, C.M.; Yang, C.T.; Pacheco, A.; Wang, D.; Lefer, D.J.; Xian, M. Ph-Controlled Hydrogen Sulfide Release for Myocardial Ischemia-Reperfusion Injury. J. Am. Chem. Soc. 2016, 138, 6336–6339. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.T.; Lai, Z.Z.; Zheng, Z.H.; Kang, J.M.; Xian, M.; Wang, R.Y.; Shi, K.; Meng, F.H.; Li, X.; Chen, L.; et al. A Novel Ph-Controlled Hydrogen Sulfide Donor Protects Gastric Mucosa from Aspirin-Induced Injury. J. Cell Mol. Med. 2017, 21, 2441–2451. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhushan, S.; Yang, C.; Otsuka, H.; Stein, J.D.; Pacheco, A.; Peng, B.; Devarie-Baez, N.O.; Aguilar, H.C.; Lefer, D.J.; et al. Controllable Hydrogen Sulfide Donors and Their Activity against Myocardial Ischemia-Reperfusion Injury. ACS Chem. Biol. 2013, 8, 1283–1290. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, C.; Organ, C.; Li, Z.; Bhushan, S.; Otsuka, H.; Pacheco, A.; Kang, J.; Aguilar, H.C.; Lefer, D.J.; et al. Design, Synthesis, and Cardioprotective Effects of N-Mercapto-Based Hydrogen Sulfide Donors. J. Med. Chem. 2015, 58, 7501–7511. [Google Scholar] [CrossRef]
- Zheng, Y.; Yu, B.; de la Cruz, L.K.; Choudhury, M.R.; Anifowose, A.; Wang, B. Toward Hydrogen Sulfide Based Therapeutics: Critical Drug Delivery and Developability Issues. Med. Res. Rev. 2018, 38, 57–100. [Google Scholar] [CrossRef]
- Zhao, Y.; Biggs, T.D.; Xian, M. Hydrogen Sulfide (H2S) Releasing Agents: Chemistry and Biological Applications. Chem. Commun. 2014, 50, 11788–11805. [Google Scholar] [CrossRef]
- Rapposelli, S.; Gambari, L.; Digiacomo, M.; Citi, V.; Lisignoli, G.; Manferdini, C.; Calderone, V.; Grassi, F. A Novel H2S-Releasing Amino-Bisphosphonate Which Combines Bone Anti-Catabolic and Anabolic Functions. Sci. Rep. 2017, 7, 11940. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Authors | H2S-Donor | In Vitro/In Vivo | Species | Main Findings |
|---|---|---|---|---|---|
| [14] | Grassi et al. |
| In vivo In vitro | Mice |
|
| [15] | Liu et al. |
| In vivo | Mice |
|
| [16] | Behera et al. |
| In vivo | Mice |
|
| [75] | Gambari et al. | GYY4137 loaded in a SF scaffold | In vitro | Human |
|
| [81] | Zhai et al. | NaHS | In vitro | Murine cell line |
|
| [82] | Lv et al. | GYY4137 | In vitro | Murine cell line |
|
| [83] | Mo et al. | GYY4137 | In vivo | Mice |
|
| [86] | Lee et al. | NaHS | In vitro | Human Mice |
|
| [88] | Jiang et al. | NaHS | In vitro | Human |
|
| [89] | Pu et al. | NaHS | In vivo | Mice |
|
| [90] | Xu et al. | GYY4137 | In vivo | Rat |
|
| [91] | Behera et al. | NaHS | In vivo In vitro | Mice |
|
| [93] | Yang et al. | NaHS | In vivo | Rats |
|
| [94] | Jiang et al. | GYY4137 | In vivo | Rabbits |
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambari, L.; Grigolo, B.; Grassi, F. Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. Int. J. Mol. Sci. 2019, 20, 5231. https://doi.org/10.3390/ijms20205231
Gambari L, Grigolo B, Grassi F. Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. International Journal of Molecular Sciences. 2019; 20(20):5231. https://doi.org/10.3390/ijms20205231
Chicago/Turabian StyleGambari, Laura, Brunella Grigolo, and Francesco Grassi. 2019. "Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives" International Journal of Molecular Sciences 20, no. 20: 5231. https://doi.org/10.3390/ijms20205231
APA StyleGambari, L., Grigolo, B., & Grassi, F. (2019). Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. International Journal of Molecular Sciences, 20(20), 5231. https://doi.org/10.3390/ijms20205231