The Role of Dimethyl Sulfoxide (DMSO) in Gene Expression Modulation and Glycosaminoglycan Metabolism in Lysosomal Storage Disorders on an Example of Mucopolysaccharidosis

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Cytotoxicity of DMSO in In Vitro Cell Cultures
2.2. Fibroblast Gene Expression Profiling upon DMSO Treatment
2.3. Gene Ontology (GO) Analysis of DMSO-Modulated Transcripts
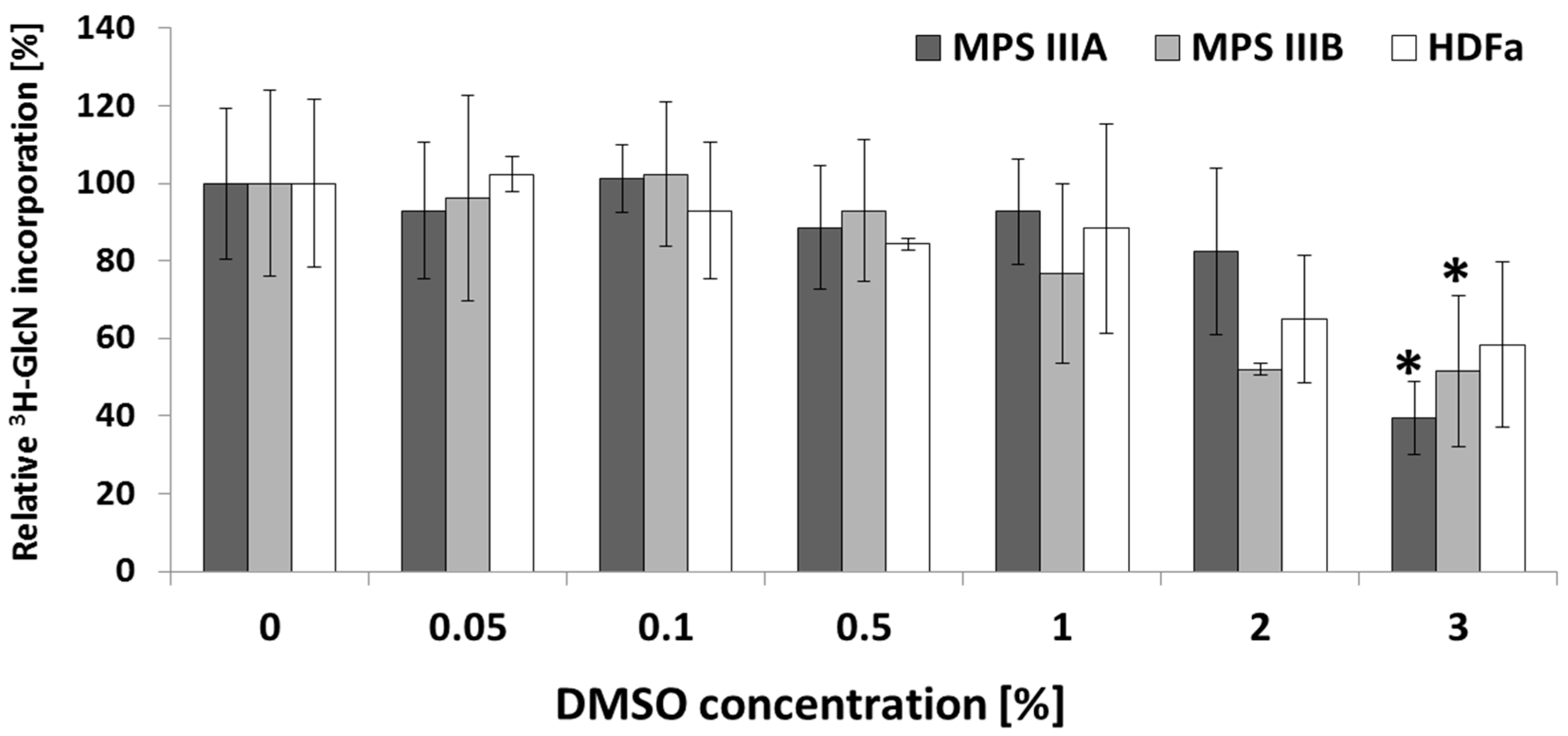
2.4. Kinetics of GAG Synthesis upon DMSO Treatment in MPS and HDFa Fibroblasts
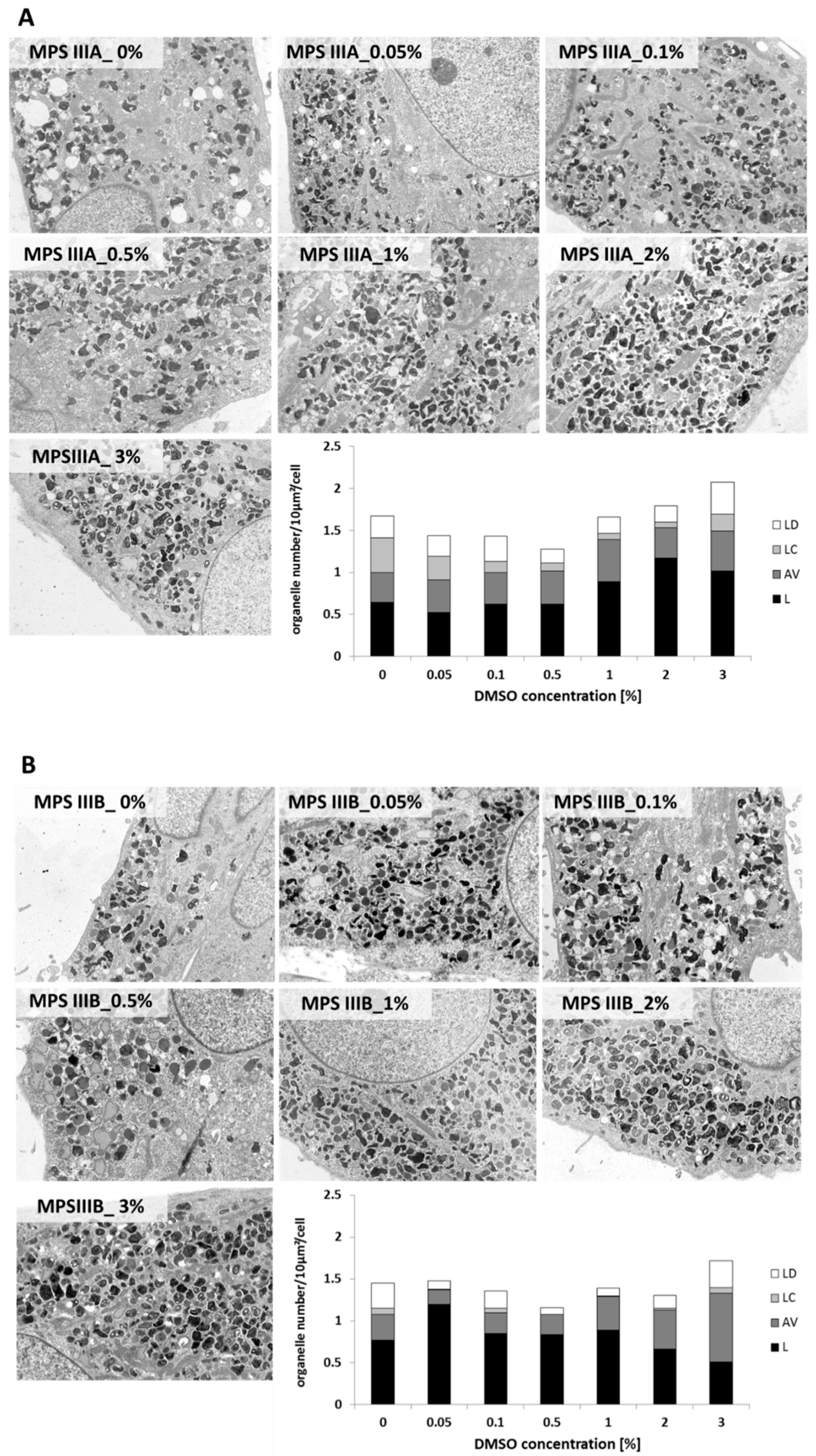
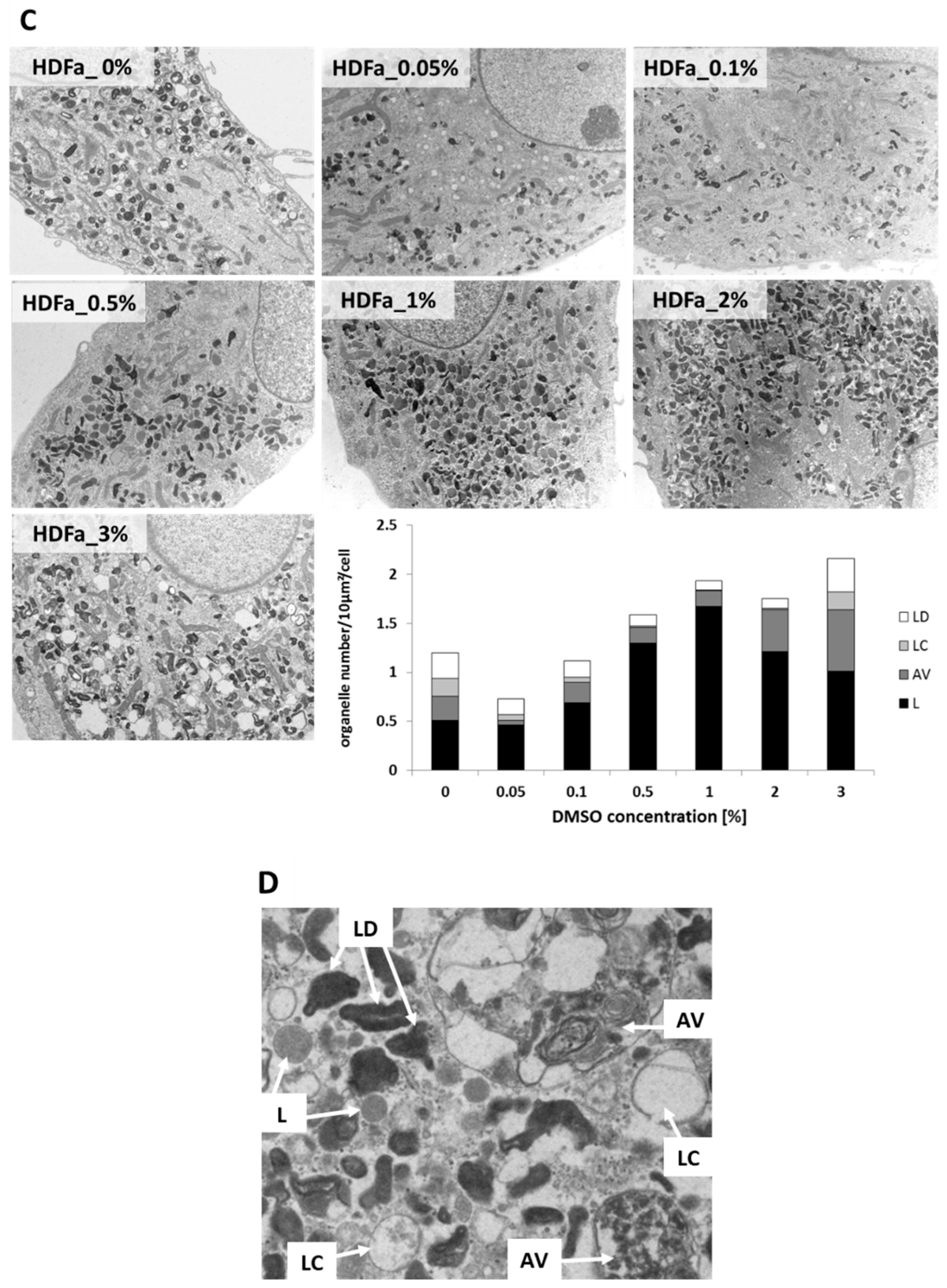
2.5. Electron Microscopy Studies of Lysosomal Compartment
3. Discussion
3.1. Cytotoxic Effect of DMSO on Human Fibroblasts at Higher Concentrations
3.2. Potential Role of DMSO in Regulation of Biological Processes on Transcriptional Level
3.3. Moderate DMSO Impact on GAG Synthesis
3.4. Alterations of Lysosomal Ultrastructure upon DMSO Treatment
4. Materials and Methods
4.1. Cell Lines, Culture Media, Supplements and Flavonoid Solutions
4.2. Cytotoxicity and Proliferation Assay
4.3. RNA Extraction
4.4. Microarray Assays for mRNA Analysis
4.5. Real-Time qRT-PCR Assays for mRNA Analysis
4.6. Measurement of Kinetics of GAG Synthesis
4.7. Transmission Electron Microscopy (TEM)
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ballabio, A.; Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochim. Biophys. Acta 2009, 1793, 684–696. [Google Scholar] [CrossRef]
- Gieselmann, V.; Braulke, T. Lysosomes. Biochimica et Biophysica Acta 2009, 1793, 603–604. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, E.U.; Muenzer, J. The mucopolysaccharidoses. In the Metabolic and Molecular Bases of Inherited Disease; Scriver CR 2001; McGraw-Hill: New York, NY, USA, 2001. [Google Scholar]
- Ohmi, K.; Kudo, L.C.; Ryazantsev, S.; Zhao, H.Z.; Karsten, S.L.; Neufeld, E.F. Sanfilippo syndrome type B, a lysosomal storage disease, is also a tauopathy. Proc. Natl. Acad. Sci. USA 2009, 106, 8332–8337. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Hulkova, H.; Dridi, L.; Dormoy-Raclet, V.; Grigoryeva, L.; Choi, Y.; Langford-Smith, A.; Wilkinson, F.L.; Ohmi, K.; DiCristo, G.; et al. Neuroinflammation, mitochondrial defects and neurodegeneration in mucopolysaccharidosis III type C mouse model. Brain 2015, 138, 336–355. [Google Scholar] [CrossRef] [PubMed]
- Beck, M. Emerging drugs for lysosomal storage diseases. Expert Opin. Emerg. Drugs 2010, 15, 495–507. [Google Scholar] [CrossRef]
- Piotrowska, E.; Jakobkiewicz-Banecka, J.; Baranska, S.; Tylki-Szymanska, A.; Czartoryska, B.; Wegrzyn, A.; Wegrzyn, G. Genistein-mediated inhibition of glycosaminoglycan synthesis as a basis for gene expression-targeted isoflavone therapy for mucopolysaccharidoses. Eur. J. Hum. Genet. 2006, 14, 846–852. [Google Scholar] [CrossRef]
- Malinowska, M.; Wilkinson, F.L.; Langford-Smith, K.J.; Langford-Smith, A.; Brown, J.R.; Crawford, B.E.; Vanier, M.T.; Grynkiewicz, G.; Wynn, R.F.; Wraith, J.E.; et al. Genistein Improves Neuropathology and Corrects Behaviour in a Mouse Model of Neurodegenerative Metabolic Disease. PLoS ONE 2010, 5, e14192. [Google Scholar] [CrossRef] [PubMed]
- Parenti, G.; Pignata, C.; Vajro, P.; Salerno, M. New strategies for the treatment of lysosomal storage diseases (review). Int. J. Mol. Med. 2013, 31, 11–20. [Google Scholar] [CrossRef]
- Giuffrida, G.; Lombardo, R.; Di Francesco, E.; Parrinello, L.; Di Raimondo, F.; Fiumara, A. Successful switch from enzyme replacement therapy to miglustat in an adult patient with type 1 Gaucher disease: A case report. J. Med. Case Rep. 2016, 10, 315. [Google Scholar] [CrossRef]
- Moskot, M.; Gabig-Ciminska, M.; Jakobkiewicz-Banecka, J.; Wesierska, M.; Bochenska, K.; Wegrzyn, G. Cell cycle is disturbed in mucopolysaccharidosis type II fibroblasts, and can be improved by genistein. Gene 2016, 585, 100–103. [Google Scholar] [CrossRef]
- Moskot, M.; Jakobkiewicz-Banecka, J.; Smolinska, E.; Banecki, B.; Wegrzyn, G.; Gabig-Ciminska, M. Activities of genes controlling sphingolipid metabolism in human fibroblasts treated with flavonoids. Metab. Brain Dis. 2015, 30, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Moskot, M.; Montefusco, S.; Jakobkiewicz-Banecka, J.; Mozolewski, P.; Wegrzyn, A.; Di Bernardo, D.; Wegrzyn, G.; Medina, D.L.; Ballabio, A.; Gabig-Ciminska, M. The Phytoestrogen Genistein Modulates Lysosomal Metabolism and Transcription Factor EB (TFEB) Activation. J. Biol. Chem. 2014, 289, 17054–17069. [Google Scholar] [CrossRef] [PubMed]
- Moskot, M.; Jakobkiewicz-Banecka, J.; Kloska, A.; Smolinska, E.; Mozolewski, P.; Malinowska, M.; Rychlowski, M.; Banecki, B.; Wegrzyn, G.; Gabig-Ciminska, M. Modulation of expression of genes involved in glycosaminoglycan metabolism and lysosome biogenesis by flavonoids. Sci. Rep. 2015, 5, 9378. [Google Scholar] [CrossRef] [PubMed]
- Brobyn, R.D. The human toxicology of dimethyl sulfoxide. Ann. N. Y. Acad. Sci. 1975, 243, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Tipping, P.G. Attenuation of adjuvant arthritis in rats by treatment with oxygen radical scavengers. Immunol. Cell Biol. 1994, 72, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Rosenstein, E.D. Topical agents in the treatment of rheumatic disorders. Rheum. Dis. Clin. N. Am. 1999, 25, 899–918. [Google Scholar] [CrossRef]
- Santos, N.C.; Figueira-Coelho, J.; Martins-Silva, J.; Saldanha, C. Multidisciplinary utilization of dimethyl sulfoxide: Pharmacological, cellular, and molecular aspects. Biochem. Pharmacol. 2003, 65, 1035–1041. [Google Scholar] [CrossRef]
- Broadwell, R.D.; Salcman, M.; Kaplan, R.S. Morphologic effects of dimethyl sulfoxide on the blood-brain barrier. Science 1982, 217, 164–166. [Google Scholar] [CrossRef]
- Ikeda, Y.; French, K.B.; Ikeda, K.; Long, D.M. Comparative effects of direct and indirect hydroxyl radical scavengers on traumatic brain edema. J. Neurosurg. 1990, 72, A360. [Google Scholar]
- Luikart, S.D.; Maniglia, C.A.; Sartorelli, A.C. Glycosaminoglycan synthesis during differentiation of HL60/HGPRT-leukemia cells induced by dimethyl sulfoxide and 12-O-tetradecanoylphorbol-13-acetate. Cancer Res. 1984, 44, 2907–2912. [Google Scholar]
- Kaneski, C.R.; Constantopoulos, G.; Brady, R.O. Effect of dimethylsulfoxide on the proliferation and glycosaminoglycan synthesis of rat prostate adenocarcinoma cells (PAIII) in vitro: Isolation and characterization of DMSO-resistant cells. Prostate 1991, 18, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Soler, R.; Bruschini, H.; Truzzi, J.C.; Martins, J.R.; Camara, N.O.; Alves, M.T.; Leite, K.R.; Nader, H.B.; Srougi, M.; Ortiz, V. Urinary glycosaminoglycans excretion and the effect of dimethyl sulfoxide in an experimental model of non-bacterial cystitis. Int. Braz j Urol 2008, 34, 503–511. [Google Scholar] [CrossRef]
- Dairkee, S.H.; Glaser, D.A. Dimethyl sulfoxide affects colony morphology on agar and alters distribution of glycosaminoglycans and fibronectin. Proc. Natl. Acad. Sci. USA 1982, 79, 6927–6931. [Google Scholar] [CrossRef] [PubMed]
- Constantopoulos, G.; Hawkins, C.S.; Kornblith, P.L. Dimethylsulfoxide inhibits glycosaminoglycan synthesis and cell growth in human gliomas in vitro. Trans. Am. Soc. Neurochem. 1986, 17, 223. [Google Scholar]
- Chiang, T.S.; Yang, K.C.; Chiou, L.L.; Huang, G.T.; Lee, H.S. Enhancement of CYP3A4 Activity in Hep G2 Cells by Lentiviral Transfection of Hepatocyte Nuclear Factor-1 Alpha. PLoS ONE 2014, 9, e94885. [Google Scholar] [CrossRef] [PubMed]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S. AmiGO Hub, Web Presence Working Group. AmiGO: Online access to ontology and annotation data. Bioinformatics 2009, 25, 288–289. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Kloska, A.; Narajczyk, M.; Jakobkiewicz-Banecka, J.; Grynkiewicz, G.; Szeja, W.; Gabig-Ciminska, M.; Wegrzyn, G. Synthetic genistein derivatives as modulators of glycosaminoglycan storage. J. Transl. Med. 2012, 10, 153. [Google Scholar] [CrossRef]
- Siller, R.; Greenhough, S.; Naumovska, E.; Sullivan, G.J. Small-molecule-driven hepatocyte differentiation of human pluripotent stem cells. Stem Cell Rep. 2015, 4, 939–952. [Google Scholar] [CrossRef]
- Rodriguez-Burford, C.; Oelschlager, D.K.; Talley, L.I.; Barnes, M.N.; Partridge, E.E.; Grizzle, W.E. The use of dimethylsulfoxide as a vehicle in cell culture experiments using ovarian carcinoma cell lines. Biotech. Histochem. 2003, 78, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Sakuragawa, N.; Sato, M.; Yoshida, Y.; Kamo, I.; Arima, M.; Satoyoshi, E. Effects of dimethylsulfoxide on sphingomyelinase in cultured human fibroblasts and correction of sphingomyelinase deficiency in fibroblasts from Niemann-Pick patients. Biochem. Biophys. Res. Commun. 1985, 126, 756–762. [Google Scholar] [CrossRef]
- Malinowska, M.; Wilkinson, F.L.; Bennett, W.; Langford-Smith, K.J.; O’Leary, H.A.; Jakobkiewicz-Banecka, J.; Wynn, R.; Wraith, J.E.; Wegrzyn, G.; Bigger, B.W. Genistein reduces lysosomal storage in peripheral tissues of mucopolysaccharide IIIB mice. Mol. Genet. Metab. 2009, 98, 235–242. [Google Scholar] [CrossRef]
- Morley, P.; Whitfield, J.F. The differentiation inducer, dimethyl sulfoxide, transiently increases the intracellular calcium ion concentration in various cell types. J. Cell. Physiol. 1993, 156, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Yoshida, Y.; Sakuragawa, N.; Arima, M. Effects of dimethylsulfoxide on sphingomyelinase activities in normal and Niemann-Pick type A, B and C fibroblasts. Biochim. Biophys. Acta 1988, 962, 59–65. [Google Scholar]
- Jakóbkiewicz-Banecka, J.; Piotrowska, E.; Narajczyk, M.; Barańska, S.; Wegrzyn, G. Genistein-mediated inhibition of glycosaminoglycan synthesis, which corrects storage in cells of patients suffering from mucopolysaccharidoses, acts by influencing an epidermal growth factor-dependent pathway. J. Biomed. Sci. 2009, 16, 26. [Google Scholar] [CrossRef]
- Kloska, A.; Jakóbkiewicz-Banecka, J.; Narajczyk, M.; Banecka-Majkutewicz, Z.; Węgrzyn, G. Effects of flavonoids on glycosaminoglycan synthesis: Implications for substrate reduction therapy in Sanfilippo disease and other mucopolysaccharidoses. Metab. Brain Dis. 2011, 26, 1–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of DEGs in Term | ||||||
|---|---|---|---|---|---|---|
| GO Term | 1 h | 24 h | 48 h | |||
| Molecular Function | [5] | [7] | [2] | [7] | [6] | [8] |
| binding (GO:0005488) | 10 (29.4%) | 73 (31.1%) | 4 (19.0%) | 35 (22.0%) | 27 (26.5%) | 37 (27.0%) |
| catalytic activity (GO:0003824) | 8 (23.5%) | 53 (22.6%) | 3 (14.3%) | 44 (27.7%) | 23 (22.5%) | 35 (25.5%) |
| transporter activity (GO:0005215) | 1 (2.9%) | 9 (3.8%) | 4 (2.5%) | 8 (7.8%) | 5 (3.6%) | |
| signal transducer activity (GO:0004871) | 1 (2.9%) | 8 (3.4%) | 2 (1.3%) | 3 (2.9%) | 4 (2.9%) | |
| receptor activity (GO:0004872) | 1 (2.9%) | 12 (5.1%) | 7 (4.4%) | 2 (2.0%) | 2 (1.5% | |
| structural molecule activity (GO:0005198) | 1 (2.9%) | 7 (3.0%) | 3 (1.9%) | 3 (2.2%) | ||
| translation regulator activity (GO:0045182) | 3 (1.3%) | 2 (1.3%) | 3 (2.2%) | |||
| Biological Process | [9] | [13] | [9] | [12] | [11] | [11] |
| metabolic process (GO:0008152) | 15 (44.1%) | 79 (33.6%) | 6 (38.1%) | 54 (34.0%) | 33 (32.4%) | 52 (38.0%) |
| cellular process (GO:0009987) | 11 (32.4%) | 113 (48.1%) | 8 (28.6%) | 70 (44.0%) | 45 (44.1%) | 52 (38.0%) |
| cellular component organization or biogenesis (GO:0071840) | 2 (5.9%) | 27 (11.5%) | 2 (9.5%) | 20 (12.6%) | 5 (4.9%) | 16 (11.7%) |
| biological regulation (GO:0065007) | 3 (8.8%) | 38 (16.2%) | 4 (19.0%) | 19 (11.9%) | 10 (9.8%) | 17 (12.4%) |
| response to stimulus (GO:0050896) | 3 (8.8%) | 32 (13.6%) | 2 (9.5%) | 16 (10.1%) | 9 (8.8%) | 15 (10.9%) |
| developmental process (GO:0032502) | 27 (11.5%) | 2 (9.5%) | 16 (10.1%) | 11 (10.8%) | 8 (5.8%) | |
| localization (GO:0051179) | 2 (5.9%) | 26 (11.1%) | 10 (6.3%) | 9 (8.8%) | 13 (9.5%) | |
| multicellular organismal process (GO:0032501) | 19 (8.1%) | 10 (6.3%) | 4 (2.9%) | |||
| Cellular Component | [4] | [7] | [3] | [7] | [5] | [7] |
| cell part (GO:0044464) | 8 (23.5%) | 84 (35.7%) | 4 (19.0%) | 44 (27.7%) | 29 (28.4%) | 49 (35.8%) |
| organelle (GO:0043226) | 6 (17.6%) | 63 (26.8%) | 4 (19.0%) | 28 (17.6%) | 21 (20.6%) | 35 (25.5%) |
| membrane (GO:0016020) | 1 (2.9%) | 27 (11.5%) | 11 (6.9%) | 10 (9.8%) | 16 (11.7%) | |
| macromolecular complex (GO:0032991) | 1 (2.9%) | 30 (12.8%) | 3 (14.3%) | 12 (7.5%) | 5 (4.9%) | 19 (13.9%) |
| extracellular region (GO:0005576) | 7 (3.0%) | 7 (4.4%) | 3 (2.9%) | 3 (2.2%) | ||
| Protein Class | [5] | [22] | [5] | [16] | [19] | [18] |
| transcription factor (PC00218) | 5 (14.7%) | 24 (10.2%) | 2 (9.5%) | 8 (5.0%) | 8 (7.8%) | 7 (5.1%) |
| nucleic acid binding (PC00171) | 4 (11.8%) | 30 (12.8%) | 2 (9.5%) | 14 (8.8%) | 7 (6.9%) | 19 (13.9%) |
| hydrolase (PC00121) | 3 (8.8%) | 17 (7.2%) | 1 (4.8%) | 8 (5.0%) | 5 (4.9%) | 11 (8.0%) |
| transferase (PC00220) | 2 (5.9%) | 19 (8.1%) | 1 (4.8%) | 10 (6.3%) | 5 (4.9%) | 10 (7.3%) |
| enzyme modulator (PC00095) | 10 (4.3%) | 6 (3.8%) | 7 (5.1%) | |||
| receptor (PC00197) | 9 (3.8%) | 1 (4.8%) | 7 (4.4%) | 3 (2.9%) | ||
| transporter (PC00227) | 7 (3.0%) | 5 (3.1%) | 7 (6.9%) | 5 (3.6%) | ||
| signaling molecule (PC00207) | 9 (3.8%) | 7 (4.4%) | 6 (5.9%) | 7 (5.1%) | ||
| transfer/carrier protein (PC00219) | 1 (2.9%) | |||||
| (A) | |||||
| Time of Exposure to DMSO | |||||
| 1 h | 24 h | 48 h | |||
| Gene Expression Modulation | |||||
| BBS12 | 0.58 ± 0.07 | ACVRL1 | 0.60 ± 0.11 | ALG2 | 0.60 ± 0.08 |
| CDKN1C | 0.60 ± 0.11 | APEG1 | 0.63 ± 0.15 | ANKRD26 | 0.64 ± 0.14 |
| DDIT4 | 0.51 ± 0.10 | ATF2 | 0.66 ± 0.02 | ANKRD38 | 0.51 ± 0.15 |
| EBF1 | 0.67 ± 0.05 | CCDC15 | 0.68 ± 0.12 | ATP7B | 0.62 ± 0.17 |
| FLJ10246 | 0.56 ± 0.08 | CHODL | 0.60 ± 0.14 | BAIAP2L1 | 0.66 ± 0.12 |
| FLJ45244 | 0.63 ± 0.11 | EDNRA | 0.61 ± 0.06 | BEX2 | 0.59 ± 0.09 |
| GM2A | 0.61 ± 0.07 | FAM21A | 0.65 ± 0.12 | DAK | 0.59 ± 0.10 |
| HCFC2 | 0.63 ± 0.06 | GGT7 | 0.68 ± 0.05 | DHRSX | 0.61 ± 0.15 |
| HSPA1L | 0.57 ± 0.08 | LMNB1 | 0.60 ± 0.04 | DKFZp761P0423 | 0.61 ± 0.04 |
| KCNJ2 | 0.62 ± 0.06 | MCPH1 | 0.67 ± 0.03 | DNAJC1 | 0.57 ± 0.08 |
| KIAA0100 | 0.63 ± 0.18 | MFHAS1 | 0.66 ± 0.00 | DNAJC3 | 0.65 ± 0.02 |
| KRCC1 | 0.66 ± 0.09 | PTHLH | 0.68 ± 0.07 | EIF2C4 | 0.55 ± 0.04 |
| LIAS | 0.64 ± 0.02 | RNF141 | 0.69 ± 0.02 | FAM127C | 0.66 ± 0.19 |
| LZTFL1 | 0.67 ± 0.01 | SNORD16 | 0.55 ± 0.02 | FAM83H | 0.52 ± 0.06 |
| MGA | 0.62 ± 0.11 | SSX2IP | 0.69 ± 0.00 | FKBPL | 0.59 ± 0.12 |
| NICN1 | 0.63 ± 0.09 | SURF2 | 0.67 ± 0.08 | FLJ11151 | 0.62 ± 0.10 |
| PLAC9 | 0.65 ± 0.02 | TAF13 | 0.68 ± 0.13 | FLJ23834 | 0.66 ± 0.04 |
| SLC40A1 | 0.66 ± 0.02 | WIPF2 | 0.65 ± 0.01 | FLJ35220 | 0.64 ± 0.10 |
| SPIN1 | 0.67 ± 0.03 | ZKSCAN3 | 0.67 ± 0.01 | FLJ90086 | 0.60 ± 0.02 |
| TAOK1 | 0.67 ± 0.10 | ZNF417 | 0.67 ± 0.10 | FRY | 0.61 ± 0.09 |
| (B) | |||||
| Time of Exposure to DMSO | |||||
| 1 h | 24 h | 48 h | |||
| Gene Expression Modulation | |||||
| ALG2 | 1.64 ± 0.27 | BMP6 | 1.65 ± 0.20 | ADAM9 | 1.62 ± 0.33 |
| ANKS3 | 1.62 ± 0.02 | FAM113B | 1.61 ± 0.21 | APCDD1L | 1.59 ± 0.47 |
| BET1L | 1.60 ± 0.23 | GFOD1 | 1.73 ± 0.29 | BHLHB2 | 1.56 ± 0.21 |
| CDC42EP3 | 2.04 ± 0.04 | GSTM5 | 1.55 ± 0.25 | CXCL1 | 1.70 ± 0.03 |
| CDK8 | 1.59 ± 0.19 | ITGBL1 | 1.79 ± 0.53 | ERCC4 | 1.54 ± 0.41 |
| DENND2C | 1.60 ± 0.15 | KCNJ16 | 1.55 ± 0.35 | ETV3 | 1.73 ± 0.41 |
| F5 | 1.59 ± 0.01 | KIAA0746 | 1.80 ± 0.08 | GFM2 | 1.69 ± 0.06 |
| FLJ30058 | 1.60 ± 0.27 | LENG8 | 1.73 ± 0.19 | GNAQ | 1.59 ± 0.26 |
| GALNAC4S-6ST | 1.69 ± 0.45 | MEGF8 | 1.72 ± 0.51 | HES4 | 1.58 ± 0.33 |
| GRK6 | 1.63 ± 0.37 | MESP1 | 1.64 ± 0.38 | HPS3 | 1.54 ± 0.14 |
| KLF10 | 1.90 ± 0.11 | MT1G | 1.62 ± 0.27 | KLF10 | 1.66 ± 0.32 |
| LMAN2 | 1.70 ± 0.19 | MTE | 1.55 ± 0.46 | MAGEL2 | 1.65 ± 0.45 |
| MAP6D1 | 1.59 ± 0.45 | NAP1L3 | 1.57 ± 0.32 | OXCT1 | 1.65 ± 0.23 |
| MYBBP1A | 1.66 ± 0.48 | NDNL2 | 1.66 ± 0.47 | PMS2 | 1.80 ± 0.34 |
| NR4A1 | 1.74 ± 0.25 | NEFM | 1.53 ± 0.37 | SNORD31 | 1.78 ± 0.41 |
| SPIN2B | 1.62 ± 0.46 | SLC38A4 | 1.69 ± 0.07 | SNX12 | 1.73 ± 0.17 |
| TJP2 | 1.84 ± 0.36 | STAC | 1.61 ± 0.13 | SPG21 | 1.57 ± 0.45 |
| UBA2 | 1.65 ± 0.20 | STK11 | 1.55 ± 0.05 | STIM2 | 1.58 ± 0.19 |
| UBA3 | 1.60 ± 0.27 | ZBBX | 1.55 ± 0.02 | TBC1D23 | 1.55 ± 0.43 |
| ZNF597 | 1.62 ± 0.22 | ZSCAN16 | 1.54 ± 0.21 | TMEM161A | 1.76 ± 0.24 |
| Gene Expression | Term | Genes | 1 h | 24 h | 48 h | |||
|---|---|---|---|---|---|---|---|---|
| FC ± SD | FC ± SD | FC ± SD | ||||||
| Microarray | Real-Time qRT-PCR | Microarray | Real-Time qRT-PCR | Microarray | Real-Time qRT-PCR | |||
| Upregulation | Glycan biosynthesis and metabolism | B4GALT1 | 0.83 ± 0.0 | 0.81 ± 0.0 | 1.32 ± 0.1 | 1.34 ± 0.3 | 0.94 ± 0.0 | |
| GALNT2 | 1.46 ± 0.2 | 1.38 ± 0.1 | 1.08 ± 0.1 | 1.12 ± 0.1 | 1.01 ± 0.4 | |||
| GALNT4 | 1.50 ± 0.2 | 1.39 ± 0.1 | 0.87 ± 0.1 | 0.93 ± 0.1 | 1.34 ± 0.2 | 1.05 ± 0.0 | ||
| GALNT6 | 1.11 ± 0.1 | 1.17 ± 0.1 | 0.91 ± 0.0 | 0.91 ± 0.0 | 1.31 ± 0.1 | 1.27 ± 0.1 | ||
| GALNT12 | 1.33 ± 0.1 | 1.29 ± 0.0 | 0.83 ± 0.0 | 0.91 ± 0.1 | 1.26 ± 0.0 | |||
| MAN2C1 | 1.37 ± 0.0 | 1.42 ± 0.0 | 0.90 ± 0.1 | 1.01 ± 0.1 | 0.78 ± 0.1 | |||
| N-Glycan biosynthesis | ALG12 | 1.12 ± 0.4 | 1.30 ± 0.1 | 1.40 ± 0.0 | 1.08 ± 0.2 | 0.78 ± 0.2 | 1.19 ± 0.1 | |
| CS/DS biosynthesis | CHST15 | 1.7 ± 0.4 | 1.73 ± 0.1 | 1.14 ± 0.4 | 1.00 ± 0.2 | 0.98 ± 0.0 | ||
| HS synthesis | EXT1 | 1.46 ± 0.1 | 1.48 ± 0.0 | 0.94 ± 0.0 | 0.83 ± 0.1 | 0.88 ± 0.3 | 0.99 ± 0.0 | |
| KS biosynthesis | B4GALT1 | 0.83 ± 0.0 | 0.81 ± 0.0 | 1.32 ± 0.1 | 1.34 ± 0.0 | 0.94 ± 0.0 | ||
| CHST2 | 1.42 ± 0.1 | 1.15 ± 0.5 | ||||||
| GAG (CS, KS) degradation | GALNS | 1.03 ± 0.2 | 0.98 ± 0.1 | 1.37 ± 0.3 | 1.42 ± 0.2 | 1.25 ± 0.1 | 1.02 ± 0.0 | |
| SL metabolism | SGPL1 | 1.34 ± 0.0 | 1.31 ± 0.3 | 1.18 ± 0.3 | 0.88 ± 0.0 | |||
| SPTLC2 | 0.95 ± 0.0 | 0.91 ± 0.1 | 1.01 ± 0.0 | 1.48 ± 0.4 | 1.31 ± 0.1 | |||
| SPTLC3 | 1.17 ± 0.0 | 1.31 ± 0.3 | 1.41 ± 0.2 | 1.37 ± 0.1 | ||||
| GSLs biosynthesis | B4GALT1 | 0.83 ± 0.0 | 0.81 ± 0.0 | 1.32 ± 0.1 | 1.34 ± 0.0 | 0.94 ± 0.0 | ||
| ST3GAL5 | 1.32 ± 0.2 | 1.32 ± 0.0 | 1.09 ± 0.1 | 1.03 ± 0.1 | 0.89 ± 0.0 | 0.97 ± 0.0 | ||
| SLC7A5 | 1.32 ± 0.1 | 1.30 ± 0.0 | 1.21 ± 0.5 | 1.07 ± 0.0 | 1.06 ± 0.0 | |||
| Sphingosine biosynthesis | SPTLC2 | 0.95 ± 0.0 | 0.91 ± 0.1 | 1.02 ± 0.2 | 1.01 ± 0.0 | 1.48 ± 0.4 | 1.31 ± 0.1 | |
| SPTLC3 | 1.17 ± 0.0 | 1.31 ± 0.3 | 1.41 ± 0.2 | 1.37 ± 0.1 | ||||
| Lysosome | ARSK | 0.83 ± 0.2 | 1.11 ± 0.6 | 1.14 ± 0.0 | 1.37 ± 0.1 | 1.44 ± 0.5 | ||
| Downregulation | N-Glycan biosynthesis | ALG2 | 1.64 ± 0.3 | 1.61 ± 0.1 | 0.97 ± 0.2 | 0.99 ± 0.0 | 0.60 ± 0.1 | 0.92 ± 0.0 |
| Lysosome | GM2A | 0.61 ± 0.1 | 0.58 ± 0.0 | 0.98 ± 0.0 | 0.99 ± 0.0 | 1.60 ± 0.1 | ||
| MCOLN1 | 1.01 ± 0.2 | 0.99 ± 0.0 | 0.97 ± 0.0 | 0.99 ± 0.0 | 0.68 ± 0.2 | 0.81 ± 0.1 | ||
| Master repressor of autophagy | ZKSCAN3 | 0.51 ± 0.0 | 0.67 ± 0.0 | 0.92 ± 0.0 | 0.69 ± 0.2 | 0.88 ± 0.0 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moskot, M.; Jakóbkiewicz-Banecka, J.; Kloska, A.; Piotrowska, E.; Narajczyk, M.; Gabig-Cimińska, M. The Role of Dimethyl Sulfoxide (DMSO) in Gene Expression Modulation and Glycosaminoglycan Metabolism in Lysosomal Storage Disorders on an Example of Mucopolysaccharidosis. Int. J. Mol. Sci. 2019, 20, 304. https://doi.org/10.3390/ijms20020304
Moskot M, Jakóbkiewicz-Banecka J, Kloska A, Piotrowska E, Narajczyk M, Gabig-Cimińska M. The Role of Dimethyl Sulfoxide (DMSO) in Gene Expression Modulation and Glycosaminoglycan Metabolism in Lysosomal Storage Disorders on an Example of Mucopolysaccharidosis. International Journal of Molecular Sciences. 2019; 20(2):304. https://doi.org/10.3390/ijms20020304
Chicago/Turabian StyleMoskot, Marta, Joanna Jakóbkiewicz-Banecka, Anna Kloska, Ewa Piotrowska, Magdalena Narajczyk, and Magdalena Gabig-Cimińska. 2019. "The Role of Dimethyl Sulfoxide (DMSO) in Gene Expression Modulation and Glycosaminoglycan Metabolism in Lysosomal Storage Disorders on an Example of Mucopolysaccharidosis" International Journal of Molecular Sciences 20, no. 2: 304. https://doi.org/10.3390/ijms20020304
APA StyleMoskot, M., Jakóbkiewicz-Banecka, J., Kloska, A., Piotrowska, E., Narajczyk, M., & Gabig-Cimińska, M. (2019). The Role of Dimethyl Sulfoxide (DMSO) in Gene Expression Modulation and Glycosaminoglycan Metabolism in Lysosomal Storage Disorders on an Example of Mucopolysaccharidosis. International Journal of Molecular Sciences, 20(2), 304. https://doi.org/10.3390/ijms20020304