Overview of the Structure–Function Relationships of Mannose-Specific Lectins from Plants, Algae and Fungi




Abstract
1. Introduction
2. Diversity of Mannose-Binding Lectins in Higher Plants
3. Structural Organization of the Plant, Algal and Fungal Mannose-Binding Lectins
3.1. Structure of Mannose-Specific Plant Lectins
3.1.1. The β-Sandwich Fold
3.1.2. The β-Prism I Fold
3.1.3. The β-Prism II Fold
3.2. Structure of Mannose-Specific Algal Lectins
3.3. Structure of Mannose-Specific Fungal Lectins
4. The Mannosyl-Binding Specificities of Mannose-Binding Lectins
4.1. The Mannose-Binding Specificity
4.2. The Oligosaccharide-Binding Specificity
5. Functions of Mannose-Specific Lectins
5.1. Insecticidal Activity
5.2. Resistance to Abiotic (and Biotic) Stresses
6. Medical Applications for the Mannose-Specific Lectins
6.1. Mannose-Specific Lectins as Immunomodulators
6.2. Mannose-Specific Lectins as Cancer Biomarkers and Anti-Cancer Drugs
7. Biomedical Perspectives for Mannose-Specific Lectins
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | Agaricus bisporus agglutinin |
| ACA | Allium cepa agglutinin |
| AMA | Arum maculatum agglutinin |
| ASA | Allium sativum agglutinin |
| Asp, D | Aspartic acid |
| Ath | Arabidopsis thaliana |
| ATP | Adenosine triphosphate |
| BMA | Bowringia mildbraedii agglutinin |
| CBA | Carbohydrate-binding agent |
| CBM | Carbohydrate-binding module |
| CBS | Carbohydrate-binding site |
| CEA | Colocasia esculenta agglutinin |
| Con A | Concanavalin A |
| CTL | Centrolobium tomentosum lectin |
| CVN | Cyanovirin N |
| EUL | Euonymus europaeus lectin |
| Glu, E | Glutamic acid |
| Gly, G | Glycine |
| GNA | Galanthus nivalis agglutinin |
| Heltuba | Helianthus tuberosus agglutinin |
| HHA | Hippeastrum hybdrid agglutinin |
| HIV | Human Immunodeficiency virus |
| HSP | Heat shock protein |
| Leu, L | Leucine |
| LcA | Lens culinaris agglutinin |
| LoL | Lathyrus ochrus lectin |
| MPA | Maclura pomifera agglutinin |
| MVL | Microcystis viridis lectin |
| OAA | Oscillatoria agardhii agglutinin |
| PAL | Pterocarpus angolensis lectin |
| PDB | Protein data bank |
| PHA | Phytohemagglutinin |
| Phe, F | Phenylalanine |
| PNA | Peanut agglutinin |
| PsA | Pisum sativum agglutinin |
| ROS | Reactive oxygen species |
| RIP | Ribosome inactivating protein |
| SBA | Soybean agglutinin |
| SNA | Sambucus nigra agglutinin |
| Trp, W | Tryptophane |
| Tyr, Y | Tyrosine |
| Val, V | Valine |
| VfA | Vicia faba agglutinin |
References
- Van Damme, E.J.M.; Rougé, P.; Peumans, W.J. Plant lectins. In Carbohydrate-Protein Interactions: Plant Lectins; Kamerling, J.P., Boons, G.J., Lee, Y.C., Suzuki, A., Taniguchi, N., Voragen, A.G.I., Eds.; Elsevier: New York, NY, USA, 2007; pp. 564–599. [Google Scholar]
- Wu, A.M.; Song, S.C.; Tsai, M.S.; Herp, A. A guide to the carbohydrate specificities of applied lectins-2. Adv. Exp. Med. Biol. 2001, 491, 551–585. [Google Scholar]
- De Schutter, K.; Van Damme, E.J.M. Protein-carbohydrate interactions as part of plant defense and animal immunity. Molecules 2015, 20, 9029–9053. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Lannoo, N.; Fouquaert, E.; Peumans, W.J. The identification of inducible cytoplasmic/nuclear carbohydrate-binding proteins urges to develop novel concepts about the role of plant lectins. Glycoconj. J. 2004, 20, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peumans, W.J.; Hause, B.; Bras, J.; Kumar, M.; Proost, P.; Barre, A.; Rougé, P.; Van Damme, E.J.M. Jasmonic acid methyl ester induces the synthesis of a cytoplasmic/nuclear chito-oligosaccharide binding lectin in tobacco leaves. FASEB J. 2002, 16, 905–907. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Peumans, W.J.; Barre, A.; Rougé, P. Plant lectins: A composite of several distinct families of structurally and evolutionary related proteins with diverse biological roles. Crit. Rev. Plant Sci. 1998, 17, 575–692. [Google Scholar] [CrossRef]
- Agrawal, B.B.; Goldstein, I.J. Specific binding of concanavalin A to cross-linked dextran gels. Biochem. J. 1965, 96, 23C–25C. [Google Scholar] [CrossRef]
- Singh, R.S.; Bhari, R.; Kaur, H.P. Mushroom lectins: Current status and future perspectives. Crit. Rev. Biotechnol. 2010, 30, 99–126. [Google Scholar] [CrossRef]
- Okuyama, S.; Nakamura-Tsuruta, S.; Tateno, H.; Hirabayashi, J.; Matsubara, K.; Hori, K. Strict binding specificity of small-sized lectins from the red alga Hypnea japonica for core (α1-6) fucosylated N-glycans. Biosci. Biotechnol. Biochem. 2009, 73, 912–920. [Google Scholar] [CrossRef]
- Debray, H.; Rougé, P. The fine sugar specificity of the Lathyrus ochrus seed lectin and isolectins. FEBS Lett. 1984, 176, 120–124. [Google Scholar] [CrossRef]
- Tateno, H.; Nakamura-Tsuruta, S.; Hirabayashi, J. Comparative analysis of core-fucose-binding lectins from Lens culinaris and Pisum sativum using frontal affinity chromatography. Glycobiology 2009, 19, 527–536. [Google Scholar] [CrossRef]
- Tateno, H.; Winter, H.C.; Petryniak, J.; Goldstein, I.J. Purification, characterization, molecular cloning, and expression of novel members of jacalin-related lectins from rhizomes of the true fern Phlebodium aureum (L.) J. Smith (Polypodiaceae). J. Biol. Chem. 2003, 278, 10891–10899. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.K.; Figueroa, M.O.R.; Lajolo, F.M. Purification and characterization of two major lectins from Araucaria brasiliensis syn Araucaria angustifolia seeds (pinhao). Plant Physiol. 1991, 97, 856–862. [Google Scholar] [CrossRef]
- Miyakawa, T.; Hatano, K.; Miyauchi, Y.; Suwa, Y.; Sawano, Y.; Tanokura, M. A secreted protein with plant-specific cysteine-rich motif functions as a mannose-binding lectin that exhibits antifungal activity. Plant Physiol. 2014, 166, 766–778. [Google Scholar] [CrossRef] [PubMed]
- Yagi, F.; Iwaya, T.; Haraguchi, T.; Goldstein, I.J. The lectin from leaves of Japanese cycad, Cycas revoluta Thunb. (Gymnosperm) is a member of the jacalin-related family. Eur. J. Biochem. 2002, 269, 4335–4341. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, M.; Haraguchi, T.; Minami, Y.; Yagi, F.; Hiemori, K.; Tateno, H.; Hirabayashi, J. Two carbohydrate recognizing domains from Cycas revoluta leaf lectin show the distinct sugar-binding specificity-A unique mannooligosaccharide recognition by N-terminal domain. J. Biochem. 2016, 160, 27–35. [Google Scholar] [CrossRef]
- Animashaun, T.; Hughes, R.C. Bowringia milbraedii agglutinin. Specificity of binding to early processing intermediates of asparagine-linked oligosaccharide and use as a marker of endoplasmic reticulum glycoproteins. J. Biol. Chem. 1989, 264, 4657–4663. [Google Scholar] [PubMed]
- Siddiqui, S.; Hasan, S.; Salahuddin, A. Isolation and characterization of Cajanus cajan lectin. Arch. Biochem. Biophys. 1995, 319, 426–431. [Google Scholar] [CrossRef]
- Souza Teixeira, C.; da Silva, H.C.; de Moura, T.R.; Pereira-Júnior, F.N.; do Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; Delatorre, P.; Rocha, B.A.; Cavada, B.S. Crystal structure of the lectin of Camptosema pedicellatum: Implication of a conservative substitution at the hydrophobic subsite. J. Biochem. 2012, 152, 87–98. [Google Scholar] [CrossRef]
- Bezerra, G.A.; Viertlmayr, R.; Moura, T.R.; Delatorre, P.; Rocha, B.A.; do Nascimento, K.S.; Figueiredo, J.G.; Bezerra, I.G.; Teixeira, C.S.; Simões, R.C.; et al. Structural studies of an anti-inflammatory lectin from Canavalia boliviana seeds in complex with dimannosides. PLoS ONE 2014, 9, e97015. [Google Scholar] [CrossRef]
- Cavada, B.S.; Silva, M.T.L.; Osterne, V.J.S.; Pinto-Junior, V.R.; Nascimento, A.P.M.; Wolin, I.A.V.; Heinrich, I.A.; Nobre, C.A.S.; Moreira, C.G.; Lossio, C.F.; et al. Canavalia bonariensis lectin: Molecular bases of glycoconjugates interaction and antiglioma potential. Int. J. Biol. Macromol. 2018, 106, 369–378. [Google Scholar] [CrossRef]
- Sanz-Aparicio, J.; Hermoso, J.; Grangeiro, T.B.; Calvete, J.J.; Cavada, B.S. The crystal structure of Canavalia brasiliensis lectin suggests a correlation between its quaternary conformation and its distinct biological properties from Concanavalin A. FEBS Lett. 1997, 405, 114–118. [Google Scholar] [CrossRef]
- Agrawal, B.B.; Goldstein, I.J. Physical and chemical characterization of concanavalin A, the hemagglutinin from jack bean (Canavalia ensiformis). Bhiochim. Biophys. Acta 1967, 133, 376–379. [Google Scholar] [CrossRef]
- Kojima, K.; Ogawa, H.; Seno, N.; Matsumoto, I. Purification and characterization of Canavalia gladiata agglutinin. Carbohydr. Res. 1991, 213, 275–282. [Google Scholar] [CrossRef]
- Barroso-Neto, I.L.; Simões, R.C.; Rocha, B.A.; Bezerra, M.J.; Pereira-Junior, F.N.; Silva Osterne, V.J.; Nacscimento, K.S.; Nagano, C.S.; Delatorre, P.; Pereira, M.G.; et al. Vasorelaxant activity of Canavalia grandiflora seed lectin: A structural analysis. Arch. Biochem. Biophys. 2014, 543, 31–39. [Google Scholar] [CrossRef]
- Perez, G.; Perez, C.; Sousa-Cavada, B.; Moreira, R.; Richardson, M. Comparison of the amino acid sequence of the lectins from seeds of Dioclea lehmanni and Canavalia maritima. Phytochemistry 1991, 30, 2619–2621. [Google Scholar] [CrossRef]
- Osterne, V.J.S.; Silva-Filho, J.C.; Santiago, M.Q.; Pinto-Junior, V.R.; Almeida, A.C.; Barreto, A.A.G.C.; Wolin, I.A.V.; Nascimento, A.P.M.; Amorim, R.M.F.; Rocha, B.A.M.; et al. Structural characterization of a lectin from Canavalia virosa seeds with inflammatory and cytotoxic activities. Int. J. Biol. Macromol. 2017, 94, 271–282. [Google Scholar] [CrossRef]
- Vasconcelos, M.A.; Alves, A.C.; Carneiro, R.F.; Dias, A.H.; Martins, F.W.; Cajazeiras, J.B.; Nagano, C.S.; Teixeira, E.H.; Nascimento, K.S.; Cavada, B.S. Purification and primary structure of a novel mannose-specific lectin from Centrolobium microchaete Mart seeds. Int. J. Biol. Macromol. 2015, 81, 600–607. [Google Scholar] [CrossRef]
- Almeida, A.C.; Osterne, V.J.; Santiago, M.Q.; Pinto-Junior, V.R.; Silva-Filho, J.C.; Lossio, C.F.; Nascimento, F.L.; Almeida, R.P.; Teixeira, C.S.; Leal, R.B.; et al. Structural analysis of Centrolobium tomentosum seed lectin with inflammatory activity. Arch. Biochem. Biophys. 2016, 596, 73–83. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Barre, A.; Bemer, V.; Rougé, P.; Van Leuven, F.; Peumans, W.J. A lectin and a lectin-relared protein are the two most prominent proteins in the bark of yellow wood (Cladrastis lutea). Plant Mol. Biol. 1995, 29, 579–598. [Google Scholar] [CrossRef]
- Del Sol, F.G.; Cavada, B.S.; Calvete, J.J. Crystal structures of floribunda seed lectin at acidic and basic pHs. Insights into the structural basis of the pH-dependent dimer-tetramer transition. J. Struct. Biol. 2007, 158, 1–9. [Google Scholar] [CrossRef]
- Varejão, N.; Correia, M.T.; Foguel, D. Chqaracterization of the unfolding process of the tetrameric and dimeric forms of Cratylia mollis seed lectin (CRAMOLL1): Effects of natural fragmentation on protein stability. Biochemistry 2011, 50, 7330–7340. [Google Scholar] [CrossRef] [PubMed]
- Rocha, B.A.; Delatorre, P.; Oliveira, T.M.; Benevides, R.G.; Pires, A.F.; Sousa, A.A.; Souza, L.A.; Assreuy, A.M.; Debray, H.; de Azevedo, X.W.F.; et al. Structural basis for noth pro- and anti-inflammatory response induced by mannose-specific legume lectin from Cymbosema roseum. Biochimie 2011, 93, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.A.; Barros, A.C.; Stewart, J.C.; Pusztai, A. Isolation and characterization of a lectin from the seeds of Dioclea grandiflora (Mart.). Planta 1983, 158, 63–69. [Google Scholar] [CrossRef]
- Rozwarski, D.A.; Swami, B.M.; Brewer, C.F.; Sacchetini, J.C. Crystal structure of the lectin from Dioclea grandiflora complexed with core trimannoside of asparagine-linked carbohydrates. J. Biol. Chem. 1998, 273, 32818–32825. [Google Scholar] [CrossRef] [PubMed]
- Wah, D.A.; Romero, A.; Gallego del Sol, F.; Cavada, B.S.; Ramos, M.V.; Grangeiro, T.B.; Sampaio, A.H.; Calvete, J.J. Crystal structure of native and Cd/Cd-substituted Dioclea guianensis seed lectin. A novel manganese-binding site and structural basis of dimer-tetramer association. J. Mol. Biol. 2001, 310, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, K.S.; Santiago, M.Q.; Pinto-Junior, V.R.; Osterne, V.J.S.; Martins, F.W.V.; Nascimento, A.P.M.; Wolin, I.A.V.; Heinrich, I.A.; Martins, M.G.Q.; Silva, M.T.L.; et al. Structural analysis of Dioclea lasiocarpa lectin: A C6 cells apoptosis-inducing protein. Int. J. Biochem. Cell Biol. 2017, 92, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Leal, R.B.; Pinto-Junior, V.R.; Osterne, V.J.S.; Wolin, I.A.V.; Nascimento, A.P.M.; Neco, A.H.B.; Araripe, D.A.; Welter, P.G.; Neto, C.C.; Correia, J.L.A.; et al. Crystal structure of DlyL, a mannose-specific lectin from Dioclea lasiophylla Mart. Ex Benth seeds that display cytotoxic effects against C6 glioma cells. Int. J. Biol. Macromol. 2018, 114, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Junior, V.R.; Osterne, V.J.; Santiago, M.Q.; Correia, J.L.; Pereira-Junior, F.N.; Leal, R.B.; Pereira, M.G.; Chicas, L.S.; Nagano, C.S.; Rocha, B.A.; et al. Structural studies of a vasorelaxant lectin from Dioclea reflexa Hook seeds: Crystal structure, molecular docking and dynamics. Int. J. Biol. Macromol. 2017, 98, 12–23. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.M.; Delatorre, P.; da Rocha, B.A.; de Souza, E.P.; Nascimento, K.S.; Bezerra, G.A.; Moura, T.R.; Benevides, R.G.; Bezerra, E.H.; Moreno, F.B.; et al. Crystal structure of Dioclea rostrata lectin: Insights into understanding the pH-dependent dimer-tetramer equilibrium and the structural basis for carbohydrate recognition in Diocleinae lectins. J. Struct. Biol. 2008, 164, 177–182. [Google Scholar] [CrossRef]
- Barroso-Neto, I.L.; Delatorre, P.; Teixeira, C.S.; Correia, J.L.; Cajazeiras, J.B.; Pereira, R.I.; Nascimento, K.S.; Laranjeira, E.P.; Pires, A.F.; Assreuy, A.M.; et al. Structural analysis of a Dioclea sclerocarpa lectin: Study on the vasorelaxant properties of Dioclea lectins. Int. J. Biol. Macromol. 2016, 82, 464–470. [Google Scholar] [CrossRef]
- Bezerra, M.J.; Rodrigues, N.V.; Pires Ade, F.; Bezerra, G.A.; Nobre, C.B.; Alencar, K.L.; Soares, P.M.; do Nascimento, K.S.; Nagano, C.S.; Martins, J.L.; et al. Crystal structure of Dioclea violacea lectin and a comparative study of vasorelaxant properties with Dioclea rostrata lectin. Int. J. Biochem. Cell Biol. 2013, 45, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Batista da Nóbrega, R.; Rocha, B.A.; Gadelha, C.A.; Santi-Gadelha, T.; Pires, A.F.; Assreuy, A.M.; Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; Cavada, B.S.; et al. Structure of Dioclea virgata lectin: Relations between carbohydrate binding site and nitric oxide production. Biochimie 2012, 94, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Rangel, T.B.; Rocha, B.A.; Bezerra, G.A.; Assreuy, A.M.; Pires Ade, F.; do Nascimento, A.S.; Bezerra, M.J.; do Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; et al. Crystal structure of a pro-inflammatory lectin from the seeds of Dioclea wilsonii Standl. Biochimie 2012, 94, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Cavada, B.S.; Richardson, M.; Yarwood, A.; Père, D.; Rougé, P. The amino acid sequences of the α subunits of the lectins from Lathyrus cicera, L. aphaca and L. articulatus. Phytochemistry 1986, 25, 115–118. [Google Scholar] [CrossRef]
- Yarwood, A.; Richardson, M.; Cavada, B.S.; Père, D.; Rougé, P. The amino acid sequences of the α subunits of the lectins from the seeds of Lathyrus hirsutus and Lathyrus tingitanus. Phytochemistry 1986, 25, 2109–2112. [Google Scholar] [CrossRef]
- Yarwood, A.; Richardson, M.; Morphet, B.; Westby, M.; Père, D.; Rougé, P. The amino acid sequences of two atypical single-chain Vicieae isolectins from seeds of Lathyrus nissolia L. Phytochemistry 1988, 27, 1719–1721. [Google Scholar] [CrossRef]
- Bourne, Y.; Abergel, C.; Cambillau, C.; Frey, M.; Rougé, P.; Fontecilla-Camps, J.C. X-ray crystal structure determination and refinement at 1.9 Å resolution of isolectin I from the seeds of Lathyrus ochrus. J. Mol. Biol. 1990, 214, 571–584. [Google Scholar] [CrossRef]
- Kolberg, J. Isolation and partial characterization of a mitogenic lectin from Lathyrus odoratus seeds. Acta Pathol. Microbiol. Scand. C 1978, 86C, 99–104. [Google Scholar] [CrossRef]
- Sletten, K.; Kolberg, J. The primary structure of the α chain of a mitogenic lectin from the seeds of Lathyrus sativus. Hoppe Seylers Z. Physiol. Chem. 1983, 364, 1047–1051. [Google Scholar] [CrossRef]
- Richardson, M.; Yarwood, A.; Rougé, P. The amino acid sequence of an atypical single-chain lectin from seeds of Lathyrus sphaericus. FEBS Lett. 1987, 216, 145–150. [Google Scholar] [CrossRef]
- Tichá, M.; Zeineddine, I.; Kocourek, J. Studies on lectins. XLVIII. Isolation and characterization of lectins from the seeds of Lathyrus odoratus L. and Lathyrus silvestris L. Acta Biol. Med. Ger. 1980, 39, 649–655. [Google Scholar] [PubMed]
- Foriers, A.; Van Driessche, E.; De Neve, R.; Kanarek, L.; Strosberg, A.D. The subunit structure and N-terminal sequences of the α- and β-subunits of the lentil lectin (Lens culinaris). FEBS Lett. 1977, 75, 237–240. [Google Scholar] [CrossRef]
- Gao, S.; An, J.; Wu, C.F.; Gu, Y.; Chen, F.; Yu, Y.; Wu, Q.Q.; Bao, J.K. Effect of amino acid residue and oligosaccharide chain chemical modifications on spectral and hemagglutinating activity of Millettia dielsiana Harms. ex Diels. lectin. Acta Biochim. Biophys. Sin. (Shangai) 2005, 37, 47–54. [Google Scholar] [CrossRef]
- Young, N.M.; Williams, R.E.; Roy, C.; Yaguchi, M. Structural comparison of the lectin from sainfoin (Onobrychis viciifolia) with concanavalin A and other D-mannose specific lectins. Can. J. Biochem. 1982, 60, 933–941. [Google Scholar] [CrossRef]
- Cavada, B.S.; da Silva, L.I.; Ramos, M.V.; Galvani, F.R.; Grangeiro, T.B.; Leite, K.B.; Assreuy, A.M.; Cajazeiras, J.B.; Calvete, J.J. Seed lectin from Pisum arvense: Isolation, biochemical characterization and amino acid sequence. Protein Pept. Lett. 2003, 10, 607–617. [Google Scholar] [CrossRef]
- Einspahr, H.; Pareks, E.H.; Suguna, K.; Subramanian, E.; Suddath, F.L. The crystal structure of pea lectin at 3.0-Å resolution. J. Biol. Chem. 1986, 261, 16518–16527. [Google Scholar]
- Loris, R.; Imberty, A.; Beeckmans, S.; Van Driessche, E.; Read, J.S.; Bouckaert, J.; De Greve, H.; Buts, L.; Wyns, L. Crystal structure of Pterocarpus angolensis lectin in complex with glucose, sucrose, and turanose. J. Biol. Chem. 2003, 278, 16297–16303. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, B.; Zhang, Z.T.; Zhou, T.T.; Bian, H.J.; Min, M.W.; Liu, Y.H.; Chen, J.; Bao, J.K. A mannose-binding lectin from Sophora flavescens induces apoptosis in HeLa cells. Phytomedicine 2008, 15, 867–875. [Google Scholar] [CrossRef]
- Naeem, A.; Ahmad, E.; Ashraf, M.T.; Khan, R.H. Purification and characterization of mannose/glucose-specific lectin from seeds of Trigonella foenumgraecum. Biochemistry (Mosc) 2007, 72, 44–48. [Google Scholar] [CrossRef]
- Baumann, C.M.; Strosberg, A.D.; Rüdiger, H. Purification and characterization of a mannose/glucose-specific lectin from Vicia cracca. Eur. J. Biochem. 1982, 122, 105–110. [Google Scholar] [CrossRef]
- Fornstedt, N.; Porath, J. Characterization studies on a new lectin found in seeds of Vicia ervilia. FEBS Lett. 1975, 57, 187–191. [Google Scholar] [CrossRef]
- Reeke, G.N., Jr.; Becker, J.W. Three-dimensional structure of favin: Saccharide binding-cyclic permutation in leguminous lectins. Science 1986, 234, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, G.; Schiltz, E.; Rüdiger, H. The amino-acid sequence of the alpha subunit of the mitogenic lectin from Vicia sativa. Eur. J. Biochem. 1981, 113, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.C.; Bari, A.U.; Rocha, B.A.; Nascimento, K.S.; Ponte, E.L.; Pires, A.F.; Delatorre, P.; Teixeira, E.H.; Debray, H.; Assreuy, A.M.; et al. Purification and primary structure of a mannose/glucose-binding lectin from Parkia biglobosa Jacq. seeds with antinociceptive and anti-inflammatory properties. J. Mol. Recognit. 2013, 26, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Farias, C.M.; Del Sol, F.G.; Santos, C.F.; Grangeiro, T.B.; Nagano, C.S.; Cavada, B.S.; Calvete, J.J. The amino acid sequence of the glucose/mannose-specific lectin isolated from Parkia platycephala seeds reveals three tandemly arranged jacalin-related domains. Eur. J. Biochem. 2001, 268, 4414–4422. [Google Scholar] [CrossRef] [PubMed]
- Cavada, B.S.; Araripe, D.A.; Silva, I.B.; Pinto-Junior, V.R.; Osterne, V.J.S.; Neco, A.H.B.; Laranjeira, E.P.P.; Lossio, C.F.; Correia, J.L.A.; Pires, A.F.; et al. Structural studies and nociceptive activity of a native lectin from Platypodium elegans seeds (nPELa). Int. J. Biol. Macromol. 2018, 107, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Junior, F.N.; Silva, H.C.; Freitas, B.T.; Rocha, B.A.; Nascimento, K.S.; Nagano, C.S.; Leal, R.B.; Sampaio, A.H.; Cavada, B.S. Purification and characterization of a mannose/N-acetyl-d-glucosamine-specific lectin from the seeds of Platymiscium floribundum Vogel. J. Mol. Recognit. 2012, 25, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Ashida, H.; Uemura, N.; Kushibe, S.; Ozaki, T.; Yoshida, M. Purification and characterization of a mannose/glucose-specific lectin from Castanea crenata. Phytochemistry 1998, 49, 667–673. [Google Scholar] [CrossRef]
- Da Silva, T.A.; Oliveira-Brito, P.K.M.; Gonçalves, T.E.; Vendruscolo, P.E.; Roque-Barreira, M.C. ArtinM mediates murine T cell activation and induces cell death in Jurkat human leukemic T cells. Int. J. Mol. Sci. 2017, 18, 1400. [Google Scholar] [CrossRef]
- Pereira-da-Silva, G.; Roque-Barreira, M.C.; Van Damme, E.J. ArtiM: A rational substitution for the names artocarpin and KM+. Immunol. Lett. 2008, 119, 114–115. [Google Scholar] [CrossRef]
- de Sousa, F.D.; da Silva, B.B.; Furtado, G.P.; Carneiro, I.S.; Lobo, M.D.P.; Guan, Y.; Guo, J.; Coker, A.R.; Lourenzoni, M.R.; Guedes, M.I.F.; et al. Frutapin, a lectin from Artocarpus incisa (breadfruit): Cloning, expression and molecular insights. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, M.; Abdul-Rahman, P.S.; Isaacs, N.W.; Hashim, O.H.; Copgdell, R.J. Crystallization and initial X-ray diffraction analysis of a mannose-binding lectin from champedak. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, M.; Abdul-Rahman, P.S.; Othman, S.; Hashim, O.H.; Cogdell, R.J. Structures and binding specificity of galactose- and mannose-binding lectins from champedak: Differences from jacfruit lectins. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 709–716. [Google Scholar] [CrossRef]
- Rosa, J.C.; De Oliveira, P.S.; Garratt, R.; Beltramini, L.; Resing, K.; Roque-Barreira, M.C.; Greene, L.J. KM+, a mannose-binding lectin from Artocarpus integrifolia: Amino acid sequence, predicted tertiary structure, carbohydrate recognition, and analysis of the β-prism fold. Protein Sci. 1999, 8, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Misquith, S.; Rani, P.G.; Surolia, A. Carbohydrate binding specificity of the B-cell maturation mitogen from Artocarpus integrifolia seeds. J. Biol. Chem. 1994, 269, 30393–30401. [Google Scholar] [PubMed]
- Bourne, Y.; Houlès-Astoul, C.; Zamboni, V.; Peumans, W.J.; Menu-Bouaouiche, L.; Van Damme, E.J.M.; Barre, A.; Rougé, P. Structural basios for the unusual carbohydrate-binding specificity of jacalin towards galactose and mannose. Biochem. J. 2002, 364, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Jeyaprakash, A.A.; Jayashree, G.; Mahanta, S.K.; Swaminathan, C.P.; Sekar, K.; Surolia, A.; Vijayan, M. Structural basis for the energetics of jacalin-sugar interactions: Promiscuity versus specificity. J. Mol. Biol. 2005, 347, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Chjowdhury, S.; Ahmed, H.; Chatterjee, B.P. Chemical modificatrion studies of Artocarpus lakoocha lectin artocarpin. Biochimie 1991, 73, 563–571. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Hause, B.; Hu, J.; Barre, A.; Rougé, P.; Proost, P.; Peumans, W.J. Two distinct jacalin-related lectins with a different specificity and subcellular location are major vegetative storage proteins in the bark of the black mulberry tree. Plant Physiol. 2002, 130, 757–769. [Google Scholar] [CrossRef]
- Bourne, Y.; Zamboni, V.; Barre, A.; Peumans, W.J.; Van Damme, E.J.; Rougé, P. Helianthus tuberosus lectin reveals a widespread scaffold for mannose-binding lectins. Structure 1999, 7, 1473–1482. [Google Scholar] [CrossRef]
- Beneteau, J.; Renard, D.; Marché, L.; Douville, E.; Lavenant, L.; Rahbé, Y.; Dupont, D.; Vilaine, F.; Dinant, S. Binding properties of the N-acetylglucosamine and high-mannose N-glycan PP2-A1 phloem lectin in Arabidopsis. Plant Physiol. 2010, 153, 1345–1361. [Google Scholar] [CrossRef]
- Peng, H.; Lv, H.; Wang, Y.; Liu, Y.H.; Li, C.Y.; Meng, L.; Chen, F.; Bao, J.K. Clematis montana lectin, a novel mannose-binding lectin from traditional Chinese medicine with antiviral and apoptosis-inducing activities. Peptides 2009, 30, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Koike, T.; Titani, K.; Suzuki, M.; Beppu, H.; Kuzuya, H.; Maruta, K.; Shimpo, K.; Fujita, K. The complete amino acid sequence of a mannose-binding lectin from "Kidachi Aloe" (Aloe arborescens Miller var. natalensis Berger). Biochem. Biophys. Res. Commun. 1995, 214, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhou, X.; Pang, Y.; Gao, H.; Fei, J.; Shen, G.A.; Wang, J.; Li, X.; Sun, X.; Tang, K. Cloning and characterization of an agglutinin gene from Arisaema lobatum. Biosci. Rep. 2005, 25, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yao, J.; Liao, Z.; Zhang, H.; Chen, F.; Wang, L.; Lu, Y.; Sun, X.; Yu, S.; Tang, K. Molecular cloning of a novel mannose-binding lectin gene from Arisaema heterophyllum. Plant Sci. 2003, 165, 55–60. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Goossens, K.; Smeets, K.; Van Leuven, F.; Verhaert, P.; Peumans, W.J. The major tuber storage protein of araceae species is a lectin. Characterization and molecular cloning of the lectin from Arum maculatum L. Plant Physiol. 1995, 107, 1147–1158. [Google Scholar] [CrossRef]
- Pereira, P.R.; Winter, H.C.; Vericimo, M.A.; Meagher, J.L.; Stckey, J.A.; Goldstein, I.J.; Paschoalin, V.M.; Silva, J.T. Structural analysis and binding properties of isoforms of tarin, the GNA-related lectin from Colocasia esculenta. Biochim. Biophys. Acta 2015, 1854, 20–30. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakamoto, H.; Tateno, H.; Hirabayashi, J.; Oguri, S. Purification, characterization, and molecular cloning of lectin from winter buds of Lysichiton camtschatcensis (L.) Schott. Biosci. Biotechnol. Biochem. 2012, 76, 25–33. [Google Scholar] [CrossRef]
- Yao, J.H.; Zhao, X.Y.; Liao, Z.H.; Lin, J.; Chen, Z.H.; Chen, F.; Song, J.; Sun, X.F.; Tang, K.X. Cloning and molecular characterization of a novel lectin gene from Pinellia ternata. Cell Res. 2003, 13, 301–308. [Google Scholar] [CrossRef]
- Shetty, K.N.; Bhat, G.G.; Inamdar, S.R.; Swamy, B.M.; Suguna, K. Crystal structure of a β-prism II lectin from Remusatia vivipara. Glycobiology 2012, 22, 56–69. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, X.; Liu, J.; Li, J.; Sun, Y.; Liu, Z.; Liu, J.; Van Damme, E.; Balzarini, J.; Bao, J. A novel mannose-binding tuber lectin from Typhonium divaricatum (L.) Decne (family Araceae) with antiviral activity against HSV-II and anti-proliferative effect on human cancer cell lines. J. Biochem. Mol. Biol. 2007, 40, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Mo, H.; Rice, K.G.; Evers, D.L.; Winter, H.C.; Peumans, W.J.; Van Damme, E.J.; Goldstein, I.J. Xanthosoma sagittifolium tubers contain a lectin with two different types of carbohydrate-binding sites. J. Biol. Chem. 1999, 274, 33300–33305. [Google Scholar] [CrossRef]
- Chen, Z.; Pang, Y.; Liu, X.; Wang, X.; Deng, Z.; Sun, X.; Tang, K. Molecular cloning and characterization of a novel mannose-binding lectin cDNA from Zantedeschia aethiopica. Biocell 2005, 29, 187–193. [Google Scholar]
- Liu, B.; Peng, H.; Yao, Q.; Li, J.; Van Damme, E.; Balzarini, J.; Bao, J.K. Bioinformatics analyses of the mannose-binding lectins from Polygonatum cyrtonema, Ophiopogon japonicus and Liparis noversa with antiproliferative and apoptosis-inducing activities. Phytomedicine 2009, 16, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Bao, J.; Zhu, D.; Zhang, Y.; Wang, D.C. Crystal structure of a novel anti-HIV mannose-binding lectin from Polygonatum cyrtonema Hua with unique ligand-binding property and super-structure. J. Struct. Biol. 2010, 171, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Barre, A.; Rougé, P.; Van Leuven, F.; Balzarini, J.; Peumans, W.J. Molecular cloning of the lectin and a lectin-related protein from common Solomon’s seal (Polygonatum multiflorum). Plant Mol. Biol. 1996, 31, 657–672. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, H.L.; Zhang, Z.T.; Liu, J.J.; Li, W.W.; Ming, H.; Bao, J.K. Characterization, molecular cloning, and in silico analysis of a novel mannose-binding lectin from Polygonatum odoratum (Mill.) with anti-HSV-II and apoptosis-inducing activities. Phytomedicine 2011, 18, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Roig-Zamboni, V.; Barre, A.; Peumans, W.J.; Astoul, C.H.; Van Damme, E.J.; Rougé, P. The crystal structure of the Calystegia sepium agglutinin reveals a novel quaternary arrangement of lectin subunits with a β-prism fold. J. Biol. Chem. 2004, 279, 527–533. [Google Scholar] [CrossRef]
- Chang, W.C.; Liu, K.L.; Hsu, F.C.; Jeng, S.T.; Cheng, Y.S. Ipomoelin, a jacalin-related lectin with a compact tetrameric association and versatile carbohydrate binding properties regulated by its N terminus. PLoS ONE 2012, 7, e40618. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Saurabh, S.; Singh, R.; Rai, P.; Dubey, N.K.; Chandrashekar, K.; Negi, K.S.; Tuli, R.; Singh, P.K. Purification and characterization of a lectin with high hemagglutination property isolated from Allium altaicum. Protein J. 2011, 30, 374–383. [Google Scholar] [CrossRef]
- Mo, H.; Van Damme, E.J.; Peumans, W.J.; Goldstein, I.J. Purificartion and characterization of a mannose-specific lectin from shallot (Allium ascalonicum) bulbs. Arch. Biochem. Biophys. 1993, 306, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Smeets, K.; Engelborghs, I.; Aelbers, H.; Balzarini, J.; Pusztai, A.; van Leuven, F.; Goldstein, I.J.; Peumans, W.J. Cloning and characterization of the lectin cDNA clones from onion, shallot and leek. Plant. Mol. Biol. 1993, 23, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Smmets, K.; Torrekens, S.; van Leuven, F.; Goldstain, I.J.; Peumans, W.J. The closely relataed homomeric and heterodimeric mannose-binding lectins from garlic are encoded by one-domain and two-domain lectin genes, respectively. Eur. J. Biochem. 1992, 206, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.; Yu, H.; Chen, C.M.; Sun, S.S.; Ooi, V.E. Isolation and characterization of a bioactive mannose-binding protein from the Chinese chive Allium tuberosum. J. Agric. Food Chem. 2002, 50, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Smeets, K.; Torrekens, S.; Van Leuven, F.; Peumans, W.J. The mannose-specific lectins from ramsons (Allium ursinum L.) are encoded by three sets of genes. Eur. J. Biochem. 1993, 217, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-F.; An, J.; He, X.-J.; Deng, J.; Hong, Z.-X.; Liu, C.; Hong-Zhou, L.; Li, Y.-J.; Wang, C.-J.; Chen, F.; Bao, J. Molecular cloning of a novel mannose-binding lectin gene from bulbs of Amaryllis vittata (Amaryllidaceae). Sheng Wu Yi Xue Gong Cheng Xue Za Zhi 2004, 46, 1301–1306. (In Chinese) [Google Scholar]
- Van Damme, E.J.M.; Smeets, K.; Van Leuven, F.; Peumans, W.J. Molecular cloning of mannose-binding lrctins from Clivia miniata. Plant Mol. Biol. 1994, 24, 825–830. [Google Scholar] [CrossRef]
- Chai, Y.; Pang, Y.; Liao, Z.; Zhang, L.; Sun, X.; Lu, Y.; Wang, S.; Tang, K. Molecular cloning and characterization of a mannose-binding lectin gene from Crinum asiaticum. J. Plant Physiol. 2003, 160, 913–920. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Kaku, H.; Perini, F.; Goldstein, I.J.; Peeters, B.; Yagi, F.; Decock, B.; Peumans, W.J. Biosynthesis, primary structure and molecular cloning of snowdrop (Galanthus nivalis L.) lectin. Eur. J. Biochem. 1991, 202, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Kaku, H.; Van Damme, E.J.; Peumans, W.J.; Goldstein, I.J. Carbohydrate-binding specificity of the daffodil (Narcissus pseudonarcissus) and amaryllis (Hippeastrum hybr.) bulb lectins. Arch. Biochem. Biophys. 1990, 279, 298–304. [Google Scholar] [CrossRef]
- Antoniuk, L.L.A.; Antoniuk, V.O. Interaction of immobilized lectin from Leucojum vernum L. with polysaccharides and glycoproteins. Ukrainskii Biokhimicheskii Zhurnal 1978, 65, 69–77. (In Ukrainian) [Google Scholar]
- Wu, C.-F.; Li, J.; An, J.; Chang, L.-Q.; Che, F.; Bao, J.-K. Purification, biological activities, and molecular cloning of a novel mannose-binding lectin from bulbs of Zephyranthes candida herb (Amaryllidaceae). J. Integr. Plant Biol. 2006, 48, 223–231. [Google Scholar] [CrossRef]
- Bao, J.; Wu, C.; An, J.; Gao, S.; Zhao, X.; Chang, L.; Rong, Y.; Wang, C.; Chen, F. Molecular cloning and analysis of a monocot mannose-binding agglutinin from Zephyranthes grandiflora (family Amaryllidaceae). Sheng Wu Yi Xue Gong Cheng Xue Za Zhi 2004, 21, 812–818. (In Chinese) [Google Scholar] [PubMed]
- Liu, J.; Xu, X.; Liu, J.; Balzarini, J.; Luo, Y.; Kong, Y.; Li, J.; Chen, F.; Van Damme, E.J.; Bao, J. A novel tetrameric lectin from Lycoris aurea with four mannose binding sites per monomer. Acta Biochim. Pol. 2007, 54, 159–166. [Google Scholar] [PubMed]
- Zhao, X.; Yao, J.; Sun, X.; Tang, K. Molecular cloning and characterization of a novel lectin gene from Lycoris radiata. DNA Seq. 2003, 14, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Ohizumi, Y.; Gaidamashvili, M.; Ohwada, S.; Matsuda, K.; Kominami, J.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Naganuma, T.; Ogawa, T.; Muramoto, K. Mannose-binding lectin from yam (Dioscorea batatas) tubers with insecticidal properties against Helicoverpa armigera (Lepidoptera:Noctuidae). J. Agric. Food Chem. 2009, 57, 2896–2902. [Google Scholar] [CrossRef]
- Sharma, M.; Vishwanathreddy, H.; Sindhura, B.R.; Kamalanathan, A.S.; Swamy, B.M.; Inamdar, S.R. Purification, characterization and biological significance of mannose binding lectin from Dioscorea bulbifera bulbils. Int. J. Biol. Macromol. 2017, 101, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Nakayama, K.; Abdul-Rahman, B.; Kinoshita, M.; Hashimoto, O.; Kawasaki, N.; Hayakawa, T.; Kakehi, K.; Tomiya, N.; Lee, Y.C. Crocus sativus lectin recognizes Man3GlncNAc in the N-glycan core structure. J. Biol. Chem. 2000, 275, 26772–27779. [Google Scholar] [PubMed]
- Escribano, J.; Rubio, A.; Alvarez-Ortí, M.; Molina, A.; Fernández, J.A. Purification and characterization of a mannan-binding lectin specifically expressed in corms of saffrom plant (Crocus sativus L.). J. Agric. Food Chem. 2000, 48, 457–463. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Astoul, C.H.; Barre, A.; Rougé, P.; Peumans, W.J. Cloning and characterization of a monocot mannose-binding lectin from Crocus vernus (family Iridaceae). Eur. J. Biochem. 2000, 267, 5067–5077. [Google Scholar] [CrossRef]
- Xu, X.; Wu, C.; Liu, C.; Luo, C.; Luo, Y.; Li, J.; Zhao, X.; Van Damme, E.; Bao, J. Purification and characterization of a mannose-binding lectin from the rhizomes of Aspidistra elatior Blume with antiproliferative activity. Acta Biochimi. Biophys. Sin. (Shanghai) 2007, 39, 507–519. [Google Scholar] [CrossRef]
- Ooi, L.S.; Ho, W.S.; Ngai, K.L.; Tian, L.; Chan, P.K.; Sun, S.S.; Ooi, V.E. Narcissus tazetta lectin shows strong inhibitory effects against respiratory syncytial virus, influenza A (H1N1, H3N2, H5N1) and B viruses. J. Biosci. 2010, 35, 95–103. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Armand-Ugon, M.; Bastida, J.; Viladomat, F.; Esté, J.A.; Stewart, D.; Codina, C. Anti-human immunodeficiency virus type 1 (HIV-1) activity of lectins from Narcissus species. Planta Med. 2003, 69, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Briké, F.; Winter, H.C.; Van Leuven, F.; Goldstein, I.J.; Peumans, W.J. Molecular cloning of two different mannose-binding lectins from tulip bulbs. Eur. J. Biochem. 1996, 236, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.; Sun, S.S.; Wang, H.; Ooi, V.E. New mannose-binding lectin isolated from the rhizome of Sarsaparilla Smilax glabra Roxb. (Liliaceae). J. Agric. Food Chem. 2004, 52, 6091–6095. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.M.; Wood, S.D.; Reynolds, C.D.; Rizkallah, P.J.; Peumans, W.J.; Van Damme, E.J.; Allen, A.K. Purification, crystalolization and preliminary X-ray analysis of a mannose-binding lectin from bluebell (Scilla campanulata) bulbs. Acta Crystallogr. D Biol. Crystallogr. 1996, 52, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Vijayan, M. Influence of glycosidic linkage on the nature of carbohydrate binding in β-prism I fold lectins: An X-ray and molecular dynamics investigation on banana lectin-carbohydrate complexes. Glycobiology 2011, 21, 23–33. [Google Scholar] [CrossRef]
- Singh, D.D.; Saikrishnan, K.; Kumar, P.; Surolia, A.; Sekar, K.; Vijayan, M. Unusual sugar specificity of banana lectin from Musa paradisiaca and its probable evolutionary origin. Crystallographic and modelling studies. Glycobiology 2005, 15, 1025–1032. [Google Scholar] [CrossRef]
- Ooi, L.S.; Sun, S.S.; Ooi, V.E. Purification and characterization of a new antiviral protein from the leaves of Pandanus amaryllifolius (Pandanaceae). Int. J. Biochem. Cell Biol. 2004, 36, 1440–1446. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Smeets, K.; Torrekens, S.; Van Leuven, F.; Peumans, W.J. Characterization and molecular cloning of mannose-binding lectins from the Orchidaceae species Listera ovata, Epipactis helleborine and Cymbidium hybrid. Eur. J. Biochem. 1994, 221, 769–777. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, X.; Tang, K. Cloning and expression of a novel cDNA encoding a mannose-binding lectin from Dendrobium officinale. Toxicon 2005, 45, 535–540. [Google Scholar] [CrossRef]
- Liu, W.; Yang, N.; Ding, J.; Huang, R.H.; Hu, Z.; Wang, D.C. Structural mechanism governing the quaternary organization of monocot mannose-binding lectin revealed by the novel monomeric structure of an orchid lectin. J. Biol. Chem. 2005, 280, 14865–14976. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Peumans, W.J.; Barre, A.; Astoul, C.H.; Rovira, P.; Rougé, P.; Proost, P.; Truffa-Bachi, P.; Jalali, A.A.; Van Damme, E.J. Isolation and characterization of a jacalin-related mannose-binding lectin from salt-stressed rice (Oryza sativa) plants. Planta 2000, 210, 970–978. [Google Scholar] [PubMed]
- Do Nascimento-Neto, L.G.; Carneiro, R.F.; da Silva, S.R.; Rocha da Silva, B.; Arruda, F.V.S.; Carneiro, V.A.; do Nascimento, K.S.; Saker-Sampaio, S.; da Silva, V.A., Jr.; Figueiredo Porto, A.L.; et al. Characterization of isoforms of the lectin isolated from the red algae Bryothamnion seaforthii and its pro-healing effect. Mar. Drugs 2012, 10, 1936–1954. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Costa, F.H.; Saker-Sampaio, S.; Murciano, M.P.; Nagano, C.S.; Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Bloch, C., Jr.; Silveira, S.B.; et al. The amino acid sequence of the agglutinin isolated from the red marine alga Bryothamnion triquetrum defines a novel lectin structure. Cell Mol. Life Sci. 2000, 57, 343–350. [Google Scholar] [CrossRef]
- Hung, L.D.; Hirayama, M.; Ly, B.M.; Hori, K. Purification, primary structure, and biological activity of the high-mannose N-glycan-specific lectin from cultivated Eucheuma denticulatum. J. Appl. Phycol. 2015, 27, 1657–1669. [Google Scholar] [CrossRef]
- Sugahara, T.; Ohama, Y.; Fukuda, A.; Hayashi, M.; Kawakubo, A.; Kato, K. The cytotoxic effect of Eucheuma serrata agglutinin (ESA) on cancer cells and its application to molecular probe for drug delivery system using lipid vesicles. Cytotechnology 2001, 36, 93–99. [Google Scholar] [CrossRef]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterizatiopn of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef]
- Zlółkowska, N.E.; O’Keefe, B.R.; Mori, T.; Zhu, C.; Giomarelli, B.; Vojdani, F.; Palmer, K.E.; McMahon, J.B.; Wlodawer, A. Domain-swapped structure of the potent antiviral protein griffithsin and its mode of carbohydrate binding. Structure 2006, 14, 1127–1138. [Google Scholar] [CrossRef]
- Nascimento, K.S.; Nagano, C.S.; Nunes, E.V.; Rodrigues, R.F.; Goersch, G.V.; Cavada, B.S.; Calvete, J.J.; Saker-Sampaio, S.; Farias, W.R.; Sampaio, A.H. Isolation and characterization of a new agglutinin from the red marine alga Hypnea cervicornis J. Agardh. Biochem. Cell Biol. 2006, 84, 49–54. [Google Scholar] [CrossRef]
- Nagano, C.S.; Moreno, F.B.; Bloch, C., Jr.; Prates, M.V.; Calvete, J.J.; Saker-Sampaio, S.; Farias, W.R.; Tavares, T.D.; Nascimento, K.S.; Grangeiro, T.B.; et al. Purification and characterization of a new lectin from the red marine alga Hypnea musciformis. Protein Pept. Lett. 2002, 9, 159–166. [Google Scholar] [CrossRef]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.D.; Sato, Y.; Hori, K. High-mannose N-glycan-specific lectin from the red alga Kappaphycus striatum (Carrageenophyte). Phytochemistry 2011, 72, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High-mannose-binding lectin with preference for the cluster of α1-2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga Halimeda renschii exhibits a potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef] [PubMed]
- Wohlschlager, T.; Butschi, A.; Grassi, P.; Sutov, G.; Gauss, R.; Hauck, D.; Schmieder, S.S.; Knobel, M.; Titz, A.; Dell, A.; et al. Methylated glycans as conserved targets of animal and fungal innate defense. Proc. Natl. Acad. Sci. USA 2014, 111, E2787–E2796. [Google Scholar] [CrossRef]
- Sommer, R.; Makshakova, O.N.; Wohlschlager, T.; Hutin, S.; Marsh, M.; Titz, A.; Künzler, M.; Varrot, A. Crystal structure of fungal tectonin in complex with O-methylated glycans suggest key role in innate immune defense. Structure 2018, 26, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Francis, F.; Jaber, K.; Colinet, F.; Portetelle, D.; Haubruge, E. Purification of a new fungal mannose-specific lectin from Penicillium chrysogenum and its aphicidal properties. Fungal Biol. 2011, 115, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Veelders, M.; Brückner, S.; Ott, D.; Unverzagt, C.; Mösch, H.U.; Essen, L.O. Structural basis of flocculin-mediated social behavior in yeast. Proc. Natl. Acad. Sci. USA 2010, 107, 22511–22516. [Google Scholar] [CrossRef]
- Goossens, K.V.; Ielasi, F.S.; Nookaew, I.; Stals, I.; Alonso-Sarduy, L.; Daenen, L.; Van Mulders, S.E.; Stassen, C.; van Eisjden, R.G.; Siewers, V.; et al. Molecular mechanism of flocculation self-recognition in yeast and its role in mating and survival. MBio 2015, 6, e00427-15. [Google Scholar] [CrossRef]
- Olson, L.J.; Orsi, R.; Peterson, F.C.; Parodi, A.J.; Kim, J.J.; D’Alessio, C.; Dahms, N.M. Crystal structure and functional analyses of the lectin domain of glucosidase II: Insights into oligomannose recognition. Biochemistry 2015, 54, 4097–4111. [Google Scholar] [CrossRef]
- Suzuki, T.; Sugiyama, K.; Hirai, H.; Ito, H.; Morita, T.; Dohra, H.; Murata, T.; Usui, T.; Tateno, H.; Hirabayashi, J.; et al. Mannose-specific lectin from the mushroom Hygrophorus russula. Glycobiology 2012, 22, 616–629. [Google Scholar] [CrossRef]
- Shimokawa, M.; Fukudome, A.; Yamashita, R.; Minami, Y.; Yagi, F.; Tateno, H.; Hirabayashi, J. Characterization and cloning of GNA-like lectin from the mushroom Marasmius oreades. Glycoconj. J. 2012, 29, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Viscomi, A.R.; Jee, J.G.; Ottonello, S.; Gronenborn, A.M. The evolutionary conserved family of cyanovirin-N homologs: Structures and carbohydrate specificity. Structure 2008, 16, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Loris, R.; Van Overberge, D.; Dao-Thi, M.H.; Poortmans, F.; Maene, N.; Wyns, L. Structural analysis of two crystal forms of lentil lectin at 1.8 Å resolution. Proteins 1994, 20, 330–346. [Google Scholar] [CrossRef] [PubMed]
- Hardman, K.D.; Ainsworth, C.F. Structure of concanavalin A at 2.4-Å resolution. Biochemistry 1972, 11, 4910–4919. [Google Scholar] [CrossRef] [PubMed]
- Olsen, L.R.; Dessen, A.; Gupta, D.; Sabesan, S.; Sacchettini, J.C.; Brewer, C.F. X-ray crystallographic studies of unique cross-linked lattices between four isomeric biantennary oligosaccharides and soybean agglutinin. Biochemistry 1997, 36, 15073–15080. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Das, K.; Ravishankar, R.; Suguna, K.; Surolia, A.; Vijayan, M. Conformation, protein-carbohydrate interactions and a novel subunit association in the refined structure of peanut lectin-lactose complex. J. Mol. Biol. 1996, 259, 281–296. [Google Scholar] [CrossRef]
- Elgavish, S.; Shaanan, B. Structures of the Erythrina corallodendron lectin and of its complexes with mono- and disaccharides. J. Mol. Biol. 1998, 277, 917–932. [Google Scholar] [CrossRef]
- Hamelryck, T.W.; Dao-Thi, M.H.; Poortmans, F.; Chrispeels, M.J.; Wyns, L.; Loris, R. The crystallographic structure of phytohemagglutinin-L. J. Biol. Chem. 1996, 271, 20479–20485. [Google Scholar] [CrossRef]
- Nagae, M.; Soga, K.; Morita-Matsumoto, K.; Hanashima, S.; Ikeda, A.; Yamamoto, K.; Yamaguchi, Y. Phytohemagglutinin from Phaseolus vulgaris (PHA-E) displays a novel glycan recognition mode using a common legume lectin fold. Glycobiology 2014, 24, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Pratap, J.V.; Jeyaprakash, A.A.; Rani, P.G.; Sekar, K.; Surolia, A.; Vijayan, M. Crystal structure of artocarpin, a Moraceae lectin with mannose specificity, and its complex with methyl-α-D-mannose: Implications to the generation of carbohydrate specificity. J. Mol. Biol. 2002, 317, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, R.; Sekar, K.; Banerjee, R.; Sharma, V.; Surolia, A.; Vijayan, M. A novel mode of carbohydrate recognition in jacalin, a Moraceae plant lectin with a β-prism fold. Nat. Struct. Biol. 1996, 3, 596–603. [Google Scholar] [CrossRef]
- Lee, X.; Thompson, A.; Zhang, Z.; Ton-that, H.; Biesterfeldt, J.; Ogata, C.; Xu, L.; Johnston, R.A.; Young, N.M. Structure of the complex of Maclura pomifera agglutinin and the T-antigen disaccharide, Galβ1,3GalNAc. J. Biol. Chem. 1998, 273, 6312–6318. [Google Scholar] [CrossRef]
- Rabijns, A.; Barre, A.; Van Damme, E.J.M.; Peumans, W.J.; De Ranter, C.J.; Rougé, P. Structural analysis of the jacalin-related lectin MornigaM from the black mulberry (Morus nigra) in complex with mannose. FEBS J. 2005, 272, 3725–3732. [Google Scholar] [CrossRef] [PubMed]
- Hester, G.; Kaku, H.; Goldstein, I.J.; Wright, C.S. Structure of mannose-specific snowdrop (Galmanthus nivalis) lectin is representative of a new plant lectin family. Nat. Struct. Biol. 1995, 2, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.R.; Meagher, J.L.; Winter, H.C.; Goldstein, I.J.; Paschoalin, V.M.; Silva, J.T.; Stuckey, J.A. High-resolution crystal structures of Colocasia esculenta tarin lectin. Glycobiology 2017, 27, 50–56. [Google Scholar] [CrossRef]
- Transue, T.R.; Smith, A.K.; Mo, H.; Goldstein, I.J.; Saper, M.A. Structure of benzyl T-antigen disaccharide bound to Amaranthus caudatus agglutinin. Nat. Struct. Biol. 1997, 4, 779–783. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, K.; Tsaneva, M.; Kulkarni, S.R.; Rougé, P.; Vandepoele, K.; Van Damme, E.J.M. Evolutionary relationships and expression analysis of EUL domain proteins in rice (Oryza sativa). Rice 2017, 10, 26. [Google Scholar] [CrossRef]
- Sharma, U.; Katre, U.V.; Suresh, C.G. Crystal structure of a plant albumin from Cicer arietinum (chickpea) possessing hemopexin fold and hemagglutination activity. Planta 2015, 241, 1061–1073. [Google Scholar] [CrossRef]
- Kostlánová, N.; Mitchell, E.P.; Lortat-Jacob, H.; Oscarson, S.; Lahmann, M.; Gilboa-Garber, N.; Chambat, G.; Wimmerová, M.; Imberty, A. The fucose-binding lectin from Ralstonia solanacearum. A new type of β-propeller architecture formed by oligomerization and interacting with fucoside, fucosyllactose, and plant xyloglucan. J. Biol. Chem. 2005, 280, 27839–27849. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sýkorová, P.; Demo, G.; Dobeš, P.; Hyršl, P.; Wimmerová, M. A novel fucose-binding lectin from Photorhabdus luminescens (PPL) with an unusual heptabladed β-propeller tetrameric structure. J. Biol. Chem. 2016, 291, 25032–25049. [Google Scholar] [CrossRef] [PubMed]
- Jančařiková, G.; Houser, J.; Dobeš, P.; Demo, G.; Hyršl, P.; Wimmerová, M. Characterization of novel bangle lectin from Photorhabdus asymbiotica with dual sugar-binding specificity and its effect on host immunity. PLoS Pathog. 2017, 13, e1006564. [Google Scholar] [CrossRef] [PubMed]
- Yadid, I.; Kirshenbaum, N.; Sharon, M.; Dym, O.; Tawfik, D.S. Metamorphic proteins mediate evolutionary transitions of structure. Proc. Natl Acad. Sci. USA 2010, 107, 7287–7292. [Google Scholar] [CrossRef] [PubMed]
- Cioci, G.; Mitchell, E.P.; Chazalet, V.; Debray, H.; Oscarson, S.; Lahmann, M.; Gautier, C.; Breton, C.; Perez, S.; Imberty, A. Beta-propeller crystal structure of Psathyrella velutina lectin: An integrin-like fungal protein interacting with monosaccharides and calcium. J. Mol. Biol. 2006, 357, 1575–1591. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.P.; Ali Abol Hassan, M.; Rouf, R.; Tiralongo, E.; May, T.W.; Day, C.J.; Imberty, A.; Tiralongo, J.; Varrot, A. Biophysical characterization and structural determination of the potent cytotoxic Psathyrella asperospora lectin. Proteins 2017, 85, 969–975. [Google Scholar] [CrossRef]
- Audfray, A.; Claudinon, J.; Abounit, S.; Ruvoën-Clouet, N.; Larson, G.; Smith, D.F.; Wimmerová, M.; Le Pendu, J.; Römer, W.; Varrot, A.; et al. Fucose-binding lectin from opportunistic pathogen Burkholderia ambifaria binds to both plant and human oligosaccharidic epitopes. J. Biol. Chem. 2012, 287, 4335–4347. [Google Scholar] [CrossRef]
- Shahzad-ul-Huassan, S.; Gustchina, E.; Ghirlando, R.; Clore, G.M.; Bewley, C.A. Solution structure of the monovalent lectin microvirin in complex with Manα(1-2)Man provides a basis for anti-HIV activity with low toxicity. J. Biol. Chem. 2011, 286, 20788–20796. [Google Scholar] [CrossRef]
- Chiba, H.; Inokoshi, J.; Okamoto, M.; Matsuzaki, K.; Iwama, M.; Mizumoto, K.; Tanaka, H.; Oheda, M.; Fujita, K.; Nakashima, H.; et al. Actinohivin, a novel anti-HIV protein from an actinomycete that inhibits syncytium formation: Isolation, characterization, and biological activites. Biochem. Biophys. Res. Commun. 2001, 282, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Lameignere, E.; Malinovská, L.; Sláviková, M.; Duchaud, E.; Mitchell, E.P.; Varrot, A.; Sedo, O.; Imberty, A.; Wimmerová, M. Structural basis for mannose recognition by a lectin from opportunistic bacteria Burkholderia cenocapacia. Biochem. J. 2008, 411, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Matei, E.; Basu, R.; Furey, W.; Shi, J.; Calnan, C.; Aiken, C.; Gronenborn, A.M. Structure and glycan binding of a new cyanovirin-N homolog. J. Biol. Chem. 2016, 291, 18967–18976. [Google Scholar] [CrossRef] [PubMed]
- Matei, E.; Louis, J.M.; Jee, J.; Gronenborn, A.M. NMR solution structure of a cyanovirin homolog from wheat head blight fungus. Proteins 2011, 79, 1538–1549. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.I.; Furey, W.; Gronenborn, A.M. Novel fold and carbohydrate specificity of the potent anti-HIV cyanobacterial lectin from Oscillatoria agardhii. J. Biol. Chem. 2011, 286, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.C., Jr.; Lee, J.Y.; Cai, M.; Bewley, C.A.; Clore, G.M. Crystal structures of the HIV-1 inhibitory protein MVL free and bound to Man3GlcNAc2: Structural basis for specificity and high-affinity binding to the core pentasaccharide from N-linked oligomannoside. J. Biol. Chem. 2005, 280, 29269–29276. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Kollipara, S.; Aiken, C.; Gronenborn, A.M. Structural insights into the anti-HIV activity of the Oscillatoria agardhii agglutinin homolog lectin family. J. Biol. Chem. 2012, 287, 33796–33811. [Google Scholar] [CrossRef] [PubMed]
- Clore, G.M.; Bewley, C.A. Using conjoined rigid body.torsion angle simulated annealing to determine the relative orientation of covalently linked protein domains from dipolar couplings. J. Magn. Reson. 2002, 154, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.; Garcia-Pino, A.; Lebbe, E.K.; Spaepen, S.; Loris, R.; De Mot, R. Structural determinants for activity and specificity of the bacterial toxin LlpA. PLoS Pathog. 2013, 9, e1003199. [Google Scholar] [CrossRef] [PubMed]
- McFeeters, R.L.; Xiong, C.; O’Keefe, B.R.; Bokesh, H.R.; McMahon, J.B.; Ratner, D.M.; Castelli, R.; Seeberger, P.H.; Byrd, R.A. The novel fold of scytovirin reveals a new twist for antiviral entry inhibitors. J. Mol. Biol. 2007, 369, 451–461. [Google Scholar] [CrossRef]
- Boraston, A.B.; Revett, T.J.; Boraston, C.M.; Nurizzo, D.; Davies, G.J. Structural and thermodynamic dissection of specific mannan recognition by a carbohydrate binding module, TmCBM27. Structure 2003, 665–675. [Google Scholar] [CrossRef]
- Bourne, Y.; Rougé, P.; Cambillau, C. X-ray structure of a (α-Man(1-3)β-Man(1-4)GlcNAc)-lectin complex at 2.1-Å resolution. The role of water in sugar-lectin interaction. J. Biol. Chem. 1990, 265, 18161–18165. [Google Scholar] [PubMed]
- Bourne, Y.; Rougé, P.; Cambillau, C. X-ray structure of a biantennary octasaccharide-lectin complex refined at 2.3-Å resolution. J. Biol. Chem. 1992, 267, 197–203. [Google Scholar] [PubMed]
- Bourne, Y.; Mazurier, J.; Legrans, D.; Rougé, P.; Montreuil, J.; Spik, G.; Cambillau, C. Structure of a legume lectin complexed with the human lactotransferrin N2 fragment, and with an isolated biantennary glycopeptide: Role of the fucose moiety. Structure 1994, 2, 209–219. [Google Scholar] [CrossRef]
- Buts, L.; Garcia-Pino, A.; Wyns, L.; Loris, R. Structural basis of carbohydrate recognition by a Man(α1-2)Man-specific lectin from Bowringia milbraedii. Glycobiology 2006, 16, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Naismith, J.H.; Field, R.A. Structural basis of trimannoside recognition by concanavalin A. J. Biol. Chem. 1996, 271, 972–976. [Google Scholar] [CrossRef] [PubMed]
- Moothoo, D.N.; Naismith, J.H. Concanavalin A distorts the β-GlcNAc-(1,2)-Man linkage of β-GlcNAc-(1,2)-α-Man-(1,3)-[β-GlcNAc-(1,2)-α-Man)(1,6)]-Man upon binding. Glycobiology 1998, 8, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Moothoo, D.N.; Canan, B.; Field, R.A.; Naismith, J.H. Manα1-2Manα-OMe-concanavalin A complex reveals a balance of forces involved in carbohydrate recognition. Glycobiology 1999, 9, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.A.; Moothoo, D.N.; Reftery, J.; Howard, A.J.; Helliwell, J.R.; Naismith, J.H. The 1.2 Å resolution structure of the Con A-dimannose complex. J. Mol. Biol. 2001, 310, 875–884. [Google Scholar] [CrossRef]
- Loris, R.; Maes, D.; Poortmans, F.; Wyns, L.; Bouckaert, J. A structure of the complex between concanavalin A and methyl-3,6-di-O-(α-d-mannopyranosyl)-α-d-mannopyranoside reveals two binding modes. J. Biol. Chem. 1996, 271, 30614–30618. [Google Scholar] [CrossRef]
- Bouckaert, J.; Hamelryck, T.W.; Wyns, L.; Loris, R. The crystal structure of Man(α1-3)Man(α1-O)Me and Man(α1-6)Man(α1-O)Me in complex with concanavalin A. J. Biol. Chem. 1999, 274, 29188–29195. [Google Scholar] [CrossRef]
- Kanellopoulos, P.N.; Pavlou, K.; Perrakis, A.; Agianian, B.; Vorgias, C.E.; Mavrommatis, C.; Soufi, M.; Tucker, P.A.; Hamodrakas, S.J. The crystal structure of the complexes of concanavalin A with 4′-nitrophenyl-α-d-mannopyranoside and 4′-nitrophenyl-α-d-glycopyranoside. J. Struct. Biol. 1996, 116, 345–355. [Google Scholar] [CrossRef]
- Naismith, J.H.; Emmerich, C.; Habash, J.; Harrop, S.J.; Helliwell, J.R.; Hunter, W.N.; Raftery, J.; Kalb, A.J.; Yariv, J. Refined structure on concanavalin A complexed with methyl α-d-mannopyranoside at 2.0 Å resolution and comparison with the saccharide-free structure. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 847–858. [Google Scholar] [CrossRef]
- Gerlits, O.O.; Coates, L.; Woods, R.J.; Kovalevsky, A. Mannobiose binding induces changes in hydrogen bonding and protonation states of acidic residues in concanavalin A as revealed by neutron crystallograohy. Biochemistry 2017, 56, 4747–4750. [Google Scholar] [CrossRef]
- Delatorre, P.; Rocha, B.A.; Souza, E.P.; Oliveira, T.M.; Benzerra, G.A.; Moreno, F.B.; Freitas, B.T.; Santi-Gadelha, T.; Sampaio, A.H.; Azevedo, W.F., Jr.; et al. Structure of a lectin from Canavalia gladiata seeds: New structureinsightys for old molecules. BMC Struct. Biol. 2007, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, G.A.; Oliveira, T.M.; Moreno, F.B.; de Souza, E.P.; da Rocha, B.A.; Benevides, R.G.; Delatorre, P.; de Azevedo, W.F., Jr.; Cavada, B.S. Structural analysis of Canavalia maritima and Canavalia gladiata lectins complexed with different dimannosides: New insights into the understaznding of the structure-biological activity relationship in legume lectins. J. Struct. Biol. 2007, 160, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Roussel, A.; Frey, M.; Rougé, P.; Fontecilla-Camps, J.C.; Cambillau, C. Three-dimensional structures of Lathyrus ochrus isolectin I with glucose and mannose: Fine specificity of the monosaccharide-binding site. Proteins 1990, 8, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Junior, V.R.; Santiago, M.Q.; Nobre, C.B.; Osterne, V.J.S.; Leal, R.B.; Cajazeiras, J.B.; Lossio, C.F.; Rocha, B.A.M.; Martins, M.G.Q.; Nobre, C.A.S.; et al. Crystal structure of Pisum arvense seed lectin (PAL) and characterization of its interaction with carbohydrates by molecular docking and dynamics. Arch. Biochem. Biophys. 2017, 630, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Ruzheinikov, S.N.; Mikhailova, I.Y.; Tsygannik, I.N.; Pangborn, W.; Duax, W.; Pletnev, V.Z. The structure of the pea lectin-D-mannopyanose complex at 2.1 Å resolution. Russ. J. Bioorg. Chem. 1998, 24, 277–279. [Google Scholar]
- Rini, J.M.; Hardman, K.D.; Einspahr, H.; Suddath, F.L.; Carver, J.P. X-ray crystal structure of a pea lectin-trimannoside complex at 2.6 Å resolution. J. Biol. Chem. 1993, 268, 10126–10132. [Google Scholar]
- Loris, R.; Van Walle, I.; De Greve, H.; Beeckmans, S.; Deboeck, F.; Wyns, L.; Bouckaert, J. Structural basis of oligomannose recognition by the Pterocarpus angolensis seed lectin. J. Mol. Biol. 2004, 335, 1227–1240. [Google Scholar] [CrossRef]
- Buts, L.; Garcia-Pino, A.; Imberty, A.; Amiot, N.; Boon, G.J.; Beeckmans, S.; Versées, W.; Wyns, L.; Loris, R. Structural basis for the recognition of complex-type biantennary oligosaccharides by Pterocarpus angolensis lectin. FEBS J. 2006, 273, 2407–2420. [Google Scholar] [CrossRef]
- Garcia-Pino, A.; Buts, L.; Wyns, L.; Loris, R. Interplay between metal binding and cis/trans isomerization in legume lectins: Structural and thermodynamic study of P. angolensis lectin. J. Mol. Biol. 2006, 361, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pino, A.; Buts, L.; Wyns, L.; Imberty, A.; Loris, R. How a plant lectin recognizes high mannose oligosaccharides. Plant Physiol. 2007, 144, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Jeyaprakash, A.A.; Srivastav, A.; Surolia, A.; Vijayan, M. Structural basis for the carbohydrate specificities of artocarpin: Variation in the length of a loop as a strategy for generating ligand specificity. J. Mol. Biol. 2004, 338, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Chandra, N.R.; Ramachandraiah, G.; Bachhawat, K.; Dam, T.K.; Surolia, A.; Vijayan, M. Crystal structure of a dimeric mannose-specific agglutinin from garlic: Quaternary association and carbohydrate specificity. J. Mol. Biol. 1999, 285, 1157–1168. [Google Scholar] [CrossRef]
- Ramachandraiah, G.; Chandra, N.R.; Surolia, A.; Vijayan, M. Re-refinement using processed data to improve the quality of the structure: A case study involving garlic lerctin. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.S.; Hester, G. The 2.0 Å structure of a cross-linked complex between snowdrop lectin and a branched mannopentaose: Evidence for two unique binding modes. Structure 1996, 4, 1339–1352. [Google Scholar] [CrossRef]
- Hester, G.; Wright, C.S. The mannose-specific bulb lectin from Galanthus nivalis (snowdrop) binds mono- and dimannosides at distinct sites. Structure analysis of refined complexes at 2.3 Å and 3.0 Å resolution. J. Mol. Biol. 1996, 262, 516–531. [Google Scholar] [CrossRef] [PubMed]
- Sauerborn, M.K.; Wright, L.M.; Reynolds, C.D.; Grossmann, J.G.; Rizkallah, P.J. Insights into carbohydrate recognition by Narcissus pseudonarcissus lectin: The crystal structure at 2 Å resolution in complex with α1-3 mannobiose. J. Mol. Biol. 1999, 290, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.D.; Boudreaux, D.M.; Salmon, L.; Chugh, J.; Winter, H.C.; Meagher, J.L.; Andre, S.; Murphy, P.V.; Oscarson, S.; Roy, R.; et al. Engineering a therapeutic lectin by uncoupling mitogenicity from antiviral activity. Cell 2015, 163, 746–758. [Google Scholar] [CrossRef]
- Nagae, M.; Mishra, S.K.; Hanashima, S.; Tateno, H.; Yamaguchi, Y. Distinct roles for each N-glycan branch interacting with mannose-binding type jacalin-related lectins Orysata and Calsepa. Glycobiology 2017, 27, 1120–1133. [Google Scholar] [CrossRef]
- Zlólkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; McMahon, J.B.; Palmer, K.E.; Dwek, R.A.; Wormald, M.R.; Wlodawer, A. Crystallographic, thermodynamic, and molecular modeling studies of the mode of binding of oligosaccharides to the potent antiviral protein griffthsion. Proteins 2007, 67, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Moulaei, T.; Shenoy, S.R.; Giomarelli, B.; Thomas, C.; McMahon, J.B.; Dauter, Z.; O’Keefe, B.R.; Wlodawer, A. Monomerization of viral entry inhibitor griffithsin elucidates the relationship between multivalent binding to carbohydrates and anti-HIV activity. Structure 2010, 18, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Hoque, M.M.; Jiang, J.; Suzuki, K.; Tsunoda, M.; Takeda, Y.; Ito, Y.; Kawai, G.; Tanaka, H.; Takenaka, A. The characteristic structure of anti-HIV actinohivin in complex with three HMTG D1 chains of HIV-gp120. Chembiochem. 2014, 15, 2766–2773. [Google Scholar] [CrossRef] [PubMed]
- Debray, H.; Decout, D.; Strecker, G.; Spik, G.; Montreuil, J. Specificity of twelve lectins towards oligosaccharides and glycopeptides related to N-glycosylproteins. Eur. J. Biochem. 1981, 117, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Fouquaert, E.; Van Damme, E.J.M. Promiscuity of the Euonymus carbohydrate-binding domain. Biomecules 2012, 2, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Gronenborn, A.M. Structural basis of the anti-HIV activity of the cyanobacterial Oscillatoria agardhii agglutinin. Structure 2011, 19, 1170–1181. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Smith, D.; Cummings, R.; Peumans, W.J. Glycan arrays to decipher the specificity of plant lectins. Adv. Exp. Med. Biol. 2011, 705, 757–767. [Google Scholar]
- Hirabayashi, J.; Arata, Y.; Kasai, K. Reinforcement of frontal affinity chromatography for effective analysis of lectin-oligosaccharide interactions. J. Chromatogr. A 2000, 890, 261–271. [Google Scholar] [CrossRef]
- Peumans, W.J.; Van Damme, E.J.M. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Fouquaert, E.; Lannoo, N.; Vanderborre, G.; Schouppe, D.; Peumans, W.J. Novel concepts about the role of lectins in the plant cell. Adv. Exp. Med. Biol. 2011, 705, 271–294. [Google Scholar]
- Sprawka, I.; Golawska, S.; Parzych, T.; Golawski, A.; Czerniewicz, P.; Sytykiewicz, H. Mechanism of entomotoxicity of the concanavalin A in Rhopalosiphum padi (Hemiptera: Aphididae). J. Insect Sci. 2014, 14, 232. [Google Scholar] [CrossRef] [PubMed]
- Majumder, P.; Banerjee, S.; Das, S. Identification of receptors responsible for binding of the mannose specific lectin to the gut epithelial membrane of the target insects. Glycoconj. J. 2004, 20, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Majumder, P.; Mondal, H.A.; Das, S. Insecticidal activity of Arum maculatum tuber lectin and its binding to the glycosylated insect gut receptors. J. Agric. Food Chem. 2005, 53, 6725–6729. [Google Scholar] [CrossRef] [PubMed]
- Cristofoletti, P.T.; de Sousa, F.A.; Rahbé, Y.; Terra, W.R. Characterization of a membrane-bound aminopeptidase purified from Acyrthosiphon pisum midgut cells. A major binding site for toxic mannose lectins. FEBS Lett. 2006, 273, 5574–5588. [Google Scholar] [CrossRef] [PubMed]
- Fitches, E.; Wiles, D.; Douglas, A.E.; Hinchliffe, G.; Audsley, N.; Gatehouse, J.A. The insecticidal activity of recombinant garlic lectins towards aphids. Insect Biochem. Mol. Biol. 2008, 38, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Sengupta, S.; Roy, A.; Ghosh, P.; Das, K.; Das, S. Functional anteration of a dimeric insecticvidal lectin to a monomeric antifungal protein correlated to its oligomeric status. PLoS ONE 2011, 6, e18593. [Google Scholar] [CrossRef]
- Roy, A.; Gupta, S.; Hess, D.; Das, K.P.; Das, S. Binding of insecticidal lectin Colocasia esculenta tuber agglutinin (CEA) to midgut receptors of Bemisia tabaci and Lipaphis erysimi provides clues to its insecticidal potential. Proteomics 2014, 14, 1646–1659. [Google Scholar] [CrossRef]
- Down, R.E.; Gatehouse, A.M.R.; Hamilton, W.D.O.; Gatehouse, J.A. Snowdrop lectin inhibits development and decreases fecundity of the glasshouse potato aphid (Aulacorthum solani) when administered in vitro and via transgenic plants both in laboratory and glasshouse trials. J. Insect Physiol. 1996, 42, 1035–1045. [Google Scholar] [CrossRef]
- Zhou, Y.; Tian, Y.; Wu, B.; Mang, K. Inhibition effect of transgenic tobacco plants expressing snowdrop lectin on the population development of Myzus persicae. Chin. J. Biotechnol. 1998, 14, 9–16. [Google Scholar]
- Down, R.E.; Ford, L.; Woodhouse, S.D.; Davison, G.M.; Majerus, M.E.; Gatehouse, J.A.; Gatehouse, A.M. Tritrophic interactions between transgenic potato expressing snowdrop lectin (GNA), an aphid pest (peach-potato aphid; Myzus persicae (Sulz.) and a beneficial predator (2-spot ladybird; Adalia bipunctata L.). Transgenic Res. 2003, 12, 229–241. [Google Scholar] [CrossRef]
- Luo, S.; Zhangsun, D.; Tang, K. Functional GNA expressed in Escherichia coli with high efficiency and its effect on Ceratovacuna lanigera Zehntner. Appl. Microbiol. Biotechnol. 2005, 69, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Down, R.E.; Fitches, E.C.; Wiles, D.P.; Corti, P.; Bell, H.A.; Gatehouse, J.A.; Edwards, J.P. Insecticidal spider venom toxin fused to snowdrop lectin is toxic to the peach-potato aphid, Myzus persicae (Hemiptera: Aphidae) and the rice brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Pest Manag. Sci. 2006, 62, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, K.; Sun, X.; Tang, K.; Zhang, J. Enhancement of resistance to aphids by introducing the snowdrop lectin gene GNA into maize plants. J. Biosci. 2005, 30, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Hogervorst, P.A.; Ferry, N.; Gatehouse, A.M.; Wäckers, F.L.; Romeis, J. Direct effects of snowdrop lectin (GNA) on larvae of three aphid predators and fate of GNA after ingestion. J. Insect Physiol. 2006, 52, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Wu, Y.; Xu, W.; Hu, L.; Yu, Z.; Xu, Q. The impact of transgenic wheat expressing GNA (snowdrop lectin) on the aphids Sitobion avenae, Schizaphis graminum, and Rhopalosiphum padi. Environ. Entomol. 2011, 40, 743–748. [Google Scholar] [CrossRef]
- Qi, G.; Lan, N.; Ma, X.; Yu, Z.; Zhao, X. Controlling Myzus persicae with recombinant endophytic fungi Chaetomium globosum expressing Pinellia ternata agglutinin: Using recombinant endophytic fungi to control aphids. J. Appl. Microbiol. 2011, 110, 1314–1322. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, K.; Ding, R.; Zhang, H.; Di, P.; Chen, J.; Zhang, L.; Chen, W. Transgenic tetraploid Isatis indigotica expressing Bt Cry1Ac and Pinellia ternata agglutinin showed enhanced resistance to moths and aphids. Mol. Biol. Rep. 2012, 39, 485–491. [Google Scholar] [CrossRef]
- Duan, X.; Hou, Q.; Liu, G.; Pang, X.; Niu, Z.; Wang, X.; Zhang, Y.; Li, B.; Liang, R. Expression of Pinellia pedatisecta lectin gene in transgenic wheat enhances resistance to wheat aphids. Molecules 2018, 23, 748. [Google Scholar] [CrossRef]
- Javaid, S.; Amin, I.; Jander, G.; Mukhtar, Z.; Saeed, N.A.; Mansoor, S. A transgenic approach to control hemipteran insects by expressing insecticidal genes under phloem-specific promoters. Sci. Rep. 2016, 6, 34706. [Google Scholar] [CrossRef]
- Chakraborti, D.; Sarkar, A.; Mondal, H.A.; Schuermann, D.; Hohn, B.; Sarmah, B.K.; Das, S. Cre/lox system to develop selectable marker free transgenic tobacco plants conferring resistance against sap sucking homopteran insect. Plant Cell Rep. 2008, 27, 1623–1633. [Google Scholar] [CrossRef]
- Chakraborti, D.; Sarkar, A.; Mondal, H.A.; Das, S. Tissue expression of potent insecticidal, Allium sativum leaf agglutinin (ASAL) in important pulse crop, chickpea (Cicer arietinum L.) to resist the phloem feeding Aphis craccivora. Transgenic Res. 2009, 18, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Broeders, S.; De Greve, H.; Hernalsteens, J.P.; Peumans, W.J.; Van Damme, E.J.M.; Smagghe, G. Expression of garlic leaf lectin under the control of the phloem-specific Asus1 from Arabidopsis thaliana protects tobacco plants against the tobacco aphid (Myzus nicotianae). Pest Manag. Sci. 2007, 63, 1215–1223. [Google Scholar] [CrossRef]
- Yao, J.; Zhao, X.; Qi, H.; Wan, B.; Chen, F.; Sun, X.; Yu, S.; Tang, K.X. Transgenic tobacco expressing an Arisaema heterophyllum agglutinin gene displays enhanced resistance to aphids. Can. J. Plant Sci. 2004, 84, 785–790. [Google Scholar]
- Kai, G.; Ji, Q.; Lu, Y.; Qian, Z.; Cui, L. Expression of Monstera deliciosa agglutinin (MDA) in tobacco confers resistance to peach-potato aphids. Integr. Biol. 2012, 4, 937–944. [Google Scholar] [CrossRef]
- Al Atalah, B.; Smagghe, G.; Van Damme, E.J.M. Orysata, a jacalin-related lectin from rice, could protect plants against biting-chewing and piercing-sucking insects. Plant Sci. 2014, 221–222, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.H.; Chen, S.; Zhang, F.; Wang, W.; Tian, Q.; Liu, J.Z.; Chen, F.; Bao, J.K. Transgenic tobacco expressing Zephyranthes grandiflora agglutinin confers enhanced resistance to aphids. Appl. Biochem. Biotechnol. 2009, 158, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Chen, L.; Chen, S.; Cai, H.; Liu, X.; Xiao, G.; Zhu, Z. Transformation of tobacco with genes encoding Helianthus tuberosus agglutinin (HTA) confers resistance to peach-potato aphid (Myzus persicae). Transgenic Res. 2003, 12, 607–614. [Google Scholar] [CrossRef]
- Sauvion, N.; Nardon, C.; Febvay, G.; Gatehouse, A.M.; Rahbé, Y. Binding of the insecticidal lectin concanavalin A in pea aphid, Acyrthosiphon pisum (Harris) and induced effects on the structure of midgut epithelial cells. J. Insect Physiol. 2004, 50, 1137–1150. [Google Scholar] [CrossRef]
- Shahidi-Noghabi, S.; Van Damme, E.J.; Mahdian, K.; Smagghe, G. Entomotoxic action of Sambucus nigra agglutinin I in Acyrthosiphon pisum aphids and Spodoptera exigua caterpillars through caspase-3-like-dependent apoptosis. Arch. Insect. Biochem. Physiol. 2010, 75, 207–220. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Barre, A.; Rougé, P.; Peumans, W.J. Cytoplasmic/nuclear plant lectins: A new story. Trends Plant Sci. 2004, 9, 484–489. [Google Scholar] [CrossRef]
- Lannoo, N.; Van Damme, E.J.M. Nucleocytoplasmic plant lectins. Biochim. Biophys. Acta 2010, 1800, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Delporte, A.; Van Holle, S.; Lannoo, N.; Van Damme, E.J.M. The tobacco lectin, prototype of the family of Nictaba-related proteins. Curr. Protein Pept. Sci. 2015, 16, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Van Holle, S.; Rougé, P.; Van Damme, E.J.M. Evolution and structural diversification of Nictaba-like genes in food crops with focus on soybean (Glycine max). Ann. Bot. 2017, 119, 901–914. [Google Scholar] [PubMed]
- Fouquaert, E.; Peumans, W.J.; Vandekerckhove, T.T.M.; Ongenaert, M.; Van Damme, E.J.M. Proteins with an Euonymus-like domain are ubiquitous in embryophyta. BMC Plant Biol. 2009, 9, 136. [Google Scholar] [CrossRef]
- Esch, L.; Schaffrath, U. An update on jacalin-like lectins and their role in plant defense. Int. J. Mol. Sci. 2017, 18, 1592. [Google Scholar] [CrossRef] [PubMed]
- Lannoo, N.; Peumans, W.J.; Pamel, E.V.; Alvarez, R.; Xiong, T.C.; Hause, G.; Mazars, C.; Van Damme, E.J.M. Localization and in vitro binding studies suggest that the cytoplasmic/nuclear tobacco lectin can interact in situ with high mannose and complex N-glycans. FEBS Lett. 2006, 580, 6329–6337. [Google Scholar] [CrossRef] [PubMed]
- Al Atalah, B.; Fouquaert, E.; Vanderschaeghe, D.; Proost, P.; Balzarini, J.; Smith, D.F.; Rougé, P.; Lasanajak, Y.; Callewaert, N.; Van Damme, E.J.M. Expression analysis of the nucleocytoplasmic lectin ‘Orysata’ from rice in Pichia pastoris. FEBS J. 2011, 278, 2064–2079. [Google Scholar] [CrossRef] [PubMed]
- Al Atalah, B.; Vanderschaeghe, D.; Bloch, Y.; Proost, P.; Plas, K.; Callewaert, N.; Savvides, S.N.; Van Damme, E.J.M. Characterization of a type D1A EUL-related lectin from rice expressed in Pichia pastoris. Biol. Chem. 2014, 395, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Van Holle, S.; Van Damme, E.J.M. Signaling through plant lectins: Modulation of plant immunity and beyond. Biochem. Soc. Trans. 2018, 46, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Van Hove, J.; De Jaeger, G.; De Winne, N.; Guisez, Y.; Van Damme, E.J.M. The Arabidopsis lectin EULS3 is involved in stomatal closure. Plant Sci. 2015, 238, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Van Holle, S.; Smagghe, G.; Van Damme, E.J.M. Overexpression of Nictaba-like lectin genes from Glycine max confers tolerance toward Pseudomonas syringae infection, aphid infestation and salt stress in transgenic Arabidopsis plants. Front. Plant Sci. 2016, 7, 1590. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, L.; Stefanowicz, K.; Van Damme, E.J.M. Nictaba homologs from Arabidopsis thaliana are involved in plant stress responses. Front. Plant Sci. 2018, 8, 2218. [Google Scholar] [CrossRef] [PubMed]
- Stewart-Jones, G.B.; Soto, C.; Lemmin, T.; Chuang, G.Y.; Druz, A.; Kong, R.; Thomas, P.V.; Wagh, K.; Zhou, T.; Behrens, A.J.; et al. Trimeric HIV-en structures define glycan shields from clades A, B, and G. Cell 2016, 165, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Lifson, J.; Coutré, S.; Huang, E.; Engleman, E. Role of evelope glycoprotein carbohydrate in human immunodeficiency virus (HIV) infectivity and virus-induced cell fusion. J. Exp. Med. 1986, 164, 2101–2106. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.E., Jr.; Montefiori, D.C.; Mitchell, W.M. Evidence that mannosyl residues are involved in human immunodeficiency virus type 1 (HIV-1) pathogenesis. AIDS Res. Hum. Retrovir. 1987, 3, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.; Renneisen, K.; Kreuter, M.H.; Schröder, H.C.; Winkler, I. The D-mannose-specific lectin from Gerardia savaglia blocks binding to human immunodeficiency virus type I to H9 cells and human lymphocytes in vitro. J. Acquir. Immune Defic. Syndr. 1988, 1, 453–458. [Google Scholar] [PubMed]
- Hansen, J.E.; Nielsen, C.M.; Nielsen, C.; Heegaard, P.; Mathiesen, L.R.; Nielsen, J.O. Correlation between carbohydrates structures on the envelope glycoprotein gp120 of HIV-1 and HIV-2 and syncytium inhibition with lectins. AIDS 1989, 3, 635–641. [Google Scholar] [CrossRef]
- Hammar, L.; Eriksson, S.; Morein, B. Human immunodeficiency virus glycoproteins: Lectin binding properties. AIDS Res. Hum. Retrovir. 1989, 5, 495–506. [Google Scholar] [CrossRef]
- Houlès Astoul, C.; Peumans, W.J.; Van Damme, E.J.M.; Rougé, P. Accessibility of the high-mannose glycans of glycoprotein gp120 from human immunodeficiency virus type 1 probed by in vitro interaction with mannose-binding lectins. Biochem. Biophys. Res. Commun. 2000, 274, 455–460. [Google Scholar] [CrossRef]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Cai, M.; Ray, S.; Ghirlando, R.; Yamaguchi, M.; Muramoto, K. New carbohydrate specificity and HIV-1 fusion bloking activity of the cyanobacterial protein MVL: NMR, ITC and sedimentation equilibrium studies. J. Mol. Biol. 2004, 339, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Férirn, G.; Huskens, D.; Noppen, S.; Koharudin, L.M.; Gronenborn, A.M.; Schols, D. Broad anti-HIV activity of the Oscillatoria agardhii agglutinin homologue lectin family. J. Antimicrob. Chemother. 2014, 69, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.; Gardella, R.S.; Sowder, R.C., 2nd; Turpin, J.; Watson, K.; Buckheit, R.W., Jr.; et al. A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef]
- Robinson, J.E.; Holton, D.; Liu, J.; McMurdo, H.; Murciano, A.; Gohd, R. A novel enzyme-linked immunosorbent assay (ELISA) for the detection of antibodies to HIV-1 envelope glycoproteins based on immobilization of viral glycoproteins in microtiter wells coated with concanavalin A. J. Immunol. Methods 1990, 132, 63–71. [Google Scholar] [CrossRef]
- Gattegno, L.; Ramdani, A.; Jouault, T.; Saffar, L.; Gluckman, J.C. Lectin-carbohydrate interactions and infectivity of human immunodeficiency virus type 1 (HIV-1). AIDS Res. Hum. Retrovir. 1992, 8, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.K.; Klinkner, A.M.; Ventre, J.; Bugelski, P.J. Morphometric analysis of envelope glycoprotein gp120 distribution on HIV-1 virions. J. Histochem. Cytochem. 1993, 41, 265–271. [Google Scholar] [CrossRef]
- Pal, R.; DeVico, A.; Rittenhouse, S.; Sarngadharan, M.G. Conformational perturbation of the envelopr glycoprotein gp120 of human immubodeficiency virus type 1 by soluble CD4 and the lectin succinyl Con A. Virology 1993, 194, 833–837. [Google Scholar] [CrossRef]
- Yeh, J.C.; Seals, J.R.; Murphy, C.I.; van Halbeek, H.; Cummings, R.D. Site-specific N-glycosylation and oligosaccharide structures of recombinant HIV-1 gp120 derived from a baculovirus expression system. Biochemistry 1993, 32, 11087–11099. [Google Scholar] [CrossRef]
- Gram, G.J.; Hemming, A.; Bolmstedt, A.; Jansson, B.; Olofsson, S.; Akerblom, L.; Nielsen, J.O.; Hansen, J.E. Identification of an N-linked glycan in the V1-loop of HIV-1 gp120 influencing neutralization by anti-V3 antibodies and soluble CD4. Arch. Virol. 1994, 139, 253–261. [Google Scholar] [CrossRef]
- Gu, J.J.; Harriss, J.V.; Ozato, K.; Gottlieb, P.D. Induction by concanavalin A of specific mRNAs and cytolytic function in a CD8-positive T cell hybridoma. J. Immunol. 1994, 153, 4408–4417. [Google Scholar] [PubMed]
- Akashi, M.; Niikawa, T.; Serizawa, T.; Hayakawa, T.; Baba, M. Capture of HIV-1 gp120 and virions by lectin-immobilized polystyrene nanospheres. Bioconj. Chem. 1998, 9, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Kawamura, M.; Okamoto, M.; Baba, M.; Niikawa, T.; Takehara, S.; Serizawa, T.; Akashi, M. Concanavalin A-immobilized polystyrene nanospheres capture HIV-1 vririons and gp120: Potential approach towards prevention of viral transmission. J. Med. Virol. 1998, 56, 327–331. [Google Scholar] [CrossRef]
- No, T.B.; Chan, Y.S.; Ng, C.C.; Wong, J.H. Purification and characterization of a lectin from green split peas (Pisum sativum). Appl. Biochem. Biotechnol. 2015, 177, 1374–1385. [Google Scholar]
- Balzarini, J.; Neyts, J.; Schols, D.; Hosoya, M.; Van Damme, E.; Peumans, W.; De Clercq, E. The mannose-specific lectins from Cymbidium hybrid and Epipactis helleborine and the (N-acetylglucosamine)n-specific plant lectin from Urtica dioica are potent and selective inhibitors of human immunodeficiency virus and cytomegalovirus replication in vitro. Antivir. Res. 1992, 18, 191–207. [Google Scholar] [PubMed]
- Balzarini, J.; Schols, D.; Neyts, J.; Van Damme, E.; Peumans, W.J.; De Clercq, E. α-(1-3)- and α-(1-6)-d-mannose-specific plant lectins are markedly inhibitory to human immunodeficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother. 1991, 35, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, N.; Hay, A.J. An ELISA utilizing immobilized snowdrop lectin GNA for the detection of envelope glycoproteins of HIV and SIV. J. Immunol. Methods 1992, 151, 9–13. [Google Scholar] [CrossRef]
- Gilljam, G. Envelope glycoproteins of HIV-1, HIV-2, and SIV purified with Galanthus nivalis agglutinin induce strong immune responses. AIDS Res. Hum. Retrovir. 1993, 9, 431–438. [Google Scholar] [CrossRef]
- Hammar, L.; Hirsch, I.; Machado, A.; De Mareuil, J.; Baillon, J.; Chermann, J.C. Lectin effects on HIV-1 infectivity. Ann. N. Y. Acad. Sci. 1994, 724, 166–169. [Google Scholar] [CrossRef]
- Hammar, L.; Hirsch, I.; Machado, A.A.; De Mareuil, J.; Baillon, J.G.; Bolmont, C.; Chermann, J.C. Lectin-mediated effects on HIV type 1 infection in vitro. AIDS Res. Hum. Retroviruses 1995, 11, 87–95. [Google Scholar] [CrossRef]
- Balzarini, J.; Hatse, S.; Vermeire, K.; Princen, K.; Aquaro, S.; Perno, C.-F.; De Clercq, E.; Egberink, H.; Vanden Mooter, G.; Peumans, W.; et al. Mannose-specific plant lectins from the amaryllidaceae family qualify as efficient microbicides for prevention of human immunodeficiency virus infection. Antimicrob. Agents Chemother. 2004, 48, 3858–3870. [Google Scholar] [CrossRef] [PubMed]
- Weiler, B.E.; Schröder, H.C.; Stefanovich, V.; Stewart, D.; Forrest, J.M.; Allen, L.B.; Bowden, B.J.; Kreuter, M.H.; Voth, R.; Müller, W.E. Sulphoevernan, a polyanionic polysaccharide, and narcissus lectin potently inhibit human immunodeficiency virus infection by binding to viral envelope protein. J. Gen. Virol. 1990, 71, 1957–1963. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zheng, B.; Wang, W.; Li, Q.; Yuan, Q. Cloning and functional characterization of a GNA-like lectin from Chinese narcissus (Narcissus tazetta var. Chinensis Roem). Physiol. Plant. 2011, 142, 193–204. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Liu, J.Z.; Wu, C.F.; Li, J.; Dai, L.; Van Damme, E.; Balzarini, J.; De Clercq, E.; Chen, F.; Bao, J.K. Anti-HIV I/II activity and molecular cloning of a novel mannose/sialic acid-binding lectin from rhizome of Polygonatum cyrtonema Hua. Acta Biochim. Biophys. Sin. (Shanghai) 2006, 38, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.D.; Winter, H.C.; Goldstein, I.J.; Markovitz, D.M. A lection isolated from bananas is a potent inhibitor of HIV replication. J. Biol. Chem. 2010, 285, 8646–8655. [Google Scholar] [CrossRef] [PubMed]
- Hoorelbeke, B.; Van Damme, E.J.; Rougé, P.; Schols, D.; Van Laethem, K.; Fouquaert, E.; Balzarini, J. Differences in the mannose oligomer specificities of the closely related lectins from Galanthus nivalis and Zea mays strongly determine their eventual anti-HIV activity. Retrovirology 2011, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jin, W.; Griffin, G.E.; Shattock, R.J.; Hu, Q. Removal of two high-mannose N-linked glycans on gp120 renders human immunodeficiency virus 1 largely resistant to the carbohydrate-binding agent griffithsin. J. Gen. Virol. 2011, 92, 2367–2373. [Google Scholar] [CrossRef]
- Hirahyama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-mannose specific lectin and its recombinants from a carrageenophyta Kappaphycus alvarezii represent a potent anti-HIV activity through high-affinity binding to the viral envelope glycoprotein gp120. Mar. Biotechnol. 2016, 18, 215–231. [Google Scholar] [CrossRef]
- Witvrouw, M.; Fikkert, V.; Hantson, A.; Pannecouque, C.; O’Keefe, B.R.; McMahon, J.; Stamatatos, L.; de Clercq, E.; Bolmstedt, A. Resistance of human immunodeficiency virus type 1 to the high-mannose binding agents cyanovirin N and concanavalin A. J. Virol. 2005, 79, 7777–7784. [Google Scholar] [CrossRef]
- Balzarini, J.; Van Laethem, K.; Peumans, W.J.; Van Damme, E.J.; Bolmstedt, A.; Gago, F.; Schols, D. Mutational pathways, resistance profile, and side effects of cyanovirin relative to human immunodefiociency virus type 1 strains with N-glycan deletions in their gp120 envelopes. J. Virol. 2006, 80, 8411–8421. [Google Scholar] [CrossRef]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Vermeire, K.; De Clercq, E.; Peumans, W.; Van Damme, E.; Vandamme, A.-M.; Böhlmstedt, A.; Schols, D. Profile of resistance of human ommunodeficiency virus to mannose-specific plant lectins. J. Virol. 2004, 78, 10617–10627. [Google Scholar] [CrossRef] [PubMed]
- Pancera, M.; Zhou, T.; Druz, A.; Georgiev, I.S.; Soto, C.; Gorman, J.; Huang, J.; Acharya, P.; Chuang, G.-Y.; Ofek, G.; et al. Structure and immune recognition of trimeric prefusion HIV1 Env. Nature 2014, 514, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Froeyen, M.; Peumans, W.; Van Damme, E.; Schols, D. Carbohydrate-binding agents cause deletions of highly conserved glycosylation sites in HIV gp120: A new therapeutic concept to hit the Achilles heel of HIV. J. Biol. Chem. 2005, 280, 41005–41014. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Froeyen, M.; Van Damme, E.; Bolmstedt, A.; Peumans, W.; De Clercq, E.; Scols, D. Marked depletion of glycosylation sites of HIV-1 gp120 under selective pressure by the mannose-specific plant lectins of Hippeastrum hybrid. and Galanthus nivalis. Mol. Pharemacol. 2005, 67, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J. Carbohydrate-binding agents: A potential future cornerstone for the chemotherapy of enveloped viruses? Antivir. Chem. Chemother. 2007, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J. The α(1,2)-mannosidase I inhibitor 1-deoxymannojirimycin potentiates the antiviral activity of carbohydrate-binding agents against wild-type and mutant HIV-1 strains containing glycan deletions in gp120. FEBS Lett. 2007, 581, 2060–2064. [Google Scholar] [CrossRef] [PubMed]
- Férir, G.; Huskens, D.; Palmer, K.E.; Boudreaux, D.M.; Swanson, M.D.; Markovitz, D.M.; Balzarini, J.; Schols, D. Combinations of griffithsin with other carbohydrate-binding agents demonstrate superior activity against HIV type 1, HIV type 2, and selected carbohydrate-ding agent-resistant HIV type 1 strains. AIDS Res. Hum. Retrovir. 2012, 28, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.W.; Ng, T.B. A monomeric mannose-binding lectin from inner shhots of the edible chive (Allium tuberosum). J. Protein Chem. 2001, 20, 361–366. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B. Examination of lectins, polysaccharopeptide, polysaccharide, alkaloid, coumarin and trypsin inhibitors for inhibitory activity against human immunodeficiency virus reverse transcriptase and glycohydrolases. Planta Med. 2001, 67, 669–672. [Google Scholar] [CrossRef]
- Pollicita, M.; Schols, D.; Aquaro, S.; Peumans, W.J.; Van Damme, E.J.; Perno, C.F.; Balzarini, J. Carbohydrate-binding agents (CBAs) inhibit HIV-1 infection in human primary monocyte-derived macrophages (MDMs) and efficienctly prevent MDM-directed viral capture and subsequent trabsmission to CD4+ T lymphocytes. Virology 2008, 370, 382–391. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Gray, E.S.; Mufhandu, H.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. The lectins griffithsin, cyanovirin-N and scytovirin inhibit HIV-1 binding to the DC-SIGN receptor and transfer to CD4(+) cells. Virology 2012, 423, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Hoorelbeke, B.; Xue, J.; LiWang, P.J.; Balzarini, J. Role of the carbohydrate-binding sites of griffithsin in the prevention of DC-SIGN-mediated capture and transmission of HIV-1. PLoS ONE 2013, 8, e64132. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antivir. Res. 2017, 142, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Lafont, V.; Dornand, J.; Covassin, L.; Liautard, J.P.; Favero, J. The lectin jacalin triggers CD4-mediated lymphocyte signaling by binding CD4 through a protein-protein interaction. J. Leukoc. Biol. 1996, 59, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Gordts, S.C.; Renders, M.; Férir, G.; Huskens, D.; Van Damme, E.J.; Peumans, W.; Balzarini, J.; Scols, D. NICTABA and UDA, two GlcNAc-binding lectins with unique antiviral activity profiles. J. Antimicrob. Chemother. 2015, 70, 1674–1685. [Google Scholar] [CrossRef]
- Poiroux, G.; Barre, A.; Van Damme, E.J.M.; Benoist, H.; Rougé, P. Plant lectins targeting O-glycans at the cell surface as tools for cancer diagnosis, prognosis and therapy. Int. J. Mol. Sci. 2017, 18, 1232. [Google Scholar] [CrossRef] [PubMed]
- Li, W.W.; Yu, J.Y.; Xu, H.L.; Bao, J.K. Concanavalin A: A potential anti-neoplastic agent targeting apoptosis, autophagy and anti-angiogenesis for cancer therapeutics. Biochem. Biophys. Res. Commun. 2011, 414, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Bao, J.K. Anti-tumor and anti-viral activities of Galanthus nivalis agglutinin (GNA)-related lectins. Glycoconj. J. 2013, 30, 269–279. [Google Scholar] [CrossRef]
- Jiang, Q.L.; Zhang, S.; Tian, M.; Zhang, S.Y.; Xie, T.; Chen, D.Y.; Chen, Y.J.; He, J.; Liu, J.; Ouyang, L.; et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 2015, 48, 17–28. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Lam, A.K.; Kabir, S.R. Pea lectin inhibits cell growth by inducing apoptosis in SW480 and SW48 cell lines. Int. J. Biol. Macromol. 2018, 117, 1050–1057. [Google Scholar] [CrossRef]
- Kabir, S.R.; Nabi, M.M.; Hague, A.; Uz Zaman, R.; Mahmud, Z.H.; Reaza, M.A. Pea lectin inhibits growth of Ehrlich ascites carcinoma cells by inducing apoptosis and G2/M cell cycle arrest in vivo in mice. Phytomedicine 2013, 20, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Asaduzzaman, A.K.M.; Hasan, I.; Chakrabortty, A.; Zaman, S.; Islam, S.S.; Ahmed, F.R.S.; Kabir, K.M.A.; Nurujjaman, M.; Uddin, M.B.; Alam, M.T.; et al. Moringa oleifera seed lectin inhibits Ehrlich ascites carcinoma cell growth by inducing apoptosis through the regulation of Bak and NF-kB gene expression. Int. J. Biol. Macromol. 2018, 107, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Luz, L.; Rossato, F.A.; Costa, R.A.P.E.; Napoleão, T.H.; Paiva, P.M.G.; Coelho, L.C.B.B. Cytotoxicity of the coagulant Moringa oleifera lectin (cMOL) to B16-F10 melanoma cells. Toxicol In Vitro 2017, 44, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Gondim, A.C.S.; Romero-Canelón, I.; Sousa, E.H.S.; Blindauer, C.A.; Butler, J.S.; Romero, M.J.; Sanchez-Cano, C.; Sousa, B.L.; Chaves, R.P.; Nagano, C.S.; et al. The potent anti-cancer activity of Dioclea lasiocarpa lectin. J. Inorg. Biochem. 2017, 175, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Luan, V.V.; Qian, Y.; Ni, X.; Chandra, T.K.; Xia, Y.; Wang, J.; Yan, Y.; Xu, B. Polygonatum odoratum lectin promotes BECN1 expression and induces autophagy in malignant melanoma by regulation of miR1290. OncoTargets Ther. 2017, 10, 4569–4577. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, T.; Xiao, Y.; Li, X.; Zhu, Y.; Zhao, Y.; Bao, J.; Wu, C. Polygonatum odoratum lectin induces apoptosis and autophagy by regulation of microRNA-1290 and microRNA-15a-3p in human lung adenocarcinoma A549 cells. Int. J. Biol. Macromol. 2016, 85, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Chen, Y.; Wang, X.Y.; Lu, R.F.; Zhang, S.Y.; Tian, M.; Xie, T.; Liu, B.; He, G. Polygonatum odoratum lectin induces apoptosis and autophagy via targeting EGFR-mediated Ras-Raf-MEK-ERK pathway in human MCF-7 breast cancer cells. Phytomedicine 2014, 21, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, J.; Lu, B.; Shi, Z.; Wang, H.; Zhang, B.; Zhao, K.; Qi, W.; Bao, J.; Wang, Y. Molecular switch of Akt in Polygonatum odoratum lectin-induced apoptosis and autophagy in human non-small cell lung cancer A549 cells. PLoS ONE 2014, 9, e101526. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, B.; Min, M.W.; Bian, H.J.; Chen, L.F.; Liu, Q.; Bao, J.K. Induction of apoptosis by Polygonatum odoratum lectin and its molecular mechanisms in murine fibrosarcoma L929 cells. Biochim. Biophys. Acta 2009, 1790, 840–844. [Google Scholar] [CrossRef]
- Naik, S.; Rawat, R.S.; Khandai, S.; Kumar, M.; Jena, S.S.; Vijayalakshmi, M.A.; Kumar, S. Biochemical characterisation of lectin from Indian hyacinth plant bulbs with potential inhibitory action against human cancer cells. Int. J. Biol. Macromol. 2017, 105, 1349–1356. [Google Scholar] [CrossRef]
- Sindhura, B.R.; Hegde, P.; Chachadi, V.B.; Inamdar, S.R.; Swamy, B.M. High mannose N-glycan binding lectin from Remusatia vivipara (RVL) limits cell growth, motility and invasiveness of human breast cancer cells. Biomed. Pharmacother. 2017, 93, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, L.; Wang, D.; Wang, H.; Chen, J.; Yang, C.; Bao, J.; Wu, C. Role of reactive oxygen species-mediated MAPK and NF-ĸB activation in Polygonatum cyrtonema lectin-induced apoptosis and autophagy in human lung carcinoma A549 cells. J. Biochem. 2016, 160, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, J.M.; Li, J.; Liu, J.J.; Li, W.W.; Li, C.Y.; Xu, H.L.; Bao, J.K. Polygonatum cyrtonema lectin induces murine fibrosarcoma L929 cell apoptosis and autophagy via blocking Ras-Raf and PI3K-Akt signaling pathways. Biochimie 2010, 92, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cheng, Y.; Bian, H.J.; Bao, J.K. Molecular mechanisms of Polygonatum cyrtonema lectin-induced apoptosis and autophagy in cancer cells. Autophagy 2009, 5, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cheng, Y.; Zhang, B.; Bian, H.J.; Bao, J.K. Polygonatum cyrtonema lectin induces apoptosis and autophagy in human melanoma A375 cells through a mitochondria-mediated ROS-p38-p53 pathway. Cancer Lett. 2009, 275, 54–60. [Google Scholar] [CrossRef]
- Feng, L.X.; Sun, P.; Mi, T.; Liu, M.; Liu, W.; Yao, S.; Cao, Y.M.; Yu, X.L.; Wu, W.Y.; Jiang, B.H.; et al. Agglutinin isolated from Arisema heterophyllum Blume induces apoptosis and autophagy in A549 cells through inhibiting PI3K:Akt pathway and inducing ER stress. Clin. J. Nat. Med. 2016, 14, 856–864. [Google Scholar]
- Chan, Y.S.; Yu, H.; Xia, L.; Ng, T.B. Lectin from green speckled lentil seeds (Lens culinaris) triggered apoptosis in nasopharyngeal carcinoma cell lines. Chin. Med. 2015, 10, 25. [Google Scholar] [CrossRef]
- Kumar, S.; Jitendra, K.; Singh, K.; Kappor, V.; Sinha, M.; Xess, I.; Das, S.N.; Sharma, S.; Singh, T.P.; Dey, S. Biological properties and characterization of ASL50 protein from aged Allium sativum bulbs. Appl. Biochem. Biotechnol. 2015, 176, 1914–1927. [Google Scholar] [CrossRef]
- Shi, Z.; Chen, J.; Li, C.Y.; An, N.; Wang, Z.J.; Yang, S.L.; Huang, K.E.; Bao, J.K. Antitumor effects of concanavalin A and Sophora flavescens lectin in vitro and in vivo. Acta Pharmacol. Sin. 2014, 35, 248–256. [Google Scholar] [CrossRef]
- Faheina-Martins, G.V.; da Silveira, A.L.; Cavalcanti, B.C.; Ramos, M.V.; Moraes, M.O.; Pessoa, C.; Araújo, D.A. Antiproliferative effects of lectins from Canavalia ensiformis and Canavalia brasiliensis in human leukemia cell lines. Toxicol. In Vitro 2012, 26, 1161–1169. [Google Scholar] [CrossRef]
- Pratt, J.; Roy, R.; Annabi, B. Concanavalin-A-induced autophagy biomarkers requires membrane type-1 metalloproteinase intracellular signaling in glioblastoma cells. Glycobiology 2012, 22, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.R.; Thakur, V.S.; Gupta, K.; Jackson, M.W.; Harada, H.; Agarwal, M.K.; Shin, D.M.; Wald, D.N.; Agarwal, M.L. Restoration of p53 functions protects cells from concanavalin A-induced apoptosis. Mol. Cancer Ther. 2010, 9, 471–479. [Google Scholar] [CrossRef]
- Liu, B.; Min, M.W.; Bao, J.K. Induction of apoptosis by concanavalin A ans its molecular mechanisms in cancer cells. Autophagy 2009, 5, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.O.; Santos, P.N.; Figueirôa, E.O.; de Melo, C.M.; de Andradre Lemoine Neves, J.K.; Arruda, F.V.; Cajazeiras, J.B.; do Nascimento, K.S.; Teixeira, E.H.; Cavada, B.S.; et al. Antiproliferative effect of Canavalia brasiliensis lectin on B16F10 cells. Res. Vet. Sci. 2014, 96, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Gao, Y.; Xu, S.; Yang, Z.; Xu, T. Purification of a mannose-binding lectin Pinellia ternata agglutinin and its induction of apoptosis in Bel-7404 cells. Protein Expr. Purif. 2014, 93, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Walde, P.; Miyazaki, T.; Sakayama, K.; Nakamura, A.; Kameda, K.; Masuda, S.; Umakoshi, H.; Kato, K. Active targeting to osteosarcoma cells and apoptotic cell death induction by the novel lectin Eucheuma serra agglutinin isolated from a marine red alga. J. Drug Deliv. 2012, 2012, 842785. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Sugahara, T.; Ueno, M.; Fukuta, Y.; Ochi, Y.; Akiyama, K.; Miyazaki, T.; Masuda, S.; Kawakubo, A.; Kato, K. The anti-tumor effect of Eucheuma serra agglutinin on colon cancer cells in vitro and in vivo. Anticancer Drugs 2006, 17, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.; Dan, X.; Ng, C.C.W.; Ng, T.B. Lectins with potential for anti-cancer therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Martínez, L.E.; Moreno-Celis, U.; Cervantes-Jiménez, R.; Ferriz-Martínez, R.A.; Blanco-Labra, A.; García-Gasca, T. Plant lectins as medical tools against digestive system cancers. Int. J. Mol. Sci. 2017, 18, 1403. [Google Scholar] [CrossRef] [PubMed]
- Lagarda-Dias, I.; Guzman-Partida, A.M.; Vazquez-Moreno, L. Legume lectins: Proteins with diverse applications. Int. J. Mol. Sci. 2017, 18, 1242. [Google Scholar] [CrossRef] [PubMed]
- Poiroux, G.; Barre, A.; Rougé, P.; Benoist, H. Targeting glycosylation aberrations to improve the efficiency of cancer phototherapy. Curr. Cancer Drug Targets 2018. [Google Scholar] [CrossRef] [PubMed]
- François, K.O.; Balzarini, J. Potential of carbohydrate-binding agents as therapeutics against enveloped viruses. Mol. Res. Rev. 2012, 32, 349–387. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, A.; Geonnotti, A.R.; Balzarini, J.; Kiser, P.F. Activity and safety of synthetic lectins based on benzoboroxole-functionalized polymers for inhibition of HIV entry. Mol. Pharm. 2011, 8, 2465–2475. [Google Scholar] [CrossRef] [PubMed]













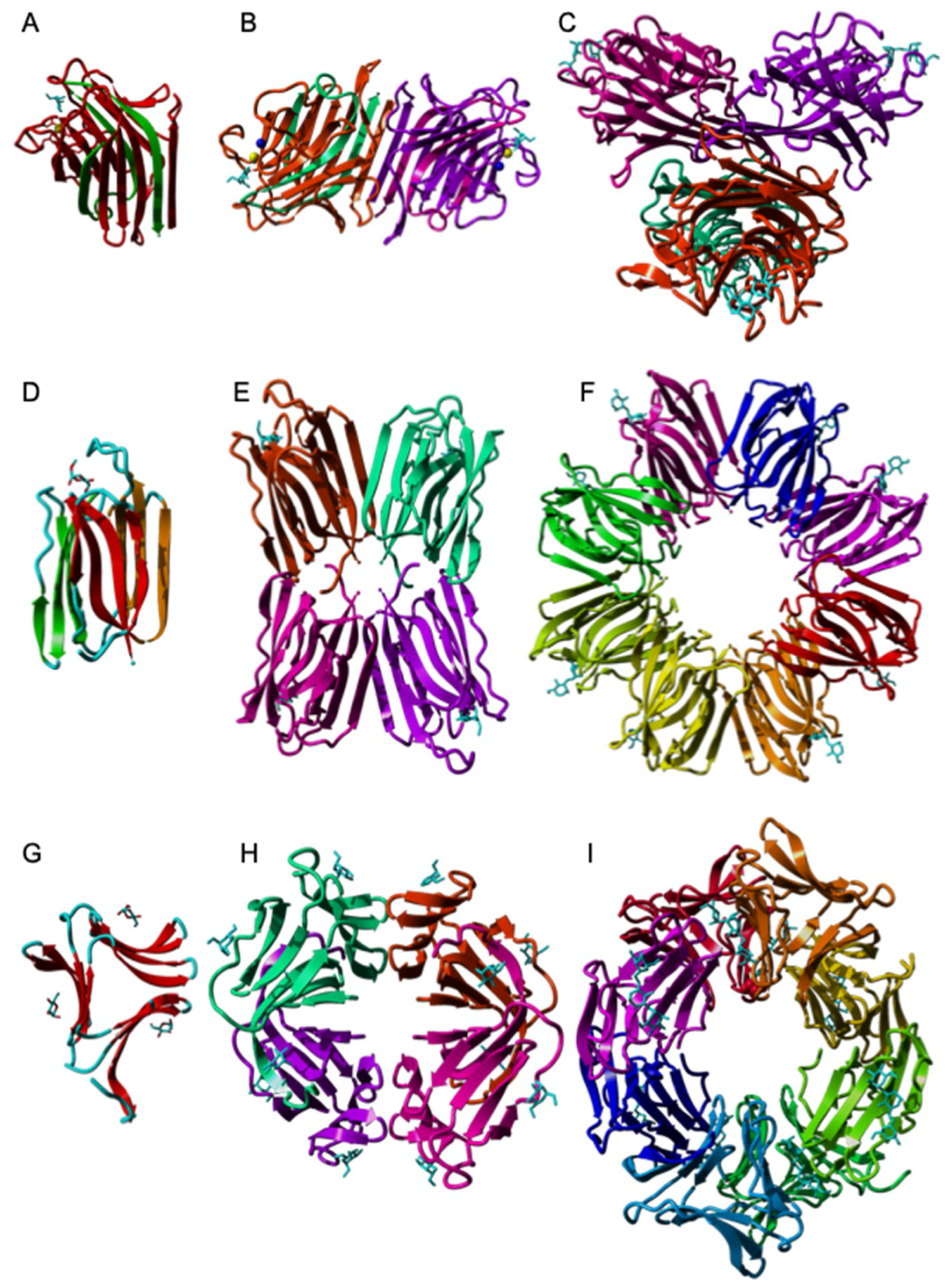{kind=link}
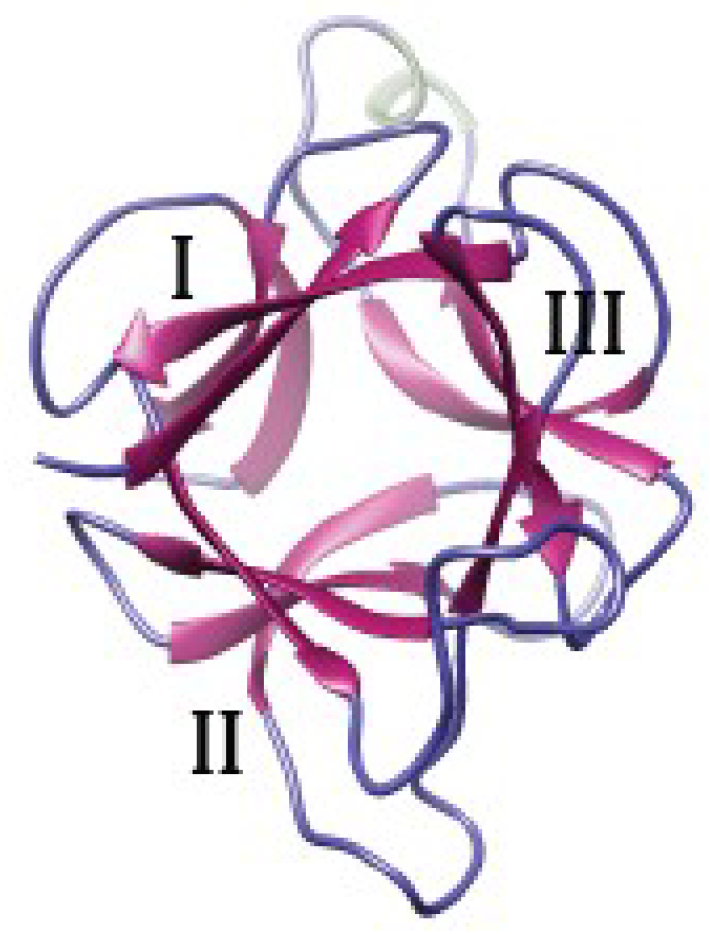{kind=link}
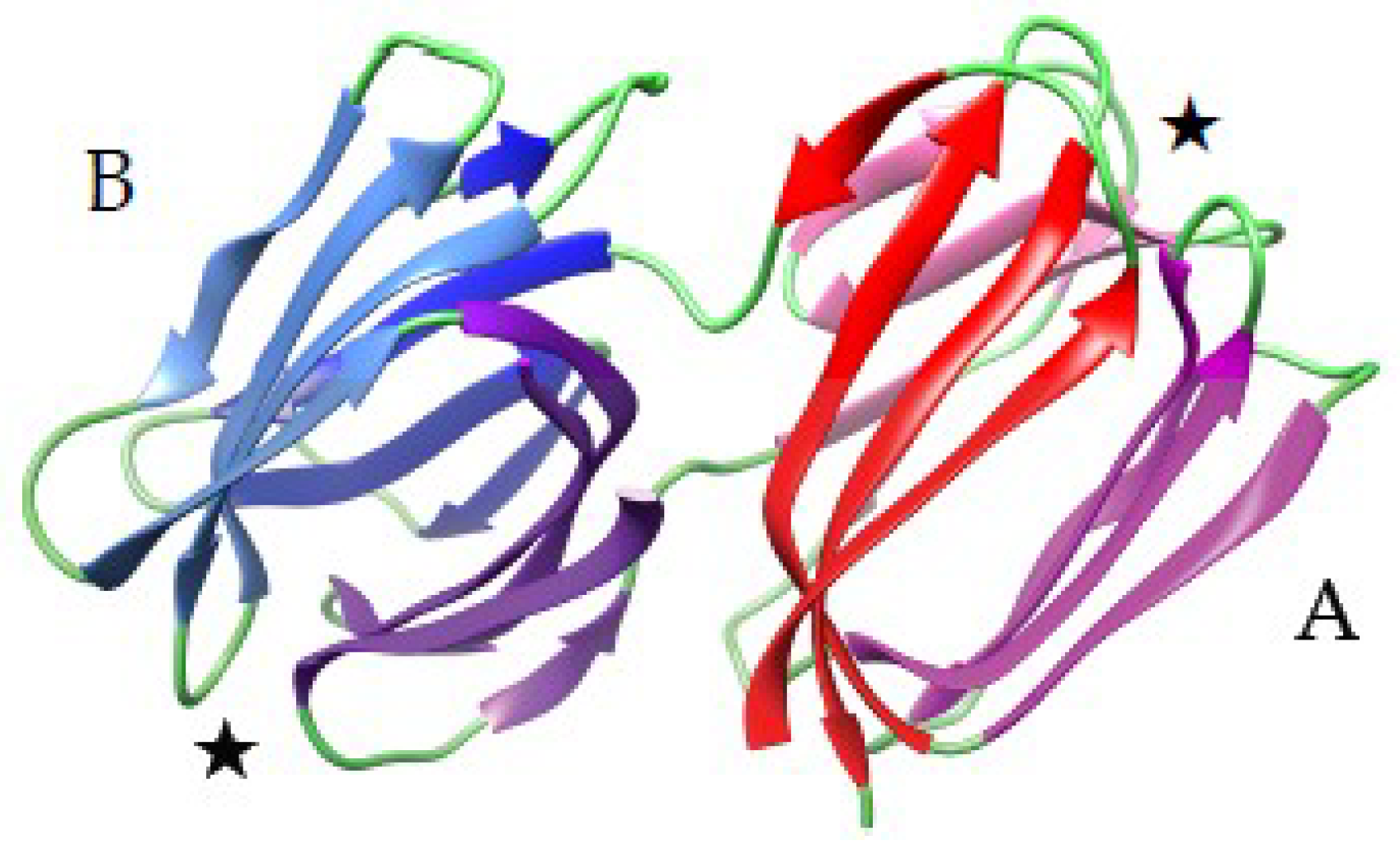{kind=link}
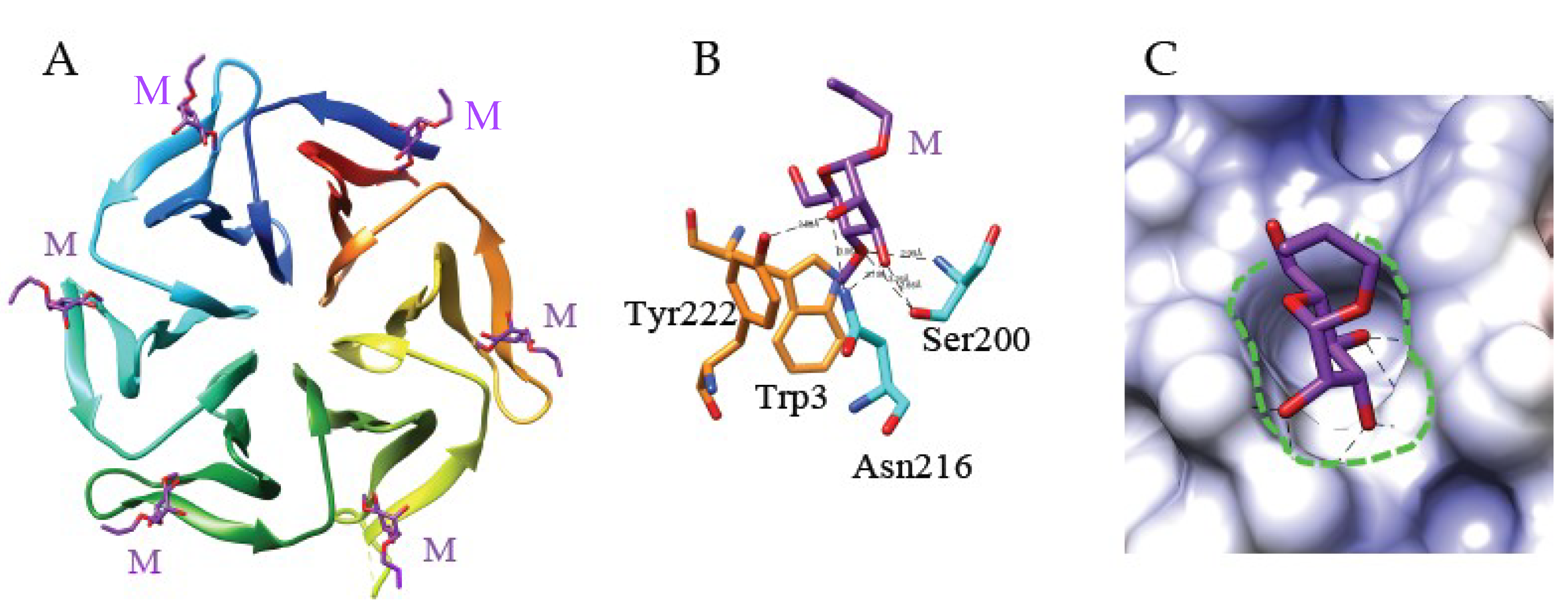{kind=link}
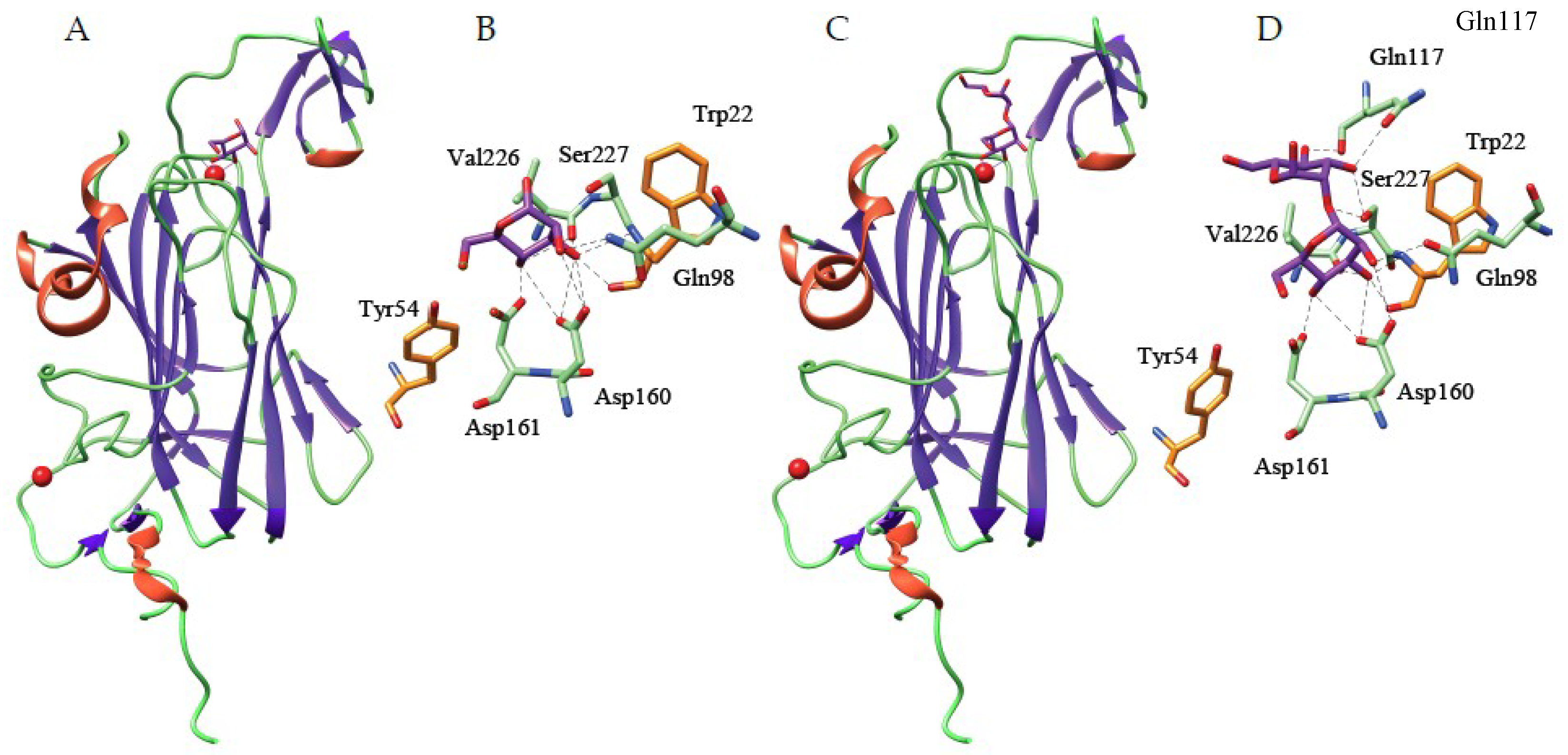{kind=link}
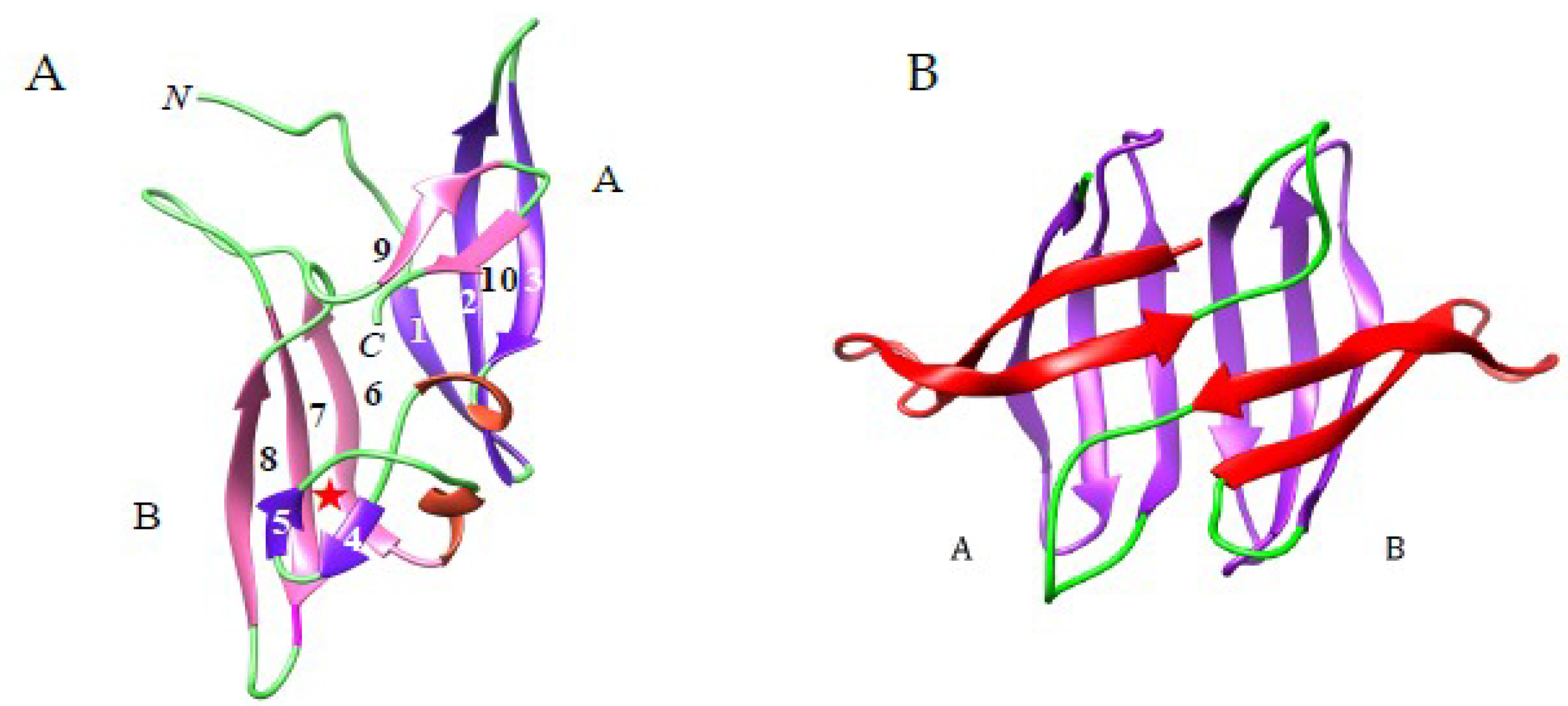{kind=link}
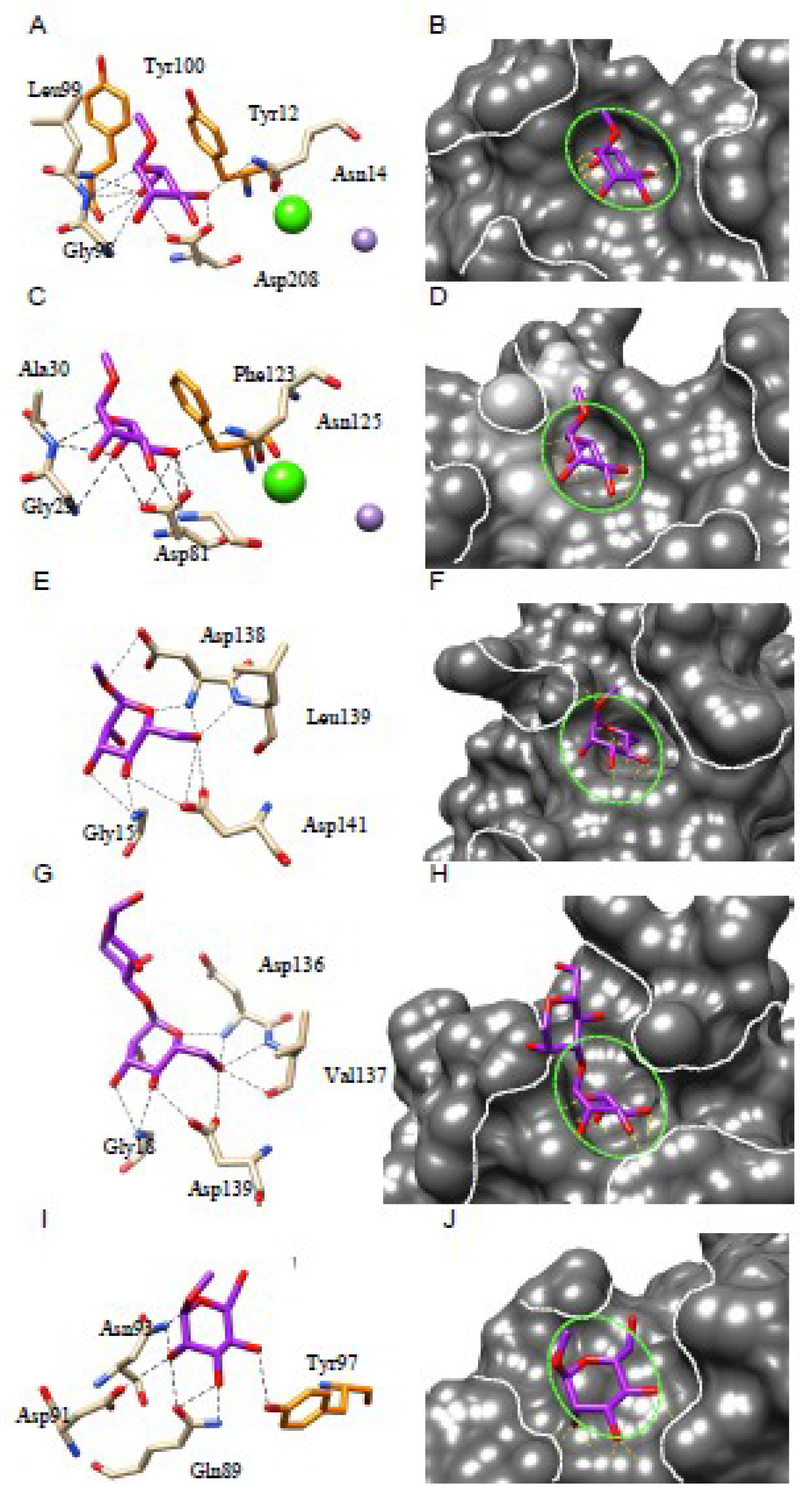{kind=link}
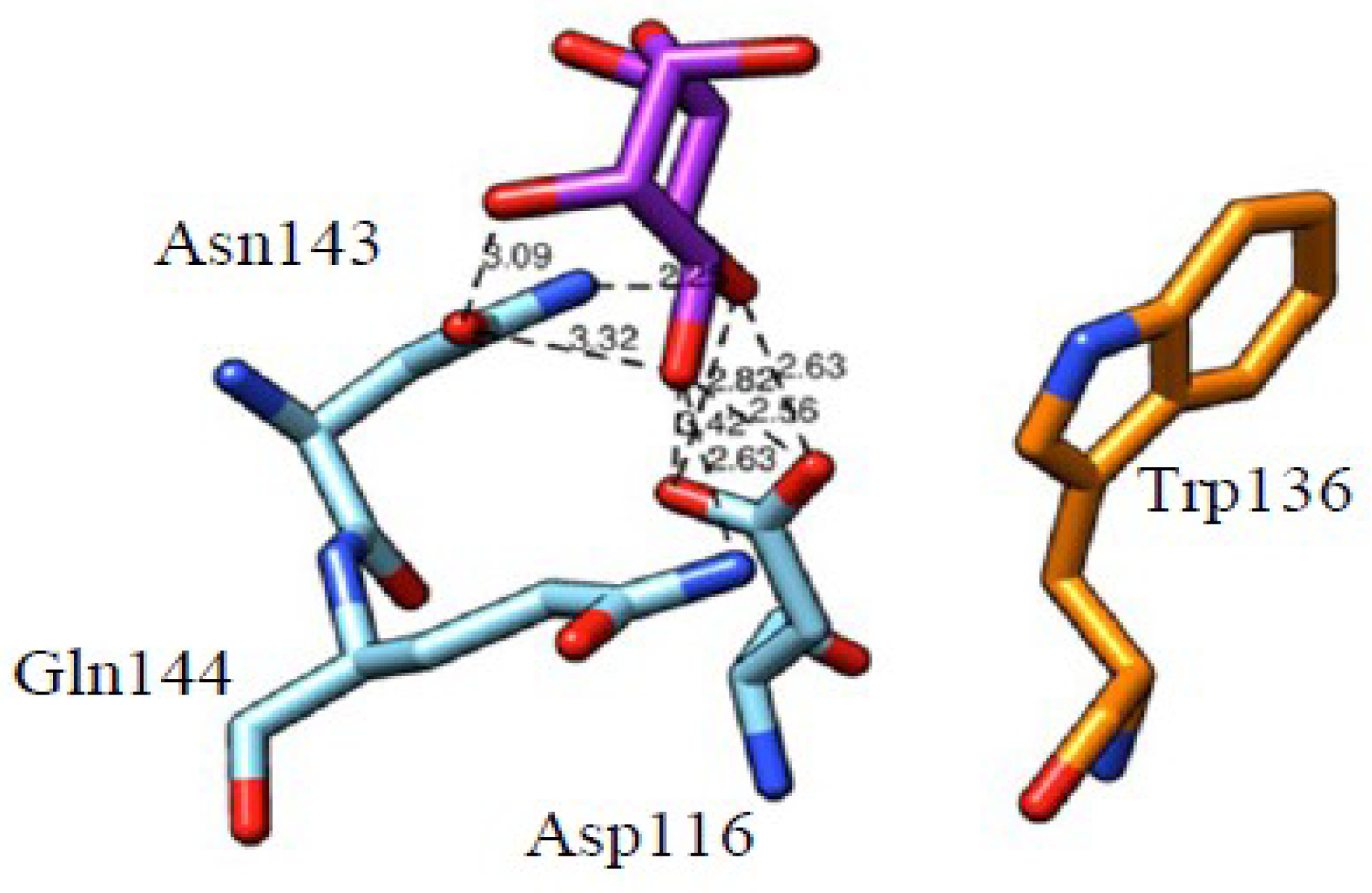{kind=link}
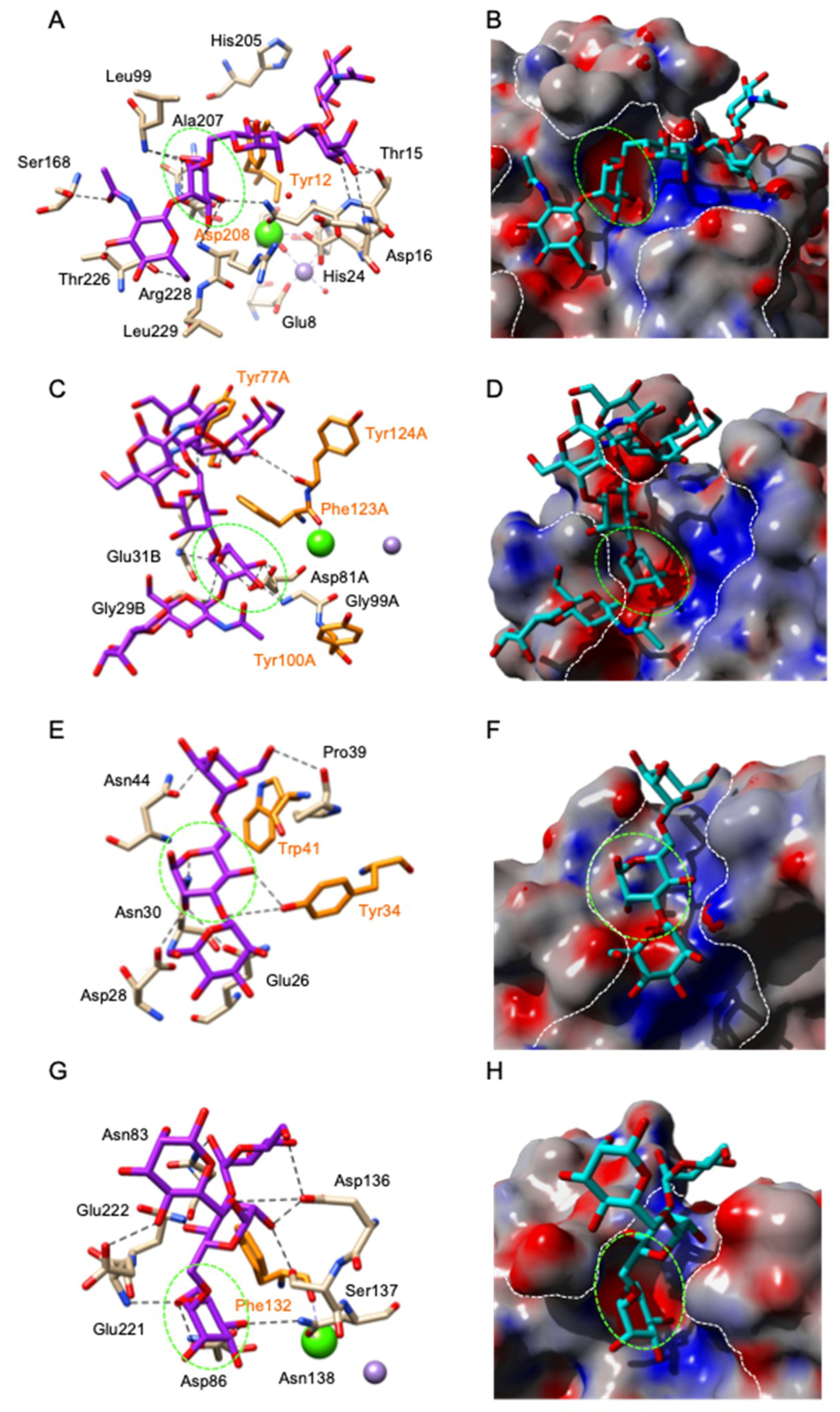{kind=link}
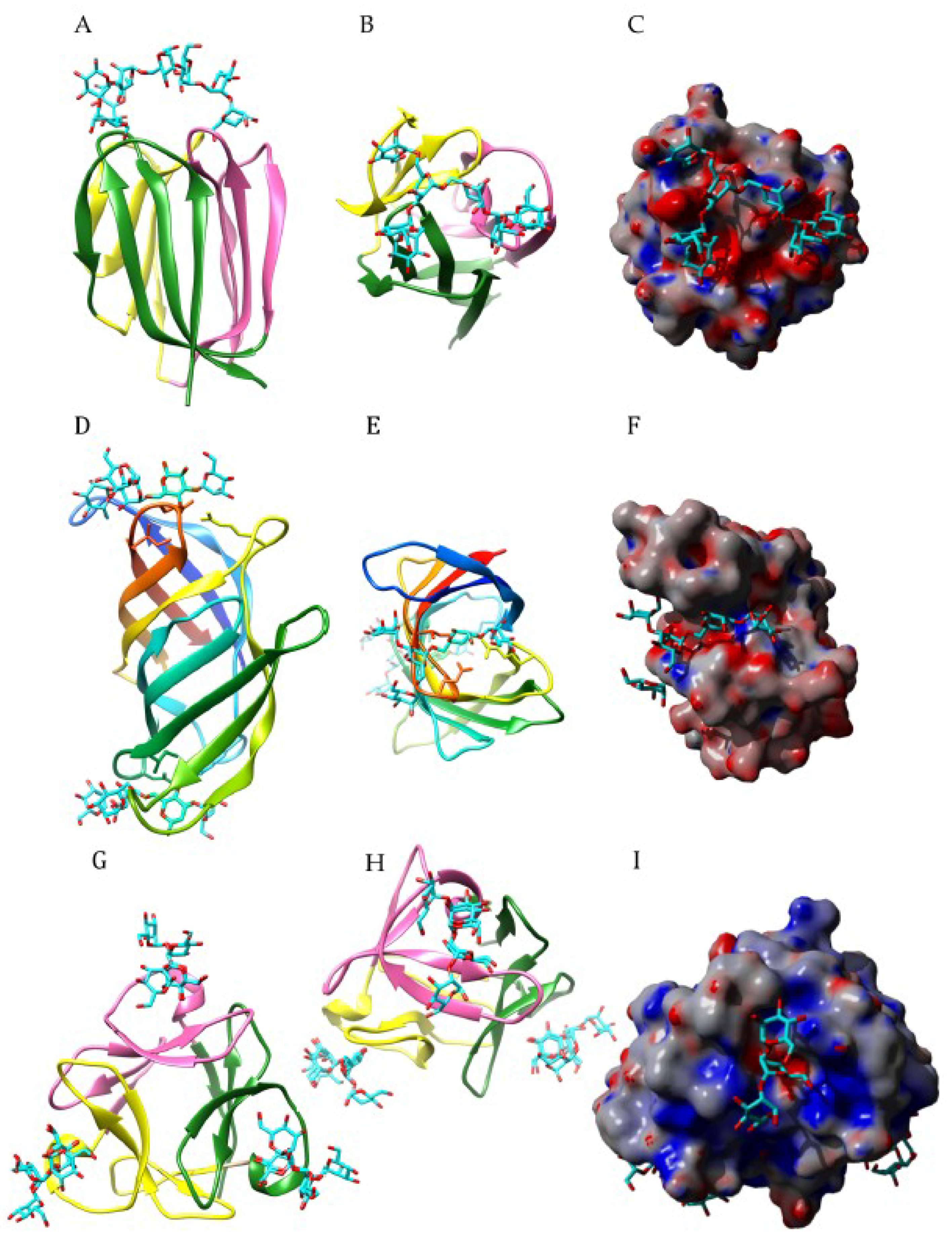{kind=link}
{kind=link}
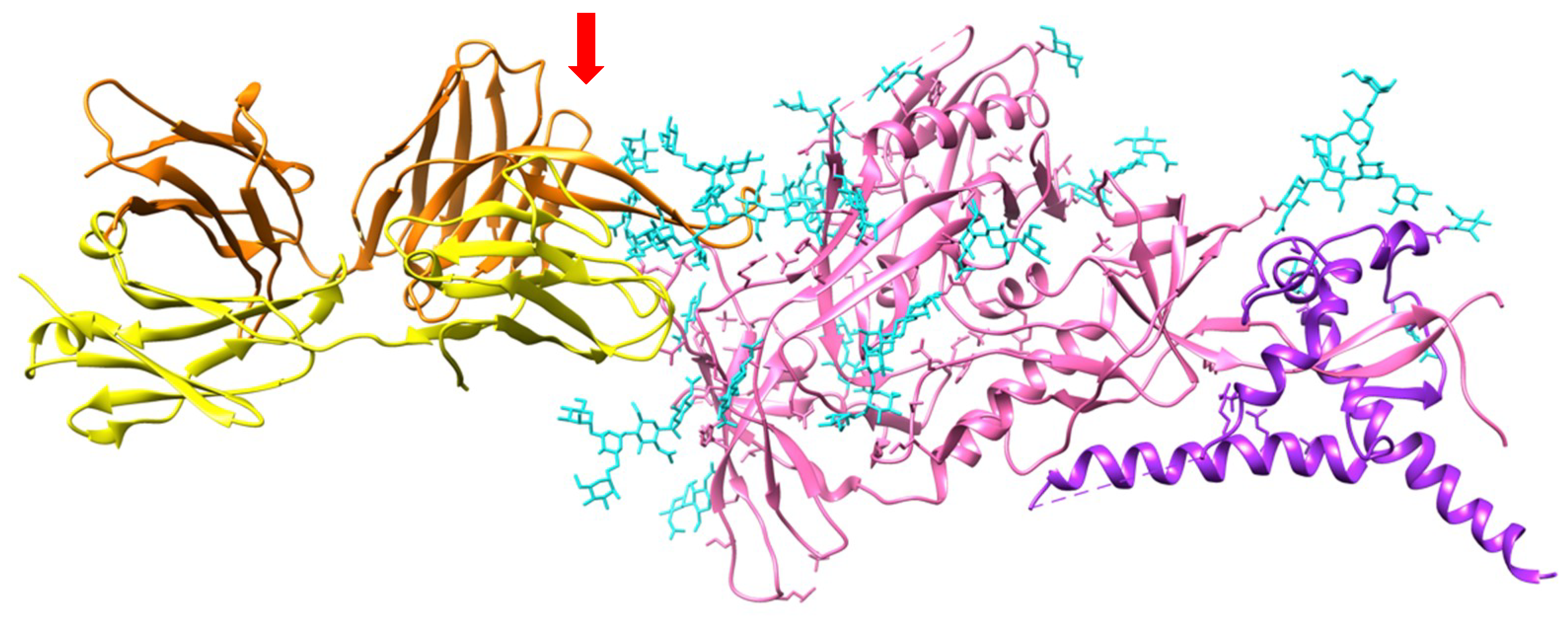{kind=link}
| Plant, Alga, Mushroom Family | Plant, Alga, Mushroom Species | Lectin | Structural Scaffold | Oligomer | Ref. |
|---|---|---|---|---|---|
| Pteridophyta | Phlebodium aureum | PAL | β barrel | 2 | [12] |
| Gymnosperms | Araucaria brasiliensis | Lectin I | n.d. | 10 | [13] |
| Lectin 2 | n.d. | 6 | |||
| Gingko biloba | Gnk2 | α β | 1 | [14] | |
| Cycas revoluta | CRLL | β-prism | 2 | [15,16] | |
| Fabaceae | Bowringia mildbraedii | BMA | β-sandwich | 2/4 | [17] |
| Cajanus cajan | CcL | βs | 2 | [18] | |
| Camptosema pedicellatum | CPL | βs | 4 | [19] | |
| Canavalia boliviana | ConBo | βs | 4 | [20] | |
| Canavalia bonariensis | CaBo | βs | 4 | [21] | |
| Canavalia brasiliensis | ConBr | βs | 4 | [22] | |
| Canavalia ensiformis | ConA | βs | 4 | [23] | |
| Canavalia gladiata | CGL | βs | 4 | [24] | |
| Canavalia grandiflora | ConGF | βs | 4 | [25] | |
| Canavalia maritima | ConM | βs | 4 | [26] | |
| Canavalia virosa | ConV | βs | 4 | [27] | |
| Centrolobium microchaete | CML | βs | 4 | [28] | |
| Centrolobium tomentosum | CTL | βs | 4 | [29] | |
| Cladrastis lutea | CLAI,II | βs | 4 | [30] | |
| Cratylia floribunda | CFL | βs | 2/4 | [31] | |
| Cratylia mollis | CRAMOLL | βs | 2/4 | [32] | |
| Cymbosema roseum | CRLI | βs | 4 | [33] | |
| Dioclea grandiflora | DGL | βs | 4 | [34,35] | |
| Dioclea guianensis | Dguia | βs | 4 | [36] | |
| Dioclea lasiocarpa | DLL | βs | 4 | [37] | |
| Dioclea lasiophylla | DlyL | βs | 4 | [38] | |
| Dioclea reflexa | DrfL | βs | 4 | [39] | |
| Dioclea rostrata | DRL | βs | 4 | [40] | |
| Dioclea sclerocarpa | DSL | βs | 4 | [41] | |
| Dioclea violacea | DVL | βs | 4 | [42] | |
| Dioclea virgata | DvirL | βs | 4 | [43] | |
| Dioclea wilsonii | DwL | βs | 4 | [44] | |
| Lathyrus aphaca | LaphL | βs | 2 | [45] | |
| Lathyrus articulatus | LarL | βs | 2 | [45] | |
| Lathyrus cicera | LcL | βs | 2 | [45] | |
| Lathyrus hirsutus | LhL | βs | 2 | [46] | |
| Lathyrus nissolia | LnL | βs | 1 | [47] | |
| Lathyrus ochrus | LoL | βs | 2 | [48] | |
| Lathyrus odoratus | LodL | βs | 2 | [49] | |
| Lathyrus sativus | LsL | βs | 2 | [50] | |
| Lathyrus sphaericus | LsphL | βs | 1 | [51] | |
| Lathyus sylvestris | LsiL | βs | 2 | [52] | |
| Lathyrus tingitanus | LtL | βs | 2 | [46] | |
| Lens culinaris | LcA | βs | 2 | [53] | |
| Millettia dielsiana | MDL | βs | 2 | [54] | |
| Onobrychis viciifolia | βs | n.d. | [55] | ||
| Pisum arvense | PAL | βs | 2 | [56] | |
| Pisum sativum | PsA | βs | 2 | [57] | |
| Pterocarpus angolensis | PAL | βs | 2 | [58] | |
| Sophora flavescens | SFL | βs | 2 | [59] | |
| Trigonella foenumgraecum | βs | n.d. | [60] | ||
| Vicia cracca | βs | 2 | [61] | ||
| Vicia ervilia | βs | 4 | [62] | ||
| Vicia faba | VfA | βs | 2 | [63] | |
| Vicia sativa | βs | 2 | [64] | ||
| Mimosaceae | Parkia biglobosa | PBL | βs | 2 | [65] |
| Parkia platycephala | PPL | βs | 2 | [66] | |
| Dalbergieae | Platypodium elegans | nPELa | βs | 2 | [67] |
| Platymiscium floribundum | PFL | βs | 2 | [68] | |
| Fagaceae | Castanea crenata | CCA | βs | 6/8 | [69] |
| Moraceae | Artocarpus heterophyllus | ArtinM | β-prism | 4 | [70,71] |
| Artocarpus incisa | Frutapin | βp | 4 | [72] | |
| Artocarpus integer | CMB | βp | 4 | [73,74] | |
| Artocarpus integrifolia | artocarpin | βp | 4 | [75,76] | |
| jacalin | βp | 4 | [77,78] | ||
| Artocarpus lakoocha | artocarpin | βp | 4 | [79] | |
| Morus nigra | Moniga-M | βp | 4 | [80] | |
| Asteraceae | Helianthus tuberosus | Heltuba | βp | 8 | [81] |
| Brassicaceae | Arabidopsis thaliana | PP2-A1 | βp | n.d. | [82] |
| Ranonculaceae | Clematis montana | CML | βp | 2 | [83] |
| Aloeae | Aloe arborescens | ALOE | βp | 4 | [84] |
| Araceae | Arisaema lobatum | ALA | n.d. | 2+2 | [85] |
| Arisaema heterophyllum | AHA | βp | n.d | [86] | |
| Arum maculatum | AMA | βp | 2+2 | [87] | |
| Colocasia esculenta | CEA, tarin | βp | 2+2 | [88] | |
| Dieffenbachia sequina | βp | 2+2 | [87] | ||
| Lysichiton camtschatcensis | βp | 2+2 | [89] | ||
| Pinellia ternata | PTA | βp | 2+2 | [90] | |
| Remusatia vivipara | RVL | βp | 2+2 | [91] | |
| Typhonium divaricatum | TDL | βp | 2+2 | [92] | |
| Xanthosoma sagittifolium | XSL | βp | 2+2 | [93] | |
| Zantedeschia aethiopica | ZAA | βp | n.d. | [94] | |
| Asparagaceae | Ophiopogon japonicus | OJL | βp | n.d. | [95] |
| Polygonatum cyrtonema | PCL | βp | 4 | [96] | |
| Polygonatum multiflorum | PMA | βp | 4 | [97] | |
| Convolvulaceae | Polygonatum odoratum | POL | βp | 4 | [98] |
| Calystegia sepium | Calsepa | βp | 2 | [99] | |
| Ipomoea batatas | ipomoelin | βp | 4 | [100] | |
| Alliaceae | Allium altaicum | AALTA | βp | 2 | [101] |
| Allium ascalonicum | AAA | βp | 2 | [102] | |
| Allium cepa | ACA | βp | 2 | [103] | |
| Allium porrum | APA | βp | 2 | [103] | |
| Allium sativum | ASA-I/II | βp | 2 | [104] | |
| Allium tuberosum | ATA | βp | 2 | [105] | |
| Allium ursinum | AUA-I/II | βp | 2 | [106] | |
| Amaryllidaceae | Amaryllis vittata | AVA | βp | n.d. | [107] |
| Clivia miniata | CMA | βs | 2 | [108] | |
| Crinum asiaticum | CAA | βp | n.d. | [109] | |
| Galanthus nivalis | GNA | βp | 4 | [110] | |
| Hippeastrum hybrid | HHA | βp | 2 | [111] | |
| Leucojum vernum | LVL | βp | n.d. | [112] | |
| Zephyranthes candida | ZCA | βp | 4 | [113] | |
| Zephyranthes grandiflora | ZGA | βp | 4 | [114] | |
| Lycoris aurea | LAA | βp | 2 | [115] | |
| Lycoris radiata | LRA | βp | 2 | [116] | |
| Dioscoreaceae | Dioscorea batatas | DB1 | βp | 1 | [117] |
| Dioscorea bulbifera | DBL | βp | 1 | [118] | |
| Iridaceae | Crocus sativus | CSL | βp | n.d. | [119,120] |
| Crocus vernus | CVA | βp | 4 | [121] | |
| Liliaceae | Aspidistra elatior | AEL | n.d. | 2 | [122] |
| Narcissus pseudonarcissus | NPA | βp | 2,4 | [111] | |
| Narcissus tazetta | NTL | βp | 2 | [123] | |
| Narcissus tortifolius | NTA | βp | n.d. | [124] | |
| Tulipa hybrid | TxLCI | βp | 4 | [125] | |
| TL-MII | βp | 2 | |||
| Smilacaceae | Smilax glabra | SGM2 | βp | 3 | [126] |
| Hyacintheae | Scilla campanulata | SCAman | βp | 2 | [127] |
| Musaceae | Musa acuminata | BanLec | βp | 2 | [128] |
| Musa paradisiaca | βp | 2 | [129] | ||
| Pandanaceae | Pandanus amaryllifolius | pandanin | βp | n.d. | [130] |
| Orchidaceae | Cymbidium hybridum | CHA | βp | 2 | [131] |
| Dendrobium officinale | DOA2 | βp | n.d. | [132] | |
| Epipactis helleborine | EHMBP | βp | 2 | [131] | |
| Gastrodia elata | gastrodianine | βp | 2 | [133] | |
| Liparis noversa | LNL | βp | 2 | [95] | |
| Listera ovata | LNL | βp | 2 | [131] | |
| Poaceae | Oryza sativa | Orysata | βp | 2 | [134] |
| Red algae | Bryothamnion seaforthii | BSL | n.d. | 1 | [135] |
| Bryothamnion triquetrum | BTL | n.d. | 1,2 | [136] | |
| Euchema denticulatum | EDA | n.d. | 1 | [137] | |
| Eucheuma serra | ESA | n.d. | 1 | [138] | |
| Griffithsia sp. | griffithsin | n.d. | 2 | [139,140] | |
| Hypnea cervicornis | HCA | n.d. | 1 | [141] | |
| Hypnea japonica | HJA | n.d. | 1 | [9] | |
| Hypnea musciformis | HMA | n.d. | 1 | [142] | |
| Kappaphycus alvarezii | KAA-2 | n.d. | 1 | [143] | |
| Kappaphycus striatum | KSA | n.d. | 1 | [144] | |
| Green algae | Boodlea coacta | BCA | β-prism | 1 | [145] |
| Halimeda renschii | HRL40-1/2 | n.d. | 4 | [146] | |
| Hydnangiaceae | Laccaria bicolor | tectonin 2 | β-propeller | n.d. | [147,148] |
| Trichocomaceae | Penicillium chrysogenum | PeCL | n.d. | n.d. | [149] |
| Saccharomycetaceae | Saccharomyces cerevisiae | Flo5A | β-sandwich | 2 | [150] |
| Saccharomyces pasteurianus | Flo1p | βs | 4 | [151] | |
| Schizosaccharo-mycetaceae | Schizosaccharomyces pombe | glucosidase | βs | 2 | [152] |
| Hygrophoraceae | Hygrophorus russula | HRL | n.d. | 4 | [153] |
| Marasmiaceae | Marasmus oreades | MOA | β-prism | 2 | [154] |
| Pteridaceae | Ceratopteris richardii | cyanovirin | CVN-fold | 1 | [155] |
| Sordariaceae | Neurospora crassa | cyanovirin | CVN-fold | 1 | [155] |
| Tuberaceae | Tuber borchii | cyanovirin | CVN-fold | 1 | [155] |
| Bacteria Family | Species | Lec Lectin | Dstructural Scscaffold | Ref. |
|---|---|---|---|---|
| Actinomycetaceae | Actinomycete sp. | actinohivin | CVN-fold | [180] |
| Bukholderiaceae | Burkholderia cenocepacia | BcLA | id. | [181] |
| Cyanothecaceae | Cyanothece sp. | Cyt-CVNH | id. | [182] |
| Nectriaceae | Gibberella zeae | Gz-CVNH | id. | [183] |
| Oscillatoriaceae | Oscillatoria agardhii | OAA | id. | [184] |
| Microcystaceae | Microcystis aeruginosa | microvirin | id. | [179] |
| Microcystis viridis | MVL | id. | [185] | |
| Myxococcaceae | Myxococcus xanthus | cyanovirin-N | id. | [186] |
| Nostocaceae | Nostoc ellipsosporum | id. | [187] | |
| Pseudomonadaceae | Pseudomonas fluorescens | PFL | id. | [186] |
| Pseudomonas putida | LLP | id. | [188] | |
| Scytonemataceae | Scytonema varium | id. | [189] | |
| Thermotogaceae | Thermotoga maritim | Tmcbm27 | β-sandwich | [190] |
| Plant Species: | Lectin: | PDB Code: | Ref. |
|---|---|---|---|
| Bowringia mildbraedii | BMA | 2FMD(o) | [194] |
| Canavalia ensiformis | ConA | 1BXH(o), 1CVN(o), 1I3H(o), 1ONA(o), 1QDC(o), 1QDO(o), 1TEI(o), 1VAM(m), 5CNA(m), 5WEY(o) | [195,196,197,198,199,200,201,202,203] |
| Canavalia gladiata | CGL | 2D7F(m), 2EF6(o), 2OVU(o) | [204,205] |
| Canavalia maritima | ConM | 2OW4(o), 2P37(o) | [205] |
| Canavalia virosa | ConV | 5F5Q(m) | [27] |
| Centrolobium tomentosum | CTL | 5EYX(o), 5EYY(o) | [29] |
| Cymbosema roseum | CRLI | 4MYE(m) | |
| Dioclea grandiflora | DGL | 1DGL(o) | [35] |
| Dioclea lasiocarpa | DLL | 5UUY(m) | [37] |
| Dioclea lasiophylla | DlyL | 6CJ9(m) | [38] |
| Dioclea reflexa | DrfL | 5TG3(m) | [39] |
| Dioclea rostrata | DRL | 2ZBJ | [40] |
| Dioclea sclerocarpa | DSL | 4NOT(m) | [41] |
| Dioclea virgata | DvirL | 3RS6(m) | [43] |
| Lathyrus ochrus | LoLI | 1LOA(m), 1LOB(m), 1LOF(o), 1LOG(o) | [191,192,206] |
| LoLII | 1LGB(c), 1LGC(c) | [193] | |
| Pisum arvense | PAL | 5T7P(m) | [207] |
| Pisum sativum | PsA | 1BQP(m), 1RIN(o) | [208,209] |
| Pterocarpus angolensis | PAL | 1Q8O(o), 1Q8P(o), 1Q8Q(o), 1Q8S(o), 1Q8V(o), 1UKG(m), 2AR6(o), 2ARB(o), 2ARE(m), 2ARX(o), 2AUY(o), 2GN3(m), 2GN7(o), 2GMM(o), 2GMP(o), 2PHF(o), 2PHR(o), 2PHT(o), 2PHU(o), 2PHW(o), 2PHX(o) | [210,211,212,213] |
| Parkia biglobosa | PBL | 4MQ0(m) | |
| Artocarpus incisa | frutapin | 5M6O(m) | [72] |
| Artocarpus integrifolia | artocarpin | 1J4U(m), 1VBO(o), 1VBP(o) | [163,214] |
| jacalin | 1KUJ(m), 1WS4(m), 1WS5(m) | [77,78] | |
| Morus nigra | Morniga-M | 1XXR(m) | [168] |
| Helianthus tuberosus | Heltuba | 1C3M(o), 1C3N(o) | [81] |
| Colocasia esculenta | tarin | 5D9Z(m), 5T20(o) | [165] |
| Ipomoea batatas | ipomoelin | 3R51(m), | [99] |
| Calystegia sepium | Calsepa | 1OUW(m), 5AV7(o), 5XF1(o) | [98] |
| Allium sativum | ASA | 1BWU(m), 1KJ1(m) | [215,216] |
| Galanthus nivalis | GNA | 1JPC(o), 1MSA(m), 1NIV(o) | [217,218] |
| Narcissus pseudonarcissus | NPA | 1NPL(o), 3DZW(o) | [219] |
| Musa acuminata | 3MIT(m), 3MIU(o), 4PIK(o), 4PIT(o) | [128,220] | |
| Musa paradisiaca | 1X1V(m) | [127] | |
| Oryza sativa | Orysata | 5XFH(c), 5XFI(c) | [221] |
| Fungal/Algal Species: | Lectin: | PDB Code: | Ref. |
| Griffthsia sp. | griffthsin | 2GUC(m), 2GUD(m), 2HYQ(o), 3LL2(c) | [140,222,223] |
| Saccharomyces cerevisiae | adhesin Flo1 | 4LHK(o), 4LHN(m) | [151] |
| Saccharomyces pastorianus | flocculin Flo5 | 2XJP(m), 2XJR(o), 2XJS(o), 2XJT(o), 2XJU(o) | [150] |
| Schizosaccharomyces pombe | glucosidase II | 4XQM(m) | [152] |
| Marasmus oreades | cyanovirin-N | 4TKC(m) | |
| Actinomyces sp. | actinohivin | 4P6A(o) | [224] |
| Oligosaccharidic Structures | Con A | LcA | Favin |
|---|---|---|---|
| Man | 1.25 | 2.5 | 0.625 |
| αMan(1,3)βΜαν(1,4)GlcNAc | 0.104 | 0.83 | 0.104 |
| αΜαν(1,2)αΜαν(1,3)βMan(1,4)GlcNAc | 0.026 | 0.21 | 0.105 |
| αΜαν(1,2)αMan(1,2)αΜαν(1,3)βΜαν(1,4)GlcNAc | 0.026 | 0.206 | 0.105 |
 | 0.0003 | 0.157 | 0.31 |
 | 0.026 | 0.003 | 0.013 |
 | 0.026 | 0.0004 | 0.0008 |
| Oligosaccharides/Glycopeptide | Complexed to: |
|---|---|
 | Con A |
 | LoLII |
 | GNA |
 | PAL |
| Lectin class | Lectin | Aphid | Ref. |
|---|---|---|---|
| Monocot lectins | GNA (Galanthus nivalis) | Aulacorthum solani | [239] |
| Myzus persicae | [240,241] | ||
| Caratovacuna lanigera | [242] | ||
| Myzus persicae | [243] | ||
| Rhopalosiphum maidis | [244] | ||
| Chrysoperla carnea* Adalia punctata*, Coccinella septempunctata* | [245] | ||
| Sitobium avenae, Schizaphis graminum, Rhopalosiphum padi | [246] | ||
| PTA (Pinellia ternata) | Myzus persicae | [247,248] | |
| PPA (Pinellia pedatisecta) | Sitobium avenae | [249] | |
| AAA (Allium altaicum) | Aphis gossypii | [100] | |
| ACA (Allium cepa)) | Myzus persicae | [250] | |
| ASA (Allium sativum) | Myzus persicae | [251] | |
| Aphis craccivora | [252] | ||
| Myzus nicotianae | [253] | ||
| Acyrthosiphon pisum | [236] | ||
| AHA (Arisaema heterophyllum) | Myzus persicae | [254] | |
| MDA (Monstera deliciosa) | Myzus persicae | [255] | |
| Orysata (Oryza sativa) | Acyrthosiphon pisum, Myzus persicae | [256] | |
| Dieffenbachia sequina | Aphis craccivora, Lipaphis erysimi | [233] | |
| CEA (Colocacia esculenta) | Lipaphis erysimi, Aphis craccivora | [233] | |
| AMA (Arum maculatum) | Aphis craccivora, Lipaphis erysimi | [234] | |
| ZGA (Zephyranthes grandiflora) | Myzus nicotianae | [257] | |
| Legume lectins | Con A (Canavalia ensiformis) | Acyrthosiphon pisum | [235] |
| Rhopalosiphum padi | [232] | ||
| β-prism lectins | HTA (Helianthus tuberosus) | Myzus persicae | [258] |
| Fungal lectins | PeCl (Penicillium chrysogenum) | Myzus persicae | [149] |
| Lectin Families | Carbohydrate-Binding Specificity |
|---|---|
| Jacalin-related lectin family | High-mannose N-glycans |
| EUL-related lectin family | Galactosides, high-mannose N-glycans |
| Nictaba-related lectin family | Chitooligosaccharides, recognition of the (GlcNAc)2-Man3 core of high-mannose N-glycans and complex glycans |
| Lectin Class | Lectin | Ref. |
|---|---|---|
| Vicieae lectins | Con A (Canavalia ensiformis) | [276,286,287,288,289,290,291,292,293,294] |
| LcA (Lens culinaris) | [280] | |
| LoLI (Lathyrus ochrus) | [280] | |
| PsA (Pisum sativum) | [280,295] | |
| Monocot lectins | CHA (Cymbidium hybrid) | [280,296] |
| EHA (Epipactis helleborine) | [280,296] | |
| GNA (Galanthus nivalis) | [280,297,298,299,300,301,302] | |
| HHA (Hippeastrum hybrid) | [280,297,302] | |
| LOA (Listera ovata) | [280,297] | |
| NPA (Narcissus pseudonarcissus) | [280,297,303] | |
| NTA (Narcissus tazetta) | [304] | |
| Narcissus confusus, N. leonensis and N. tortifolius | [124] | |
| PCL (Polygonatum cytonema) | [96,305] | |
| Convolvulaceae | Calsepa (Calystegia sepium) | [280] |
| Urticaceae | UDA (Urtica dioica) | [296] |
| Araceae | RVL (Remusatia vivipara) | [91] |
| Musaceae | BanLec (Musa acuminata) | [306] |
| Poaceae | GNAmaize (Zea mays) | [307] |
| Asteraceae | Heltuba (Helianthus tuberosus) | [280] |
| Red algae | Griffithsin (Griffithsia sp.) | [139,308] |
| Green algae | KAA (Kappaphycus alvarezii) | [309] |
| BCA (Boodlea coacta) | [143] |
| Lectin | Cancer Cell | Apoptosis | Autophagy | Ref. |
|---|---|---|---|---|
| Cabo (Canavalia bonariensis) | glioma | + | [21] | |
| PsA (Pisum sativum) | colorectal cancer | + | [331] | |
| Erlich acites carcinoma | + | [332] | ||
| MOSL (Moringa oleifera) | Erlich acites carcinoma, | + | [333] | |
| murine malanoma | + | [334] | ||
| DLasiL (Dioclea lasiocarpa) | glioma | + | [37] | |
| ovarian, lung, brestbreast, prostate carcinoma | + | [335] | ||
| POL (Polygonatum odoratum) | melanoma | + | + | [336] |
| lung adenocarcinoma | + | + | [337] | |
| breast cancer | + | + | [338] | |
| lung cancer (non-small cells) | + | [339] | ||
| melanoma | + | [98] | ||
| murine fibrosarcoma | + | [340] | ||
| Hyacinthus sp. | Caco-2, Hela | + | [341] | |
| RVL (Remusatia vivipara) | breast cancer | + | + | [342] |
| ArtinM (Artocarpus heterophyllus) | Jurkat T cells | + | + | [70] |
| PCL (Polygonatum cyrtonema) | lung adenocarcinoma | + | [343] | |
| murine fibrosarcoma | + | + | [344] | |
| melanoma | + | + | [345] | |
| melanoma | + | + | [346] | |
| AHA (Arisema heterophyllum) | lung cancer | + | [347] | |
| LcA (Lens culinaris) | nasopharyngeal carcinoma | + | [348] | |
| ASA (Allium sativum) | oral carcinoma | + | [349] | |
| Con A (Canavalia ensiformis) | breast carcinoma | + | [350] | |
| leukemia | + | + | [351] | |
| glioblastoma | + | + | [352] | |
| ovarian cancer | + | + | [353] | |
| melanoma | + | + | [354] | |
| SFL (Sophora flavescens) | HeLa cells | + | [59] | |
| breast carcinoma | + | [350] | ||
| CML (Clematis montana) | HeLa, breast cancer, | + | [83] | |
| hepatocellular carcinoma | ||||
| ConBr (Canavalia brasiliensis) | murine melanoma | + | [355] | |
| leukemia | [351] | |||
| PTA (Pinellia ternata) | hepatoma | + | [356] | |
| ESA (Eucheuma serra) | osteosarcoma | + | [357] | |
| mice colon adenocarcinoma | + | [358] | ||
| colon cancer, HeLa | + | [138] | ||
| OJL (Ophiopogon japonicus) | murine fibrosarcoma | + | [96] | |
| LNL (Liparis noversa) | murine fribrosarcoma | + | [96] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barre, A.; Bourne, Y.; Van Damme, E.J.M.; Rougé, P. Overview of the Structure–Function Relationships of Mannose-Specific Lectins from Plants, Algae and Fungi. Int. J. Mol. Sci. 2019, 20, 254. https://doi.org/10.3390/ijms20020254
Barre A, Bourne Y, Van Damme EJM, Rougé P. Overview of the Structure–Function Relationships of Mannose-Specific Lectins from Plants, Algae and Fungi. International Journal of Molecular Sciences. 2019; 20(2):254. https://doi.org/10.3390/ijms20020254
Chicago/Turabian StyleBarre, Annick, Yves Bourne, Els J. M. Van Damme, and Pierre Rougé. 2019. "Overview of the Structure–Function Relationships of Mannose-Specific Lectins from Plants, Algae and Fungi" International Journal of Molecular Sciences 20, no. 2: 254. https://doi.org/10.3390/ijms20020254
APA StyleBarre, A., Bourne, Y., Van Damme, E. J. M., & Rougé, P. (2019). Overview of the Structure–Function Relationships of Mannose-Specific Lectins from Plants, Algae and Fungi. International Journal of Molecular Sciences, 20(2), 254. https://doi.org/10.3390/ijms20020254