Cytogenetics and Cytogenomics Evaluation in Cancer


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
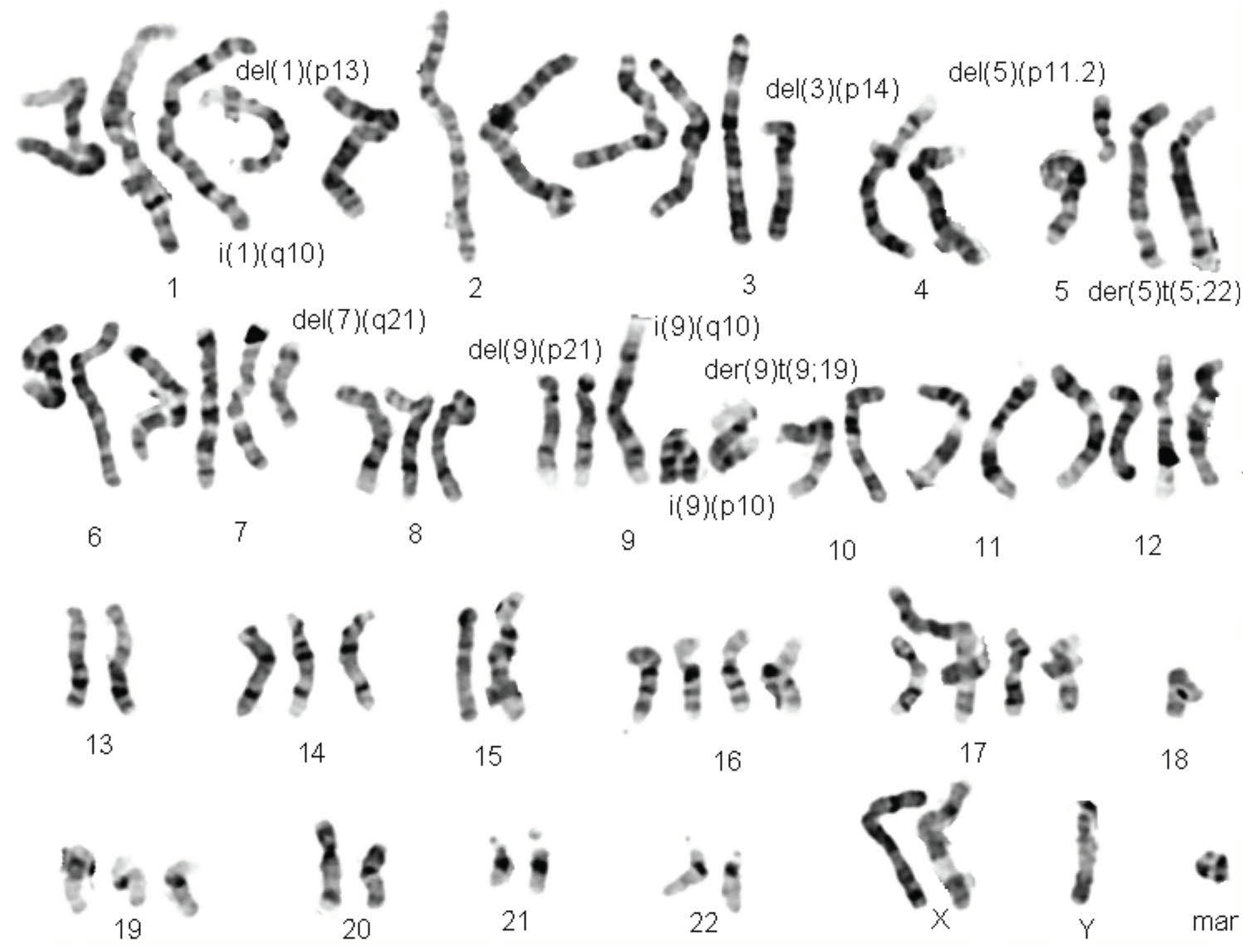
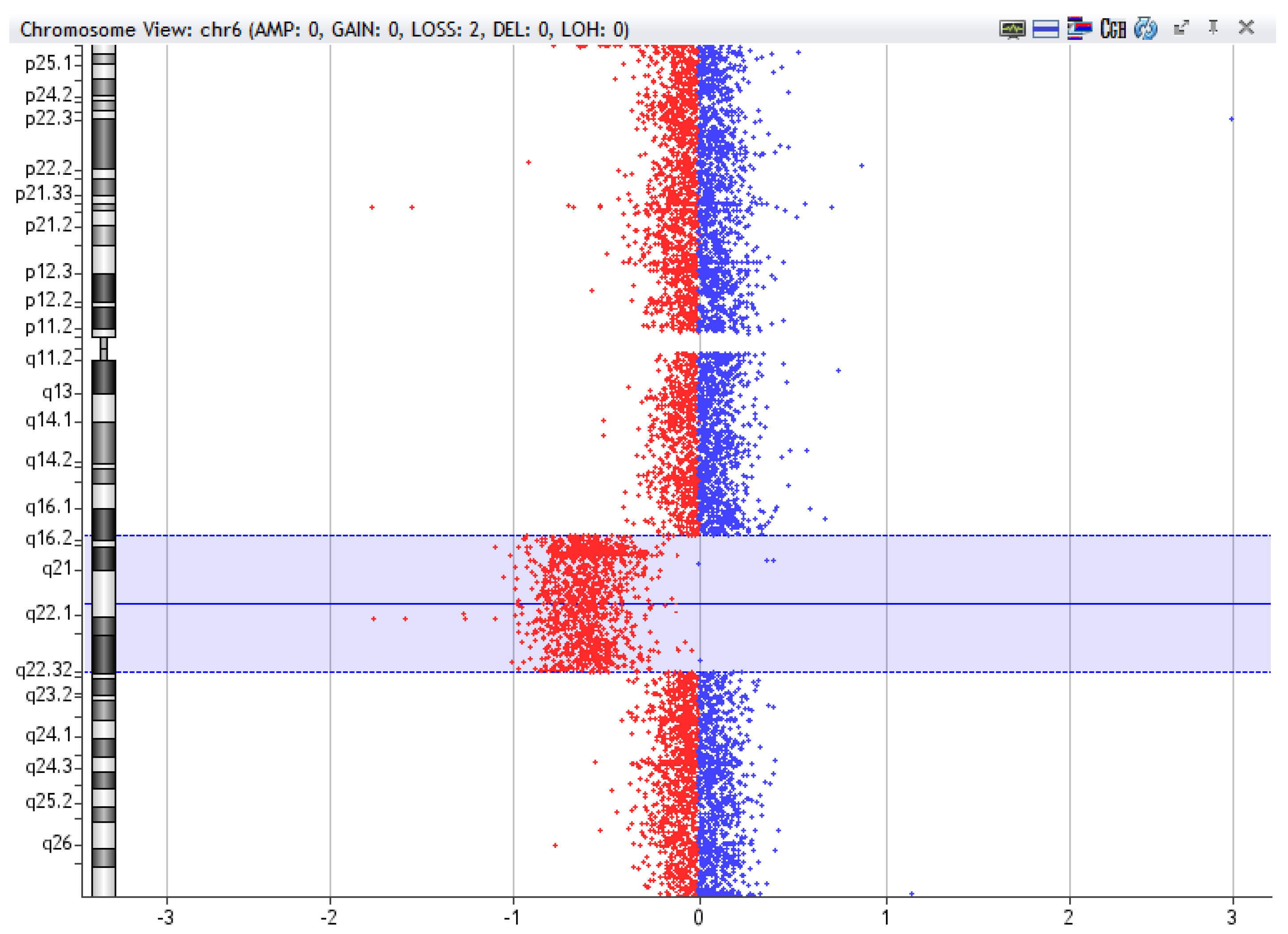
2. Technological Evolution: From Cytogenetics to Cytogenomics Analysis
3. Cytogenetic and Genomic Rearrangements in Cancer
4. Heterogeneity in Cancer
5. Biomarkers in Cancer
6. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Pinkel, D.; Albertson, D.G. Array comparative genomic hybridization and its applications in cancer. Nat. Genet. 2005, 37, S11–S17. [Google Scholar] [CrossRef] [PubMed]
- Oehler, V.G.; Yeung, K.Y.; Choi, Y.E.; Bumgarner, R.E.; Raftery, A.E.; Radich, J.P. The derivation of diagnostic markers of chronic myeloid leukemia progression from microarray data. Blood 2009, 114, 3292–3298. [Google Scholar] [CrossRef]
- Bernheim, A. Cytogenomics of cancers: From chromosome to sequence. Mol. Oncol. 2010, 4, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.B. Attacking cancer at its root. Cell 2009, 138, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthi, B.V.; Nepal, S.; Varambally, S. Genomic and Epigenomic Alterations in Cancer. Am. J. Pathol. 2016, 186, 1724–1735. [Google Scholar] [CrossRef] [PubMed]
- McGranahan, N.; Swanton, C. Clonal heterogeneity and tumor evolution: Past, present, and the future. Cell 2017, 168, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Mody, R.J.; Wu, Y.M.; Lonigro, R.J.; Cao, X.; Roychowdhury, S.; Vats, P.; Frank, K.M.; Prensner, J.R.; Asangani, I.; Palanisamy, N.; et al. Integrative clinical sequencing in the management of refractory or relapsed cancer in youth. Jama 2015, 314, 913–925. [Google Scholar] [CrossRef]
- Heppt, M.V.; Siepmann, T.; Engel, J.; Schubert-Fritschle, G.; Eckel, R.; Mirlach, L.; Kirchner, T.; Jung, A.; Gesierich, A.; Ruzicka, T.; et al. Prognostic significance of BRAF and NRAS mutations in melanoma: A German study from routine care. BMC Cancer 2017, 17, 536. [Google Scholar] [CrossRef]
- Ribeiro, I.P.; de Melo, J.B.; Carreira, I.M. Head and neck cancer: Searching for genomic and epigenetic biomarkers in body fluids—The state of art. Mol. Cytogenet. 2019, 12, 33. [Google Scholar] [CrossRef]
- Wan, T.S. Cancer cytogenetics: An introduction. In Cancer Cytogenetics; Humana Press: New York, NY, USA, 2017; Volume 1541, pp. 1–10. [Google Scholar]
- Gollin, S.M. Acquired chromosome abnormalities: The cytogenetics of cancer. In Encyclopedia of Genetics, Genomics, Proteomics and Bioinformatics; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2005; pp. 1–14. [Google Scholar]
- Das, K.; Tan, P. Molecular cytogenetics: Recent developments and applications in cancer. Clin. Genet. 2013, 84, 315–325. [Google Scholar] [CrossRef]
- Cui, C.; Shu, W.; Li, P. Fluorescence in situ hybridization: Cell-based genetic diagnostic and research applications. Front. Cell Dev. Biol. 2016, 4, 89. [Google Scholar] [CrossRef] [PubMed]
- Kearney, L. Molecular cytogenetics. Best Pract. Res. Clin. Haematol. 2001, 14, 645–669. [Google Scholar] [CrossRef] [PubMed]
- Weier, H.U.; Greulich-Bode, K.M.; Ito, Y.; Lersch, R.A.; Fung, J. FISH in cancer diagnosis and prognostication: From cause to course of disease. Expert Rev. Mol. Diagn. 2002, 2, 109–119. [Google Scholar] [CrossRef]
- Dimashkieh, H.; Wolff, D.J.; Smith, T.M.; Houser, P.M.; Nietert, P.J.; Yang, J. Evaluation of urovysion and cytology for bladder cancer detection: A study of 1835 paired urine samples with clinical and histologic correlation. Cancer Cytopathol. 2013, 121, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Ried, T.; Liyanage, M.; du Manoir, S.; Heselmeyer, K.; Auer, G.; Macville, M.; Schrock, E. Tumor cytogenetics revisited: Comparative genomic hybridization and spectral karyotyping. J. Mol. Med. 1997, 75, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.J.; Wilson, I.M.; Lam, W.L. Array CGH technologies and their applications to cancer genomes. Chromosome Res. 2005, 13, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Solinas-Toldo, S.; Lampel, S.; Stilgenbauer, S.; Nickolenko, J.; Benner, A.; Dohner, H.; Cremer, T.; Lichter, P. Matrix-based comparative genomic hybridization: Biochips to screen for genomic imbalances. Genes Chromosomes Cancer 1997, 20, 399–407. [Google Scholar] [CrossRef]
- Pollack, J.R.; Perou, C.M.; Alizadeh, A.A.; Eisen, M.B.; Pergamenschikov, A.; Williams, C.F.; Jeffrey, S.S.; Botstein, D.; Brown, P.O. Genome-wide analysis of DNA copy-number changes using cDNA microarrays. Nat. Genet. 1999, 23, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, W.W.; Chari, R.; Chi, B.; Lam, W.L. Recent advances in array comparative genomic hybridization technologies and their applications in human genetics. Eur. J. Hum. Genet. 2006, 14, 139–148. [Google Scholar] [CrossRef]
- Li, P.; Cui, C. A broader view of cancer cytogenetics: From nuclear aberrations to cytogenomic abnormalities. J. Mol. Genet. Med. 2017, 10, e108. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Homig-Holzel, C.; Savola, S. Multiplex ligation-dependent probe amplification (MLPA) in tumor diagnostics and prognostics. Diagn. Mol. Pathol. 2012, 21, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.P.; Marques, F.; Caramelo, F.; Pereira, J.; Patricio, M.; Prazeres, H.; Ferrao, J.; Juliao, M.J.; Castelo-Branco, M.; de Melo, J.B.; et al. Genetic gains and losses in oral squamous cell carcinoma: Impact on clinical management. Cell. Oncol. 2014, 37, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.P.; Marques, F.; Caramelo, F.; Ferrao, J.; Prazeres, H.; Juliao, M.J.; Rifi, W.; Savola, S.; de Melo, J.B.; Baptista, I.P.; et al. Genetic imbalances detected by multiplex ligation-dependent probe amplification in a cohort of patients with oral squamous cell carcinoma-the first step towards clinical personalized medicine. Tumour Biol. 2014, 35, 4687–4695. [Google Scholar] [CrossRef] [PubMed]
- Xuan, J.; Yu, Y.; Qing, T.; Guo, L.; Shi, L. Next-generation sequencing in the clinic: Promises and challenges. Cancer Lett. 2013, 340, 284–295. [Google Scholar] [CrossRef]
- Kamps, R.; Brandao, R.D.; Bosch, B.J.; Paulussen, A.D.; Xanthoulea, S.; Blok, M.J.; Romano, A. Next-generation sequencing in oncology: Genetic diagnosis, risk prediction and cancer classification. Int. J. Mol. Sci. 2017, 18, 308. [Google Scholar] [CrossRef]
- Parisi, F.; Micsinai, M.; Strino, F.; Ariyan, S.; Narayan, D.; Bacchiocchi, A.; Cheng, E.; Xu, F.; Li, P.; Kluger, H.; et al. Integrated analysis of tumor samples sheds light on tumor heterogeneity. Yale J. Biol. Med. 2012, 85, 347–361. [Google Scholar]
- Navin, N.E. The first five years of single-cell cancer genomics and beyond. Genome Res. 2015, 25, 1499–1507. [Google Scholar] [CrossRef]
- Liang, S.B.; Fu, L.W. Application of single-cell technology in cancer research. Biotechnol. Adv. 2017, 35, 443–449. [Google Scholar] [CrossRef]
- Navin, N.E. Cancer genomics: One cell at a time. Genome Biol. 2014, 15, 452. [Google Scholar] [CrossRef]
- Sierant, M.C.; Choi, J. Single-cell ssequencing in cancer: Recent applications to immunogenomics and multi-omics tools. Genom. Inform. 2018, 16, e17. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Surrey, L.F.; Luo, M.; Chang, F.; Li, M.M. The genomic era of clinical oncology: Integrated genomic analysis for precision cancer care. Cytogenet. Genome Res. 2016, 150, 162–175. [Google Scholar] [CrossRef]
- Albertson, D.G.; Collins, C.; McCormick, F.; Gray, J.W. Chromosome aberrations in solid tumors. Nat. Genet. 2003, 34, 369–376. [Google Scholar] [CrossRef]
- Mitelman, F.; Johansson, B.; Mandahl, N.; Mertens, F. Clinical significance of cytogenetic findings in solid tumors. Cancer Genet. Cytogenet. 1997, 95, 1–8. [Google Scholar] [CrossRef]
- Grade, M.; Difilippantonio, M.J.; Camps, J. Patterns of chromosomal aberrations in solid tumors. Recent Results Cancer Res. 2015, 200, 115–142. [Google Scholar]
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The cancer genome. Nature 2009, 458, 719–724. [Google Scholar] [CrossRef]
- Miller, D.G. On the nature of susceptibility to cancer. The presidential address. Cancer 1980, 46, 1307–1318. [Google Scholar] [CrossRef]
- Talbot, S.J.; Crawford, D.H. Viruses and tumours-an update. Eur. J. Cancer 2004, 40, 1998–2005. [Google Scholar] [CrossRef]
- Min, J.W.; Kim, W.J.; Han, J.A.; Jung, Y.J.; Kim, K.T.; Park, W.Y.; Lee, H.O.; Choi, S.S. Identification of distinct tumor subpopulations in lung adenocarcinoma via single-cell RNA-seq. PLoS ONE 2015, 10, e0135817. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Greaves, M.; Maley, C.C. Clonal evolution in cancer. Nature 2012, 481, 306–313. [Google Scholar] [CrossRef]
- Khoo, B.L.; Chaudhuri, P.K.; Ramalingam, N.; Tan, D.S.; Lim, C.T.; Warkiani, M.E. Single-cell profiling approaches to probing tumor heterogeneity. Int. J. Cancer 2016, 139, 243–255. [Google Scholar] [CrossRef]
- Greaves, M. Evolutionary determinants of cancer. Cancer Discov. 2015, 5, 806–820. [Google Scholar] [CrossRef]
- Raynaud, F.; Mina, M.; Tavernari, D.; Ciriello, G. Pan-cancer inference of intra-tumor heterogeneity reveals associations with different forms of genomic instability. PLoS Genet. 2018, 14, e1007669. [Google Scholar] [CrossRef]
- Kelloff, G.J.; Sigman, C.C. Cancer biomarkers: Selecting the right drug for the right patient. Nat. Rev. Drug Discov. 2012, 11, 201–214. [Google Scholar] [CrossRef]
- Navin, N.E.; Hicks, J. Tracing the tumor lineage. Mol. Oncol. 2010, 4, 267–283. [Google Scholar] [CrossRef]
- Sottoriva, A.; Spiteri, I.; Piccirillo, S.G.; Touloumis, A.; Collins, V.P.; Marioni, J.C.; Curtis, C.; Watts, C.; Tavare, S. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 4009–4014. [Google Scholar] [CrossRef]
- Tan, D.S.; Camilleri-Broet, S.; Tan, E.H.; Alifano, M.; Lim, W.T.; Bobbio, A.; Zhang, S.; Ng, Q.S.; Ang, M.K.; Iyer, N.G.; et al. Intertumor heterogeneity of non-small-cell lung carcinomas revealed by multiplexed mutation profiling and integrative genomics. Int. J. Cancer 2014, 135, 1092–1100. [Google Scholar] [CrossRef]
- Gerlinger, M.; Rowan, A.J.; Horswell, S.; Math, M.; Larkin, J.; Endesfelder, D.; Gronroos, E.; Martinez, P.; Matthews, N.; Stewart, A.; et al. Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. N. Engl. J. Med. 2012, 366, 883–892. [Google Scholar] [CrossRef]
- Polyak, K. Heterogeneity in breast cancer. J. Clin. Investig. 2011, 121, 3786–3788. [Google Scholar] [CrossRef]
- Brocks, D.; Assenov, Y.; Minner, S.; Bogatyrova, O.; Simon, R.; Koop, C.; Oakes, C.; Zucknick, M.; Lipka, D.B.; Weischenfeldt, J.; et al. Intratumor DNA methylation heterogeneity reflects clonal evolution in aggressive prostate cancer. Cell Rep. 2014, 8, 798–806. [Google Scholar] [CrossRef]
- Fisher, R.; Pusztai, L.; Swanton, C. Cancer heterogeneity: Implications for targeted therapeutics. Br. J. Cancer 2013, 108, 479–485. [Google Scholar] [CrossRef]
- Chatterjee, S.K.; Zetter, B.R. Cancer biomarkers: Knowing the present and predicting the future. Future Oncol. 2005, 1, 37–50. [Google Scholar] [CrossRef]
- Ribeiro, I.P.; Barroso, L.; Marques, F.; Melo, J.B.; Carreira, I.M. Early detection and personalized treatment in oral cancer: The impact of omics approaches. Mol. Cytogenet. 2016, 9, 85. [Google Scholar] [CrossRef]
- Goossens, N.; Nakagawa, S.; Sun, X.; Hoshida, Y. Cancer biomarker discovery and validation. Trans. Cancer Res. 2015, 4, 256–269. [Google Scholar]
- Roukos, D.H. Integrated clinical genomics: New horizon for diagnostic and biomarker discoveries in cancer. Expert Rev. Mol. Diagn. 2013, 13, 1–4. [Google Scholar] [CrossRef][Green Version]
- Ribeiro, I.P.; Caramelo, F.; Marques, F.; Domingues, A.; Mesquita, M.; Barroso, L.; Prazeres, H.; Juliao, M.J.; Baptista, I.P.; Ferreira, A.; et al. WT1, MSH6, GATA5 and PAX5 as epigenetic oral squamous cell carcinoma biomarkers—A short report. Cell. Oncol. 2016, 39, 573–582. [Google Scholar] [CrossRef]
- Paris, P.L.; Andaya, A.; Fridlyand, J.; Jain, A.N.; Weinberg, V.; Kowbel, D.; Brebner, J.H.; Simko, J.; Watson, J.E.; Volik, S.; et al. Whole genome scanning identifies genotypes associated with recurrence and metastasis in prostate tumors. Hum. Mol. Genet. 2004, 13, 1303–1313. [Google Scholar] [CrossRef]
- Callagy, G.; Pharoah, P.; Chin, S.F.; Sangan, T.; Daigo, Y.; Jackson, L.; Caldas, C. Identification and validation of prognostic markers in breast cancer with the complementary use of array-CGH and tissue microarrays. J. Pathol. 2005, 205, 388–396. [Google Scholar] [CrossRef]
- Weiss, M.M.; Kuipers, E.J.; Postma, C.; Snijders, A.M.; Pinkel, D.; Meuwissen, S.G.; Albertson, D.; Meijer, G.A. Genomic alterations in primary gastric adenocarcinomas correlate with clinicopathological characteristics and survival. Cell. Oncol. 2004, 26, 307–317. [Google Scholar]
- Rubio-Moscardo, F.; Climent, J.; Siebert, R.; Piris, M.A.; Martin-Subero, J.I.; Nielander, I.; Garcia-Conde, J.; Dyer, M.J.; Terol, M.J.; Pinkel, D.; et al. Mantle-cell lymphoma genotypes identified with CGH to BAC microarrays define a leukemic subgroup of disease and predict patient outcome. Blood 2005, 105, 4445–4454. [Google Scholar] [CrossRef]
- Romond, E.H.; Perez, E.A.; Bryant, J.; Suman, V.J.; Geyer, C.E., Jr.; Davidson, N.E.; Tan-Chiu, E.; Martino, S.; Paik, S.; Kaufman, P.A.; et al. Trastuzumab plus adjuvant chemotherapy for operable HER2-positive breast cancer. N. Engl. J. Med. 2005, 353, 1673–1684. [Google Scholar] [CrossRef]
- Bang, Y.J.; Van Cutsem, E.; Feyereislova, A.; Chung, H.C.; Shen, L.; Sawaki, A.; Lordick, F.; Ohtsu, A.; Omuro, Y.; Satoh, T.; et al. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label, randomised controlled trial. Lancet 2010, 376, 687–697. [Google Scholar] [CrossRef]
- Kantarjian, H.; Sawyers, C.; Hochhaus, A.; Guilhot, F.; Schiffer, C.; Gambacorti-Passerini, C.; Niederwieser, D.; Resta, D.; Capdeville, R.; Zoellner, U.; et al. Hematologic and cytogenetic responses to imatinib mesylate in chronic myelogenous leukemia. N. Engl. J. Med. 2002, 346, 645–652. [Google Scholar] [CrossRef]
- Amado, R.G.; Wolf, M.; Peeters, M.; Van Cutsem, E.; Siena, S.; Freeman, D.J.; Juan, T.; Sikorski, R.; Suggs, S.; Radinsky, R.; et al. Wild-type KRAS is required for panitumumab efficacy in patients with metastatic colorectal cancer. J. Clin. Oncol. 2008, 26, 1626–1634. [Google Scholar] [CrossRef]
- Dematteo, R.P.; Ballman, K.V.; Antonescu, C.R.; Maki, R.G.; Pisters, P.W.; Demetri, G.D.; Blackstein, M.E.; Blanke, C.D.; von Mehren, M.; Brennan, M.F.; et al. Adjuvant imatinib mesylate after resection of localised, primary gastrointestinal stromal tumour: A randomised, double-blind, placebo-controlled trial. Lancet 2009, 373, 1097–1104. [Google Scholar] [CrossRef]
- Pao, W.; Girard, N. New driver mutations in non-small-cell lung cancer. Lancet. Oncol. 2011, 12, 175–180. [Google Scholar] [CrossRef]
- Chapman, P.B.; Hauschild, A.; Robert, C.; Haanen, J.B.; Ascierto, P.; Larkin, J.; Dummer, R.; Garbe, C.; Testori, A.; Maio, M.; et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 2011, 364, 2507–2516. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Myasoedova, V.A.; Grechko, A.V.; Melnichenko, A.A.; Orekhov, A.N. New biomarkers for diagnosis and prognosis of localized prostate cancer. Semin. Cancer Biol. 2018, 52, 9–16. [Google Scholar] [CrossRef]
- Paik, S.; Shak, S.; Tang, G.; Kim, C.; Baker, J.; Cronin, M.; Baehner, F.L.; Walker, M.G.; Watson, D.; Park, T.; et al. A multigene assay to predict recurrence of tamoxifen-treated, node-negative breast cancer. N. Engl. J. Med. 2004, 351, 2817–2826. [Google Scholar] [CrossRef]
- Brandao, M.; Ponde, N.; Piccart-Gebhart, M. Mammaprint: A comprehensive review. Future Oncol. 2019, 15, 207–224. [Google Scholar] [CrossRef]
- Boutros, P.C. The path to routine use of genomic biomarkers in the cancer clinic. Genome Res. 2015, 25, 1508–1513. [Google Scholar] [CrossRef]
- Ma, M.; Zhu, H.; Zhang, C.; Sun, X.; Gao, X.; Chen, G. ″Liquid biopsy″-ctDNA detection with great potential and challenges. Ann. Transl. Med. 2015, 3, 235. [Google Scholar]
- De Rubis, G.; Rajeev Krishnan, S.; Bebawy, M. Liquid biopsies in cancer diagnosis, monitoring, and prognosis. Trends Pharmacol. Sci. 2019, 40, 172–186. [Google Scholar] [CrossRef]
- Kwapisz, D. The first liquid biopsy test approved. Is it a new era of mutation testing for non-small cell lung cancer? Ann. Transl. Med. 2017, 5, 46. [Google Scholar] [CrossRef]
- Riethdorf, S.; O’Flaherty, L.; Hille, C.; Pantel, K. Clinical applications of the CellSearch platform in cancer patients. Adv. Drug Deliv. Rev. 2018, 125, 102–121. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, I.P.; Melo, J.B.; Carreira, I.M. Cytogenetics and Cytogenomics Evaluation in Cancer. Int. J. Mol. Sci. 2019, 20, 4711. https://doi.org/10.3390/ijms20194711
Ribeiro IP, Melo JB, Carreira IM. Cytogenetics and Cytogenomics Evaluation in Cancer. International Journal of Molecular Sciences. 2019; 20(19):4711. https://doi.org/10.3390/ijms20194711
Chicago/Turabian StyleRibeiro, Ilda Patrícia, Joana Barbosa Melo, and Isabel Marques Carreira. 2019. "Cytogenetics and Cytogenomics Evaluation in Cancer" International Journal of Molecular Sciences 20, no. 19: 4711. https://doi.org/10.3390/ijms20194711
APA StyleRibeiro, I. P., Melo, J. B., & Carreira, I. M. (2019). Cytogenetics and Cytogenomics Evaluation in Cancer. International Journal of Molecular Sciences, 20(19), 4711. https://doi.org/10.3390/ijms20194711