Insight into the Phytoremediation Capability of Brassica juncea (v. Malopolska): Metal Accumulation and Antioxidant Enzyme Activity

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
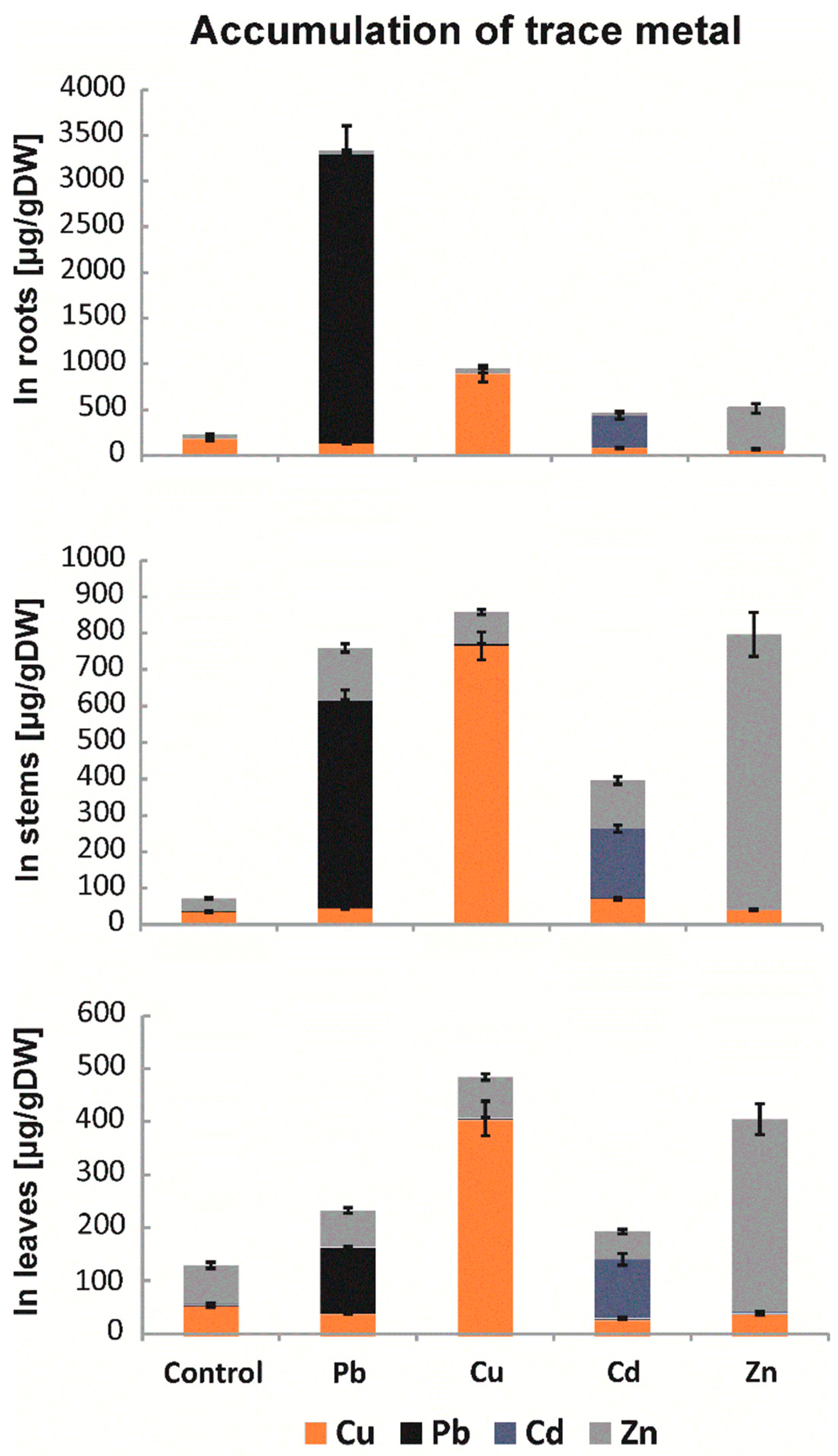
2.1. Levels of Metal Accumulation
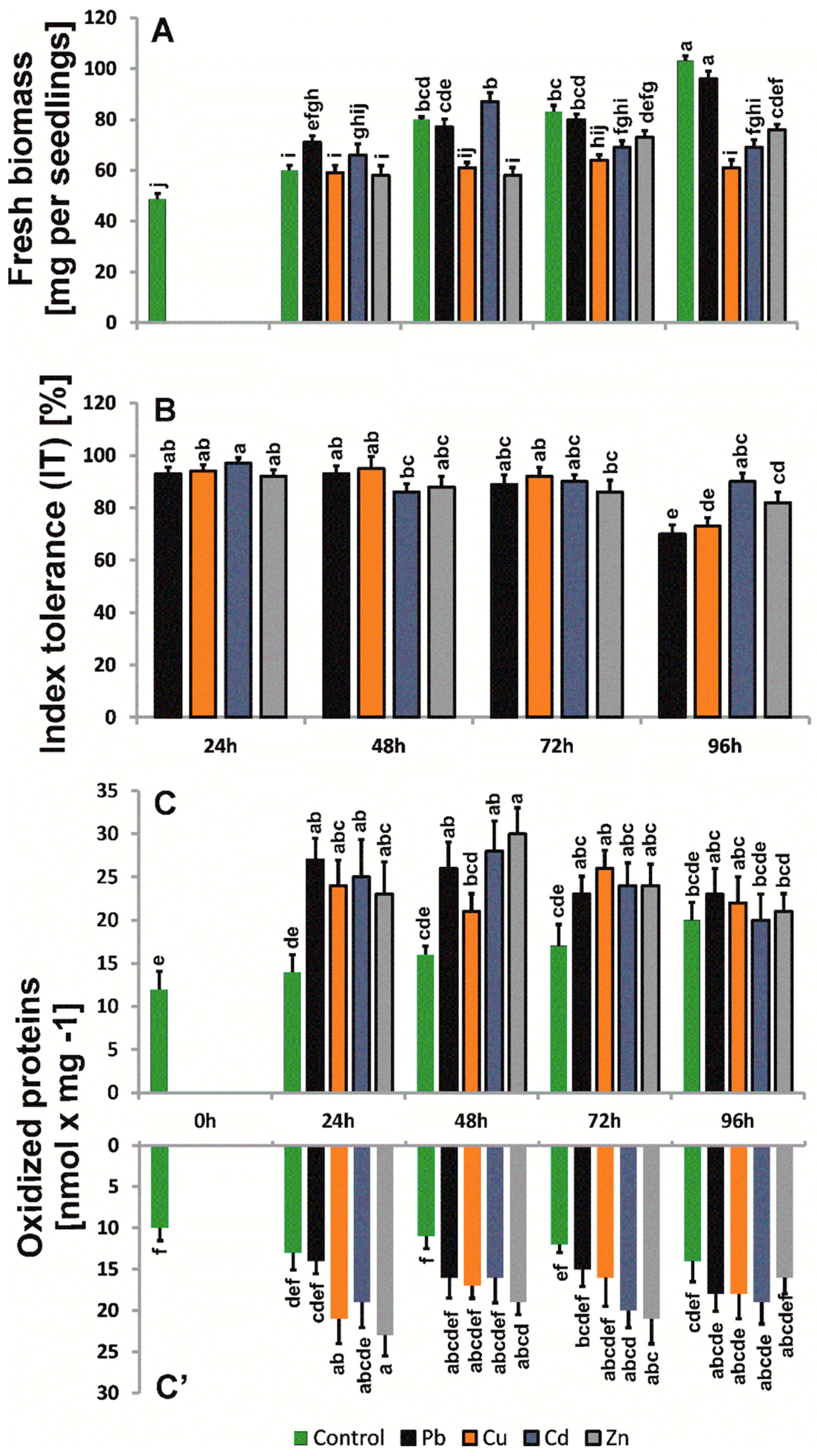
2.2. Biomass and Morphological Changes
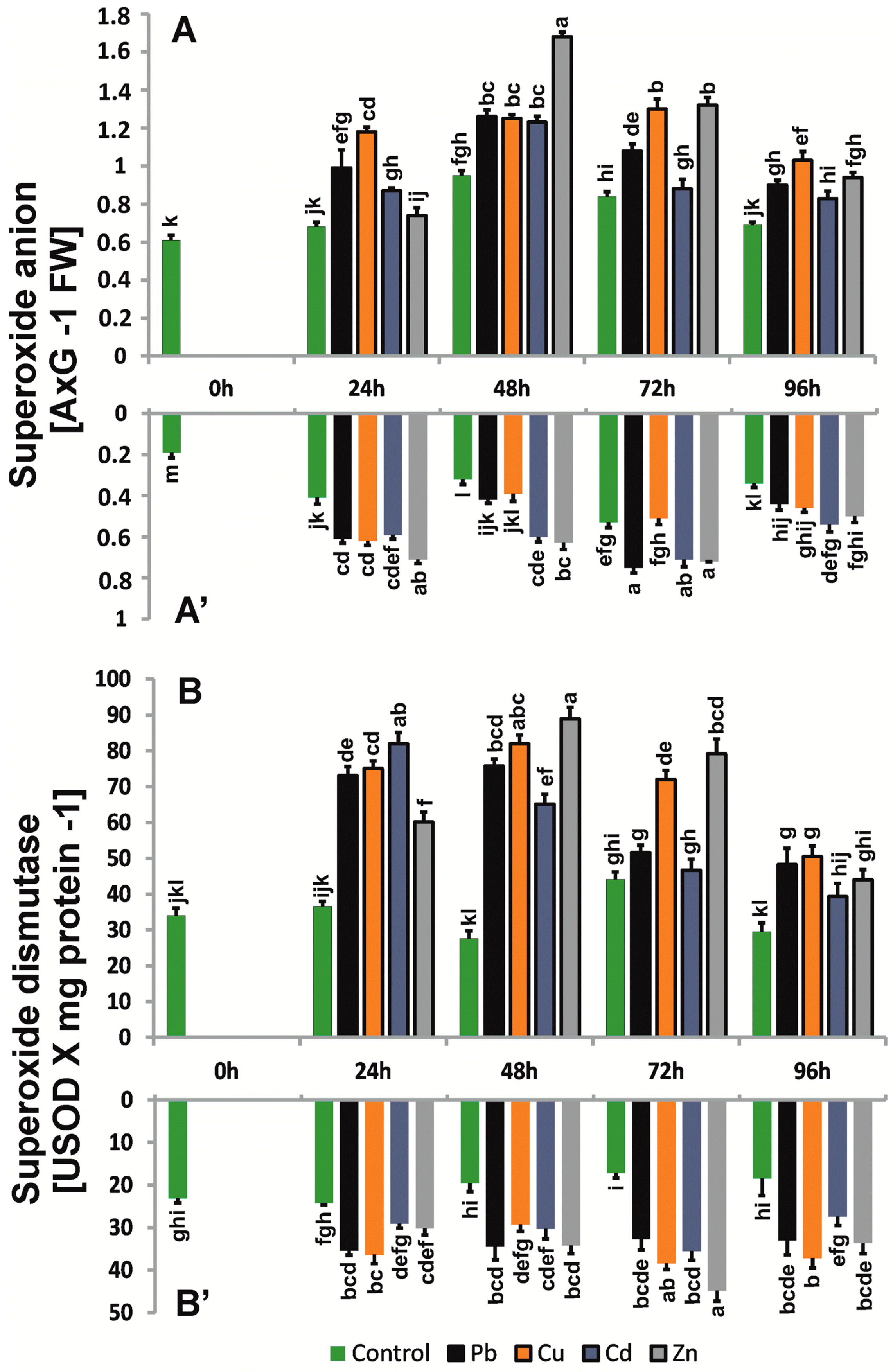
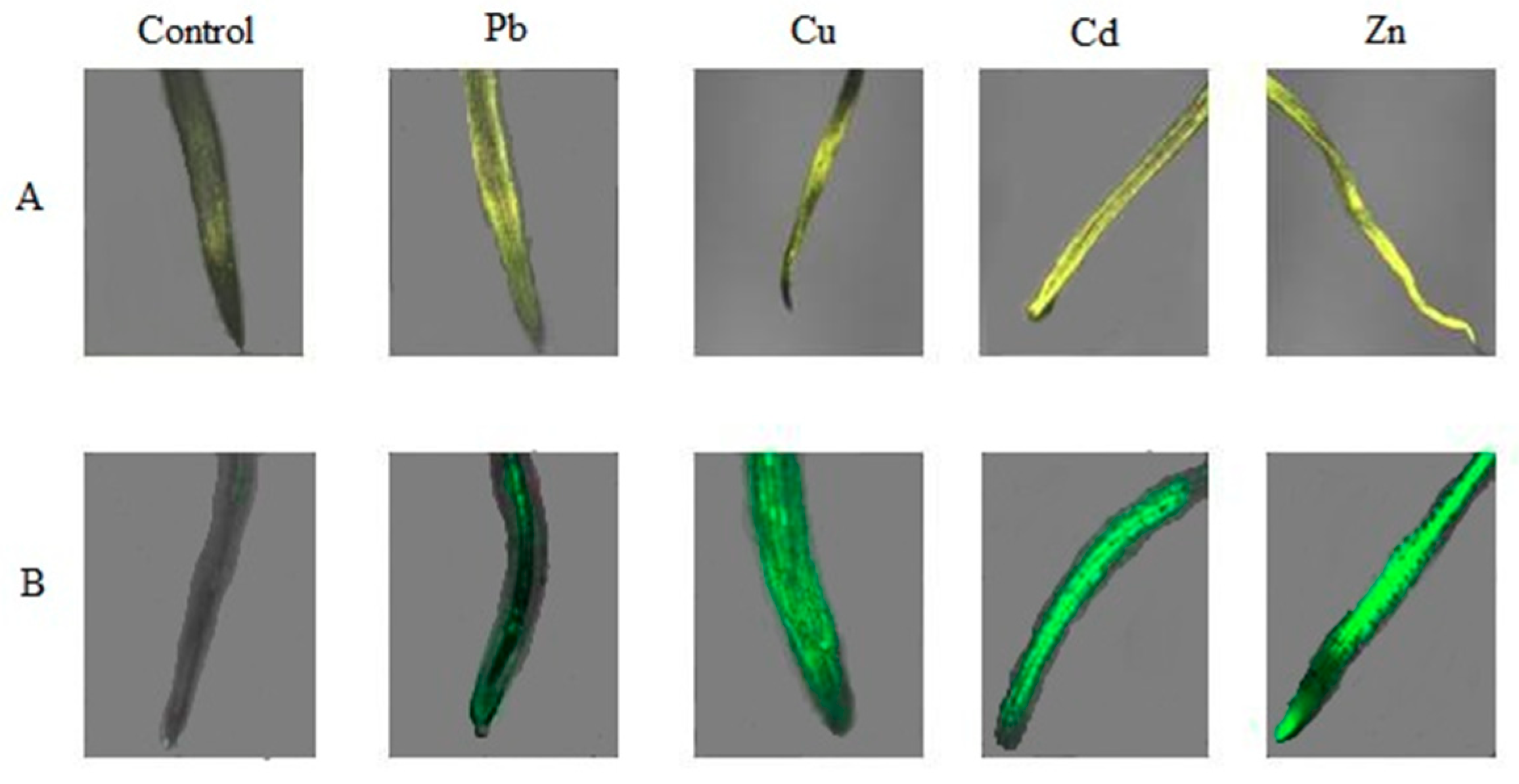
2.3. Production and Localization of ROS
2.4. Levels of Oxidized Proteins
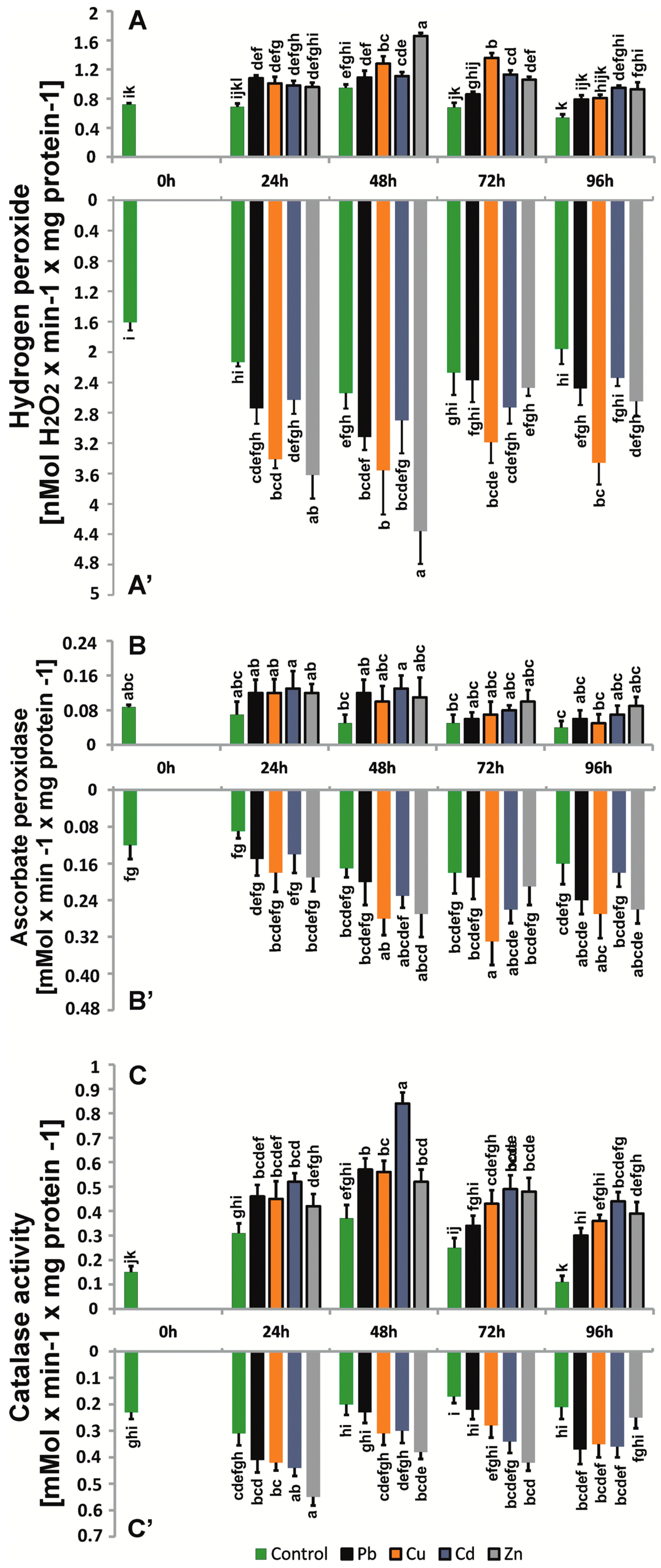
2.5. Enzyme Antioxidant Activity
2.6. Levels of Gene Transcripts
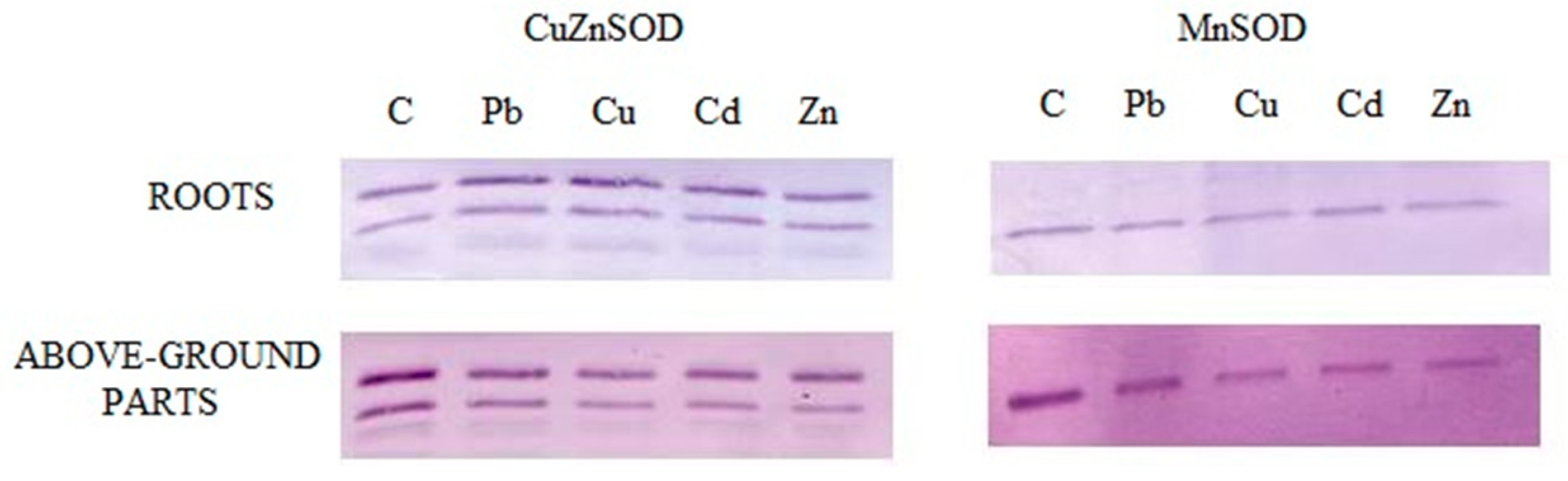
2.7. Identification of Enzyme Forms
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Phytotoxic Test
4.3. Accumulation of Trace Metals
4.4. Superoxide Anion Determination
4.5. Hydrogen Peroxide Content
4.6. In Situ Detection of Superoxide Anion and Hydrogen Peroxide
4.7. Estimation of Protein Oxidation
4.8. Determination of Antioxidant Enzyme Activities
4.9. Isolation of Total RNA and RT-PCR
4.10. Western Blot
4.11. Protein Quantification
4.12. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Z.; Zhao, Y.; Gu, L.; Wang, S.; Li, Y.; Dong, F. Accumulation and Localization of Cadmium in Potato (Solanum tuberosum) Under Different Soil Cd Levels. Bull. Environ. Contam. Toxicol. 2014, 92, 745–751. [Google Scholar] [CrossRef]
- Dalvi, A.A.; Bhalerao, S.A. Response of Plants towards Heavy Metal Toxicity: An overview of Avoidance. Tolerance and Uptake Mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- Molas, J. Changes of chloroplast ultrastructure and total chlorophyll concentration in cabbage leaves caused by excess of organic Ni (II) complexes. Environ. Exp. Bot. 2002, 47, 115–126. [Google Scholar] [CrossRef]
- Kramer, U.; Clemens, S. Functions and homeostasis of zinc, copper and nickel in plants. In Molecular Biology of Metal Homeostasis and Detoxification; Topics in Current Genetics; Springer: Berlin/Hiedelberg, Germany, 2005; Volume 14, p. 215. [Google Scholar]
- Bhardwaj, P.; Chaturvedi, A.K.; Prasad, P. Effect of Enhanced Lead and Cadmium in soil on Physiological and Biochemical attributes of Phaseolus vulgaris L. Nat. Sci. 2009, 7, 63–75. [Google Scholar]
- Bankaji, I.; Sleimi, N.; Lopez-Climent, M.F.; Perez-Clmente, R.M.; Gomez-Cadenas, A. Effects of combined abiotic stresses on grownth trace element accumulation and phytohormone regulation in two halophytic species. J. Plant Growth Regul. 2014, 33, 632–643. [Google Scholar] [CrossRef]
- Malecka, A.; Kutrowska, A.; Piechalak, A. High Peroxide Level May Be a Characteristic Trait of a Hyperaccumulator. Water Air Soil Pollut. 2015, 226, 84. [Google Scholar] [CrossRef]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef]
- Bankaji, I.; Cacador, I.; Sleimi, N. Physiological and biochemical responses of Sudaeda fruticosa to cadmium and copper stresses: Grownth, nutrient uptake, antioxidant enzymes, phytochelatin, and glutatione levels. Environ. Sci. Pollut. Res. 2015, 22, 13058–13069. [Google Scholar] [CrossRef]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Cadmium Accumulation by Plants of Brassicaceae Family and Its Connection with Their Primary and Secondary Metabolism. In The Plant Family Brassicaceae; Springer: Dordrecht, The Netherlands, 2012; pp. 71–97. [Google Scholar]
- Eapen, S.; Souza, S.F.D. Prospects of genetic engineering of plants for phytoremediation of toxic metals. Biotechnol. Adv. 2005, 23, 97–114. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metals Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Ent van der, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and matalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, D.; Hou, W. Hyperaccumulation of lead by roots, hypocotyls, and shoots of Brassica juncea. Biol. Plant. 2008, 43, 603–606. [Google Scholar] [CrossRef]
- Meyers, D.E.R.; Auchterlonie, G.J.; Webb, R.I.; Wood, B. Uptake and localisation of lead in the root system of Brassica juncea. Pollut. Environ. 2008, 153, 323–332. [Google Scholar] [CrossRef]
- Prasad, M.N.V.; Freitas, H. Metal hyperaccumulation in plants—Biodiversity prospecting for phytoremediation technology. Electron. J. Biotechnol. 2003, 6. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, P.; da Silva, J.A.T.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylgloxal and in heavy metal Chelation. J. Bot. 2012. [Google Scholar] [CrossRef]
- Malecka, A.; Piechalak, A.; Zielińska, B.; Kutrowska, A.; Tomaszewska, B. Response of the pea roots defense systems to the two-element combinations of metals (Cu, Zn, Cd, Pb). Acta Biochim. Pol. 2014, 61, 23–28. [Google Scholar] [CrossRef]
- Israr, M.; Jewell, A.; Kumar, D.; Sahi, S.V. Interactive effects of lead, copper, nickel and zinc on growth, metal uptake and antioxidative metabolism of Sesbania drummondii. J. Hazard. Mater. 2011, 186, 1520–1526. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Liu, D.; Zou, J.; Men, Q.; Zou, J.; Jiang, W. Uptake and accumulation and oxidative stress in garlic (Allium sativum L.) under lead phytotoxicity. Ecotoxicology 2009, 18, 134. [Google Scholar] [CrossRef]
- Marshner, P. Marschner’s Mineral Nutration of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Zaier, H.; Mudarra, A.; Kutcher, D.; Fernandez de la Campa, M.R.; Abdelly, C.; Sanz-Medel, A. Induced lead binding phytochelatins in Brassica juncea and Sesuvium portulacastrum investigated by orthogonal chromatography inductively coupled plasma-mass spectrometry and matrix assisted laser desorption ionization-time of flight-mass spectrometry. Anal. Chim. Acta 2010, 671, 48–54. [Google Scholar] [CrossRef]
- Yadav, P.; Kaur, R.; Kanwar, M.K.; Bhardwaj, R.; Sirhindi, G.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Ameliorative Role of Castasterone on Copper Metal Toxicity by Improving Redox Homeostsis in Brassica juncea L. J. Plant Growth Regul. 2018, 37, 575–590. [Google Scholar] [CrossRef]
- Irfan, M.; Ahmad, A.; Shamsul, H. Effect of cadmium on the growth and antioxidant enzymes in two varieties of Brassica juncea. Saudi, J. Biol. Sci. 2014, 21, 125–131. [Google Scholar] [CrossRef]
- Sinha, S.; Sinam, G.; Mishra, R.K.; Mallick, S. Metal accumulation, growth, antioxidants and oil yield of Brassica juncea L. exposed to different metals. Ecotoxicol. Environm. Saf. 2010, 73, 1352–1361. [Google Scholar] [CrossRef]
- Kutrowska, A.; Małecka, A.; Piechalak, A.; Masiakowski, W.; Hanc, A.; Barałkiewicz, D.; Andrzejewska, B.; Zbierska, J.; Tomaszewska, B. Effects of binary metal combinations on zinc, copper, cadmium and lead uptake and distribution in Brassica juncea. J. Trace Elem. Med. Biol. 2017, 44, 32–39. [Google Scholar] [CrossRef]
- Cherif, J.; Mediouni, C.; Ammar, W.B.; Jemal, F. Interactions of zinc and cadmium toxicity in their effects on growth and in antioxidative systems in tomato plants (Solanum lycopersicum). J. Environ. Sci. 2011, 23, 837–844. [Google Scholar] [CrossRef]
- Purakayastha, T.J.; Viswanath, T.; Bhadraray, S.; Chhonkar, P.K.; Adhikari, P.P.; Suribabu, K. Phytoextraction of Zinc, Copper, Nickel and Lead from a Contaminated Soil by Different Species of Brassica. Int. J. Phytoremed. 2008, 10, 61–72. [Google Scholar] [CrossRef]
- Quartacci, M.F.; Argilla, A.; Baker, A.J.M.; Navari-Izzo, F. Phytoextraction of metals from a multiply contaminated soil by Indian mustard. Chemosphere 2006, 63, 918–925. [Google Scholar] [CrossRef]
- Hanć, A.; Małecka, A.; Kutrowska, A.; Bagniewska –Zadworna, A.; Tomaszewska, B.; Barałkiewicz, D. Direct analysis of elemental biodistribution in pea seedlings by LA-ICP-MS, EDAX and confocal microscopy: Imaging and quantification. Microchem. J. 2016, 128, 305–311. [Google Scholar] [CrossRef]
- Sharma, R.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Corpas, F.J.; Rodriguez-Serrano, M.; Gomez, M.; del Rıo, L.A.; Sandalio, L.M. Differential expression and regulation of antioxidative enzymes by cadmium in pea plant. J. Plant Physiol. 2007, 164, 1346–1357. [Google Scholar] [CrossRef]
- Pena, L.B.; Tomaro, M.L.; Gallego, S.M. Effect of different metals on protease activity in sunflower cotyledons. Electron. J. Biotechnol. 2006, 9, 259–262. [Google Scholar] [CrossRef]
- Markovska, Y.K.; Gorinova, N.J.; Nedkovska, M.P.; Miteva, K.M. Cadmium-induced oxidative damage and antioxidant responses in Brassica juncea plants. Biol. Plant. 2009, 53, 151–154. [Google Scholar] [CrossRef]
- Wang, S.H.; Yang, Z.M.; Yang, H.; Lu, B.; Li, S.Q.; Lu, Y.P. Copper-induced stress and antioxidative responses in roots of Brassica juncea L. Bot. Bull. Acad. Sin. 2004, 45, 203–212. [Google Scholar]
- Malecka, A.; Piechalak, A.; Mensinger, A.; Hanc, A.; Barałkiewicz, D.; Tomaszewska, B. Antioxidative defense system in Pisum sativum roots exposed to heavy metals (Pb, Cu, Cd, Zn). Pol. J. Environ. Stud. 2012, 21, 1721–1730. [Google Scholar]
- Kanwar, M.K.; Poonam; Bhardwaj, R. Arsenuc induced modulation of antioxidative defense system and brassinosteroids in Brassica juncea L. Ecotoxicol. Environ. Saf. 2015, 115, 119–125. [Google Scholar] [CrossRef]
- Wang, S.L.; Liao, W.B.; Lu, F.Q.; Liao, B.; Shu, W.S. Hyperaccumulation of lead, zinc and cadmium in plants growing on a lead/zinc outcrop in Yunnan Province, China. Environ. Geol. 2009, 58, 471–476. [Google Scholar] [CrossRef]
- Pinto, A.P.; Alves, A.S.; Candeias, A.J.; Cardoso, A.L.; de Varennes, A.; Martins, L.L.; Mourato, M.P.; Goncales, M.L.S.; Mota, A.M. Cadmium accumulation and antioxidative defences in Brassica juncea L. Czer, Nicotiana tabacum L. and Solanum nigrum L. Int. J. Environ. Anal. Chem. 2009, 89, 661–676. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Castagna, A.; Ranieri, A.; Sanita di Toppi, L. Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. Plant Physiol. Biochem. 2012, 22, 13058–13069. [Google Scholar] [CrossRef]
- Fidalgo, F.; Freita, R.; Ferreira, R.; Pessona, A.M.; Teixeira, J. Solanum nigrum L. antioxidant defence system isozymes are regulated transcriptionally and posttranslationally in Cd-induced stress. Environ. Exp. Bot 2011, 72, 312–319. [Google Scholar] [CrossRef]
- Luo, H.; Li, H.; Zhang, X.; Fu, J. Antioxidant responses and gene expression in perennial ryegrass (Lollium perenne L.) under cadmium stress. Ecotoxicology 2011, 20, 770–778. [Google Scholar] [CrossRef]
- Bernard, F.; Brulle, F.; Dumez, S.; Lemiere, S.; Platel, A.; Nesslany, F.; Cuny, D.; Deramz, A.; Vandenbulcke, F. Antioxidant responses of Annelids, Brassicaceae and Fabaceae to pollutants: A review. Ecotoxicol. Environ. Saf. 2015, 114, 273–303. [Google Scholar] [CrossRef]
- Wilkins, D.A. A technique for the measurement of lead tolerance in plants. Nature 1957, 180, 37–38. [Google Scholar] [CrossRef]
- Hanć, A.; Olszewska, H.; Barałkiewicz, D. Quantitative analysis of elements migration in human teeth with and without filling using LA-ICP-MS. Microchem. J. 2013, 110, 61–69. [Google Scholar] [CrossRef]
- Doke, N. Invovement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Mol. Plant Pathol. 1983, 23, 345–355. [Google Scholar] [CrossRef]
- Patterson, B.D.; Macrae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium(IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase in vitro. In Methods of Enzymatic Analyses; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1983; Volume 3, pp. 273–282. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantitiesof protein utilizing the principle of protein- dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Małecka, A.; Konkolewska, A.; Hanć, A.; Barałkiewicz, D.; Ciszewska, L.; Ratajczak, E.; Staszak, A.M.; Kmita, H.; Jarmuszkiewicz, W. Insight into the Phytoremediation Capability of Brassica juncea (v. Malopolska): Metal Accumulation and Antioxidant Enzyme Activity. Int. J. Mol. Sci. 2019, 20, 4355. https://doi.org/10.3390/ijms20184355
Małecka A, Konkolewska A, Hanć A, Barałkiewicz D, Ciszewska L, Ratajczak E, Staszak AM, Kmita H, Jarmuszkiewicz W. Insight into the Phytoremediation Capability of Brassica juncea (v. Malopolska): Metal Accumulation and Antioxidant Enzyme Activity. International Journal of Molecular Sciences. 2019; 20(18):4355. https://doi.org/10.3390/ijms20184355
Chicago/Turabian StyleMałecka, Arleta, Agnieszka Konkolewska, Anetta Hanć, Danuta Barałkiewicz, Liliana Ciszewska, Ewelina Ratajczak, Aleksandra Maria Staszak, Hanna Kmita, and Wiesława Jarmuszkiewicz. 2019. "Insight into the Phytoremediation Capability of Brassica juncea (v. Malopolska): Metal Accumulation and Antioxidant Enzyme Activity" International Journal of Molecular Sciences 20, no. 18: 4355. https://doi.org/10.3390/ijms20184355
APA StyleMałecka, A., Konkolewska, A., Hanć, A., Barałkiewicz, D., Ciszewska, L., Ratajczak, E., Staszak, A. M., Kmita, H., & Jarmuszkiewicz, W. (2019). Insight into the Phytoremediation Capability of Brassica juncea (v. Malopolska): Metal Accumulation and Antioxidant Enzyme Activity. International Journal of Molecular Sciences, 20(18), 4355. https://doi.org/10.3390/ijms20184355