Wnt/β-Catenin Signaling: The Culprit in Pancreatic Carcinogenesis and Therapeutic Resistance

 ,
, 
Abstract
1. Introduction
1.1. Pancreatic Cancer
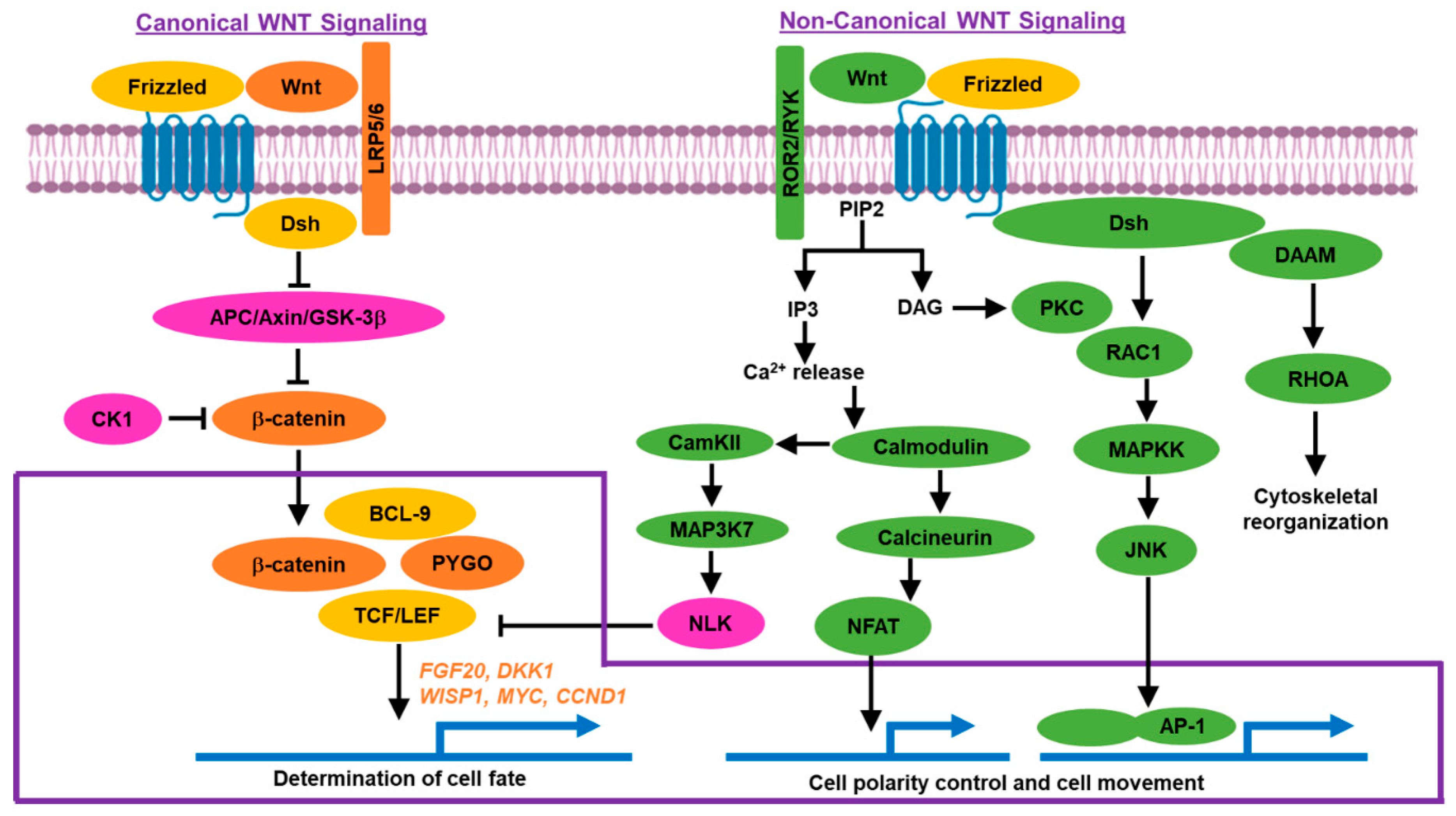
1.2. Wnt/β-Catenin Signaling Pathway
1.2.1. Canonical Pathway
1.2.2. Non-Canonical Pathway
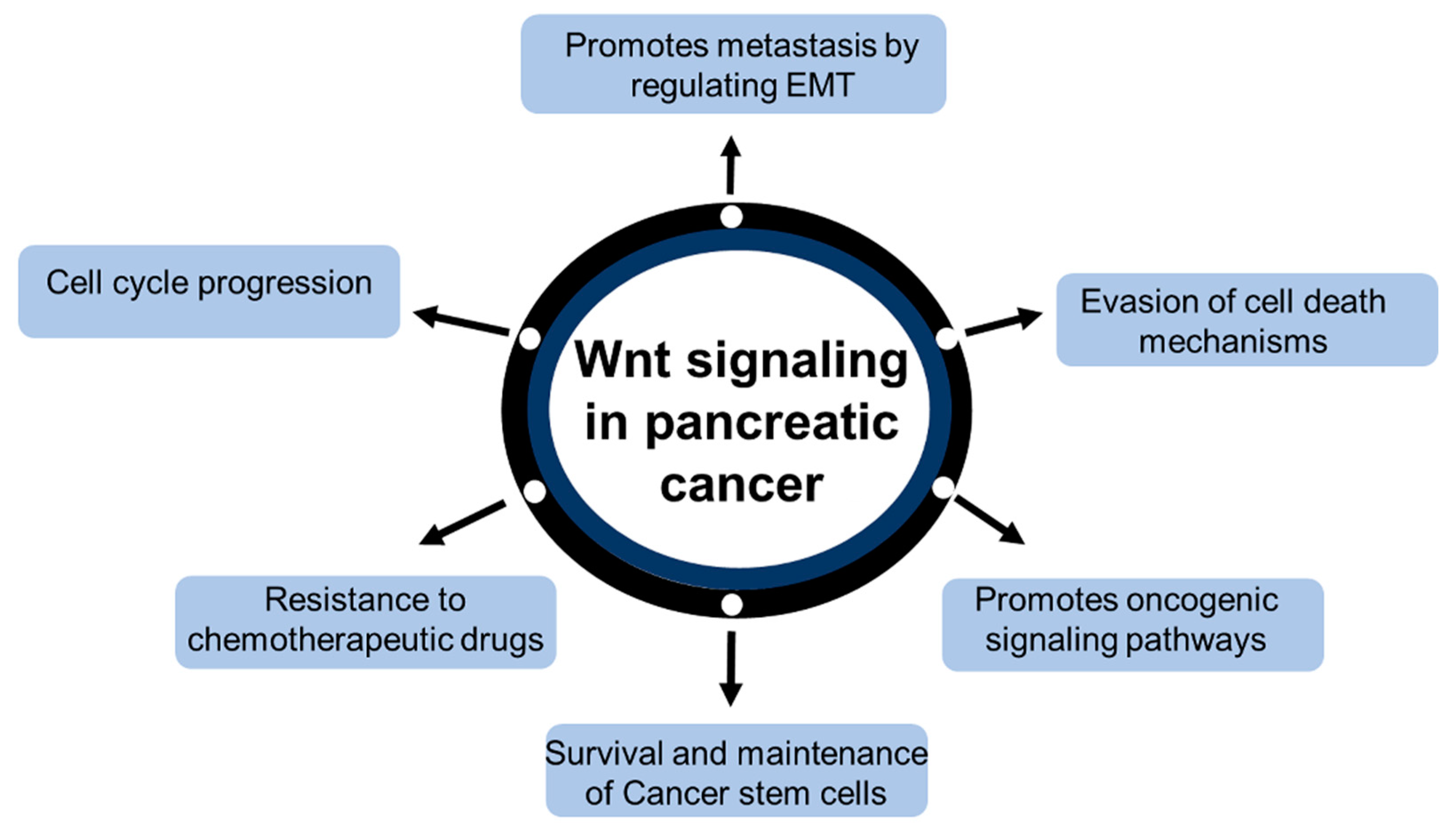
2. Wnt/β-Catenin Signaling in Pancreatic Cancer
3. Inhibitors for Wnt/β-Catenin Signaling Pathway
3.1. Porcupine Inhibitors
3.2. Antibodies Against Wnt Family Proteins
3.2.1. OMP-18R5 (Vantictumab)
3.2.2. OMP-54 F28 (Ipafricept)
3.2.3. Other Antibodies
3.3. β-Catenin Inhibitors
3.3.1. PRI-724
3.3.2. Wnt5a Mimetics
4. Wnt/β-Catenin Signaling in Pancreatic Cancer Drug Resistance
5. Challenges of Targeting the Wnt Pathway
6. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PDAC | Pancreatic ductal adenocarcinoma |
| EMT | Epithelial mesenchymal transition |
| LRP | Lipoprotein receptor related protein |
| DKK1 | Dickkopf 1 |
| Dsh | Disheveled |
| CBP | CREB binding protein |
| PCP | Planar cell polarity |
| ODC | Ornithine decarboxylase |
| OMP | 18R5 Vantictumab free survival |
| PFS | Median progression |
| OMP | 54 F28 Ipafricept |
| CSCs | Cancer stem cells |
References
- Kleeff, J.; Korc, M.; Apte, M.; La Vecchia, C.; Johnson, C.D.; Biankin, A.V.; Neale, R.E.; Tempero, M.; Tuveson, D.A.; Hruban, R.H.; et al. Pancreatic cancer. Nat. Rev. Dis. Primers 2016, 2, 16022. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.; Makena, M.; Bhowmick, K.; Pandey, M. Advancement of NF-κB Signaling Pathway: A Novel Target in Pancreatic Cancer. Int. J. Mol. Sci. 2018, 19, 3890. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.M.; Turk, T.; Al-Husseini, M.J.; Abdel-Rahman, O. Trends in pancreatic adenocarcinoma incidence and mortality in the United States in the last four decades; a SEER-based study. BMC Cancer 2018, 18, 688. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Ilic, M.; Ilic, I. Epidemiology of pancreatic cancer. World J. Gastroenterol. 2016, 22, 9694–9705. [Google Scholar] [CrossRef] [PubMed]
- Neoptolemos, J.P.; Kleeff, J.; Michl, P.; Costello, E.; Greenhalf, W.; Palmer, D.H. Therapeutic developments in pancreatic cancer: Current and future perspectives. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Morrison, A.H.; Byrne, K.T.; Vonderheide, R.H. Immunotherapy and prevention of pancreatic cancer. Trends Cancer 2018, 4, 418–428. [Google Scholar] [CrossRef]
- Pelosi, E.; Castelli, G.; Testa, U. Pancreatic cancer: Molecular characterization, clonal evolution and cancer stem cells. Biomedicines 2017, 5, 65. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Makena, M.R.; Ranjan, A.; Thirumala, V.; Reddy, A. Cancer stem cells: Road to therapeutic resistance and strategies to overcome resistance. Biochim. Biophys Acta (BBA)-Mol. Basis Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kahn, M. Can we safely target the WNT pathway? Nat. Rev. Drug Discov. 2014, 13, 513. [Google Scholar] [CrossRef] [PubMed]
- Bastakoty, D.; Young, P.P. Wnt/β-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. FASEB J. 2016, 30, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.J.; Ro, E.J.; Choi, K.Y. Interaction between Wnt/β-catenin and RAS-ERK pathways and an anti-cancer strategy via degradations of β-catenin and RAS by targeting the Wnt/β-catenin pathway. NPJ Precis. Oncol. 2018, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Gómez-Orte, E.; Sáenz-Narciso, B.; Moreno, S.; Cabello, J. Multiple functions of the noncanonical Wnt pathway. Trends Genet. 2013, 29, 545–553. [Google Scholar]
- Dang, D.K.; Makena, M.R.; Llongueras, J.P.; Prasad, H.; Ko, M.; Bandral, M.; Rao, R. A Ca2+-ATPase Regulates E-cadherin Biogenesis and Epithelial–Mesenchymal Transition in Breast Cancer Cells. Mol. Cancer Res. 2019. [Google Scholar] [CrossRef]
- Makena, M.R.; Dang, D.K.; Ko, M.; Bandral, M.; Rao, R. Secretory Pathway Calcium ATPase-2 (SPCA2) Regulates Metastasis by Suppressing Mesenchymal Markers in Triple Negative Breast Cancer Cell Lines. Am. Assoc. Cancer Res. 2019, 1125. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, Z.; Tang, Y.; Chen, X.; Zhou, J. Identification of potential target genes in pancreatic ductal adenocarcinoma by bioinformatics analysis. Oncol. Lett. 2018, 16, 2453–2461. [Google Scholar] [CrossRef]
- Modi, S.; Kir, D.; Banerjee, S.; Saluja, A. Control of apoptosis in treatment and biology of pancreatic cancer. J. Cell. Biochem. 2016, 117, 279–288. [Google Scholar] [CrossRef]
- Jiang, H.; Li, Q.; He, C.; Li, F.; Sheng, H.; Shen, X.; Zhang, X.; Zhu, S.; Chen, H.; Chen, X. Activation of the Wnt pathway through Wnt2 promotes metastasis in pancreatic cancer. Am. J. Cancer Res. 2014, 4, 537. [Google Scholar] [PubMed]
- Bo, H.; Gao, L.; Chen, Y.; Zhang, J.; Zhu, M. Upregulation of the expression of Wnt5a promotes the proliferation of pancreatic cancer cells in vitro and in a nude mouse model. Mol. Med. Rep. 2016, 13, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.J.; Jiang, Y.S.; He, R.Z.; Tao, L.Y.; Yang, M.W.; Fu, X.L.; Yang, J.Y.; Zhu, K. High expression of WNT7A predicts poor prognosis and promote tumor metastasis in pancreatic ductal adenocarcinoma. Sci. Rep. 2018, 8, 15792. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Wang, Z.; Fu, B.; Pan, F.; Yachida, S.; Dhara, M.; Albesiano, E.; Li, L.; Naito, Y.; Vilardell, F. GATA6 activates Wnt signaling in pancreatic cancer by negatively regulating the Wnt antagonist Dickkopf-1. PLoS ONE 2011, 6, e22129. [Google Scholar] [CrossRef] [PubMed]
- Chartier, C.; Raval, J.; Axelrod, F.; Bond, C.; Cain, J.; Dee-Hoskins, C.; Ma, S.; Fischer, M.M.; Shah, J.; Wei, J. Therapeutic targeting of tumor-derived R-spondin attenuates β-catenin signaling and tumorigenesis in multiple cancer types. Cancer Res. 2016, 76, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Ilmer, M.; Boiles, A.R.; Regel, I.; Yokoi, K.; Michalski, C.W.; Wistuba, I.I.; Rodriguez, J.; Alt, E.; Vykoukal, J. RSPO2 enhances canonical Wnt signaling to confer stemness-associated traits to susceptible pancreatic cancer cells. Cancer Res. 2015, 75, 1883–1896. [Google Scholar] [CrossRef]
- He, Y.M.; Xiao, Y.S.; Wei, L.; Zhang, J.Q.; Peng, C.H. CUL4B promotes metastasis and proliferation in pancreatic cancer cells by inducing epithelial-mesenchymal transition via the Wnt/β-catenin signaling pathway. J. Cell. Biochem. 2018, 119, 5308–5323. [Google Scholar] [CrossRef]
- Xu, W.; Wang, Z.; Zhang, W.; Qian, K.; Li, H.; Kong, D.; Li, Y.; Tang, Y. Mutated K-ras activates CDK8 to stimulate the epithelial-to-mesenchymal transition in pancreatic cancer in part via the Wnt/β-catenin signaling pathway. Cancer Lett. 2015, 356, 613–627. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Ji, D.; Kong, X.; Liu, D.; Zhao, Z.; Yan, J.; Chen, S. Expression of Rab5a correlates with tumor progression in pancreatic carcinoma. Virchows Arch. 2017, 470, 527–536. [Google Scholar] [CrossRef]
- Yu, L.; Li, X.; Li, H.; Chen, H.; Liu, H. Rab11a sustains GSK3β/WNT/β-catenin signaling to enhance cancer progression in pancreatic cancer. Tumor Biol. 2016, 37, 13821–13829. [Google Scholar] [CrossRef]
- Liu, X.; Caffrey, T.; Steele, M.; Mohr, A.; Singh, P.; Radhakrishnan, P.; Kelly, D.; Wen, Y.; Hollingsworth, M. MUC1 regulates cyclin D1 gene expression through p120 catenin and β-catenin. Oncogenesis 2014, 3, e107. [Google Scholar] [CrossRef] [PubMed]
- Zhi, X.; Tao, J.; Xie, K.; Zhu, Y.; Li, Z.; Tang, J.; Wang, W.; Xu, H.; Zhang, J.; Xu, Z. MUC4-induced nuclear translocation of β-catenin: A novel mechanism for growth, metastasis and angiogenesis in pancreatic cancer. Cancer Lett. 2014, 346, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Manegold, P.; Lai, K.K.; Wu, Y.; Teo, J.L.; Lenz, H.J.; Genyk, Y.S.; Pandol, S.J.; Wu, K.; Lin, D.P.; Chen, Y. Differentiation Therapy Targeting the β-Catenin/CBP Interaction in Pancreatic Cancer. Cancers 2018, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Gatla, H.R.; Zou, Y.; Uddin, M.M.; Singha, B.; Bu, P.; Vancura, A.; Vancurova, I. Histone deacetylase (HDAC) inhibition induces IκB kinase (IKK)-dependent interleukin-8/CXCL8 expression in ovarian cancer cells. J. Biol. Chem. 2017, 292, 5043–5054. [Google Scholar] [CrossRef] [PubMed]
- Gatla, H.R.; Zou, Y.; Uddin, M.M.; Vancurova, I. Epigenetic regulation of interleukin-8 expression by class I HDAC and CBP in ovarian cancer cells. Oncotarget 2017, 8, 70798. [Google Scholar] [CrossRef] [PubMed]
- Arensman, M.D.; Telesca, D.; Lay, A.R.; Kershaw, K.M.; Wu, N.; Donahue, T.R.; Dawson, D.W. The CREB binding protein inhibitor ICG-001 suppresses pancreatic cancer growth. Mol. Cancer Ther. 2014, 13, 2303–2314. [Google Scholar] [CrossRef]
- Zhang, Q.; Lou, Y.; Zhang, J.; Fu, Q.; Wei, T.; Sun, X.; Chen, Q.; Yang, J.; Bai, X.; Liang, T. Hypoxia-inducible factor-2α promotes tumor progression and has crosstalk with Wnt/β-catenin signaling in pancreatic cancer. Mol. Cancer 2017, 16, 119. [Google Scholar] [CrossRef]
- Song, M.M.; Makena, M.R.; Hindle, A.; Koneru, B.; Nguyen, T.H.; Verlekar, D.U.; Cho, H.; Maurer, B.J.; Kang, M.H.; Reynolds, C.P. Cytotoxicity and molecular activity of fenretinide and metabolites in T-cell lymphoid malignancy, neuroblastoma, and ovarian cancer cell lines in physiological hypoxia. Anti-Cancer Drugs 2019, 30, 117–127. [Google Scholar] [CrossRef]
- Criscimanna, A.; Duan, D.; Rhodes, J.A.; Fendrich, V.; Wickline, E.D.; Hartman, D.J.; Monga, S.P.; Lotze, M.; Gittes, G.K.; Fong, G.H. PanIN-specific regulation of Wnt signaling by HIF2. Cancer Res. 2013, 73, 4781–4790. [Google Scholar] [CrossRef]
- Jiang, J.X.; Sun, C.Y.; Tian, S.; Yu, C.; Chen, M.Y.; Zhang, H. Tumor suppressor Fbxw7 antagonizes WNT signaling by targeting β-catenin for degradation in pancreatic cancer. Tumor Biol. 2016, 37, 13893–13902. [Google Scholar]
- Quan, M.; Cui, J.; Xia, T.; Jia, Z.; Xie, D.; Wei, D.; Huang, S.; Huang, Q.; Zheng, S.; Xie, K. Merlin/NF2 suppresses pancreatic tumor growth and metastasis by attenuating the FOXM1-mediated Wnt/beta-catenin signaling. Cancer Res. 2015, 75, 4778–4789. [Google Scholar] [CrossRef]
- Fakhar, M.; Gul, M.; Rashid, S. Antagonistic role of Klotho-derived peptides dynamics in the pancreatic cancer treatment through obstructing WNT-1 and Frizzled binding. Biophys. Chem. 2018, 240, 107–117. [Google Scholar] [CrossRef]
- Qian, Y.; Li, J.; Xia, S. ZNF281 Promotes Growth and Invasion of Pancreatic Cancer Cells by Activating Wnt/β-Catenin Signaling. Dig. Dis. Sci. 2017, 62, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Douchi, D.; Ohtsuka, H.; Ariake, K.; Masuda, K.; Kawasaki, S.; Kawaguchi, K.; Fukase, K.; Oikawa, M.; Motoi, F.; Naitoh, T. Silencing of LRRFIP1 reverses the epithelial–mesenchymal transition via inhibition of the Wnt/β-catenin signaling pathway. Cancer Lett. 2015, 365, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Chetty, R. Rnf43. J. Clin. Pathol. 2018, 71, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Song, K.S.; Jing, K.; Kim, J.S.; Yun, E.J.; Shin, S.; Seo, K.S.; Park, J.H.; Heo, J.Y.; Kang, J.X.; Suh, K.S. Omega-3-polyunsaturated fatty acids suppress pancreatic cancer cell growth in vitro and in vivo via downregulation of Wnt/Beta-catenin signaling. Pancreatology 2011, 11, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, W.; Li, X.; Meng, W.; Zhang, L.; Zhu, X.; Zhu, K.; Bai, Z.; Yan, J.; Liu, T. 5-Azacytidine suppresses the proliferation of pancreatic cancer cells by inhibiting the Wnt/β-catenin signaling pathway. Genet. Mol. Res. 2014, 13, 5064–5072. [Google Scholar] [CrossRef]
- Arensman, M.D.; Nguyen, P.; Kershaw, K.M.; Lay, A.R.; Ostertag-Hill, C.A.; Sherman, M.H.; Downes, M.; Liddle, C.; Evans, R.M.; Dawson, D.W. Calcipotriol targets LRP6 to inhibit Wnt signaling in pancreatic cancer. Mol. Cancer Res. 2015, 13, 1509–1519. [Google Scholar] [CrossRef]
- Jung, D.B.; Yun, M.; Kim, E.O.; Kim, J.; Kim, B.; Jung, J.H.; Wang, E.; Mukhopadhyay, D.; Hammond, E.; Dredge, K. The heparan sulfate mimetic PG545 interferes with Wnt/β-catenin signaling and significantly suppresses pancreatic tumorigenesis alone and in combination with gemcitabine. Oncotarget 2015, 6, 4992. [Google Scholar] [CrossRef]
- Xu, Z.; Li, C.; Qu, H.; Li, H.; Gu, Q.; Xu, J. MicroRNA-195 inhibits the proliferation and invasion of pancreatic cancer cells by targeting the fatty acid synthase/Wnt signaling pathway. Tumor Biol. 2017, 39, 1010428317711324. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, Q.; Zhou, M.; Yang, W.; Shi, H.; Shan, Y.; Zhang, Q.; Yu, F. Restoration of miRNA-148a in pancreatic cancer reduces invasion and metastasis by inhibiting the Wnt/β-catenin signaling pathway via downregulating maternally expressed gene-3. Exp. Ther. Med. 2019, 17, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Shi, C.; Li, T.; Kuang, T. MicroRNA-454 shows anti-angiogenic and anti-metastatic activity in pancreatic ductal adenocarcinoma by targeting LRP6. Am. J. Cancer Res. 2017, 7, 139. [Google Scholar] [PubMed]
- Yang, H.W.; Liu, G.H.; Liu, Y.Q.; Zhao, H.C.; Yang, Z.; Zhao, C.L.; Zhang, X.F.; Ye, H. Over-expression of microRNA-940 promotes cell proliferation by targeting GSK3β and sFRP1 in human pancreatic carcinoma. Biomed. Pharmacother. 2016, 83, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liang, J.; Bai, L. MicroRNA-449a functions as a tumor suppressor in pancreatic cancer by the epigenetic regulation of ATDC expression. Biomed. Pharmacother. 2018, 103, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Q.; Chen, S.; Gu, J.N.; Zhu, Y.; Zhan, Q.; Cheng, D.F.; Chen, H.; Deng, X.X.; Shen, B.Y.; Peng, C.H. MicroRNA-300 promotes apoptosis and inhibits proliferation, migration, invasion and epithelial-mesenchymal transition via the Wnt/β-catenin signaling pathway by targeting CUL4B in pancreatic cancer cells. J. Cell. Biochem. 2018, 119, 1027–1040. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, Y.; Gou, S.; Xiong, J.; Wu, H.; Wang, C.; Yan, H.; Liu, T. MiR-744 increases tumorigenicity of pancreatic cancer by activating Wnt/β-catenin pathway. Oncotarget 2015, 6, 37557. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.C.; Ma, J.; Zhang, J.; Wang, J.C. Long non-coding RNA LINC01133 silencing exerts antioncogenic effect in pancreatic cancer through the methylation of DKK1 promoter and the activation of Wnt signaling pathway. Cancer Biol. Ther. 2019, 20, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhu, Q.; Yang, W.; Shan, Y.; Yu, Z.; Zhang, Q.; Wu, H. LncRNA H19/miR-194/PFTK1 axis modulates the cell proliferation and migration of pancreatic cancer. J. Cell. Biochem. 2019, 120, 3874–3886. [Google Scholar] [CrossRef] [PubMed]
- Saukkonen, K.; Hagström, J.; Mustonen, H.; Juuti, A.; Nordling, S.; Kallio, P.; Alitalo, K.; Seppänen, H.; Haglund, C. PROX1 and β-catenin are prognostic markers in pancreatic ductal adenocarcinoma. BMC Cancer 2016, 16, 472. [Google Scholar] [CrossRef]
- Mohammed, A.; Janakiram, N.B.; Madka, V.; Ritchie, R.L.; Brewer, M.; Biddick, L.; Patlolla, J.M.R.; Sadeghi, M.; Lightfoot, S.; Steele, V.E. Eflornithine (DFMO) prevents progression of pancreatic cancer by modulating ornithine decarboxylase signaling. Cancer Prev. Res. 2014, 7, 1198–1209. [Google Scholar] [CrossRef]
- Makena, M.R.; Cho, H.E.; Nguyen, T.H.; Koneru, B.; Verlekar, D.U.; Hindle, A.; Kang, M.H.; Reynolds, C.P. Cytotoxic activity of difluoromethylornithine compared with fenretinide in neuroblastoma cell lines. Pediatric Blood Cancer 2018, 65, e27447. [Google Scholar] [CrossRef] [PubMed]
- Proffitt, K.D.; Virshup, D.M. Precise regulation of porcupine activity is required for physiological Wnt signaling. J. Biol. Chem. 2012, 287, 34167–34178. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hao, H.-X.; Growney, J.D.; Woolfenden, S.; Bottiglio, C.; Ng, N.; Lu, B.; Hsieh, M.H.; Bagdasarian, L.; Meyer, R. Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci. USA 2013, 110, 12649–12654. [Google Scholar] [CrossRef] [PubMed]
- Madan, B.; Ke, Z.; Harmston, N.; Ho, S.Y.; Frois, A.; Alam, J.; Jeyaraj, D.A.; Pendharkar, V.; Ghosh, K.; Virshup, I.H. Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 2016, 35, 2197. [Google Scholar] [CrossRef] [PubMed]
- Madan, B.; Harmston, N.; Nallan, G.; Montoya, A.; Faull, P.; Petretto, E.; Virshup, D.M. Temporal dynamics of Wnt-dependent transcriptome reveal an oncogenic Wnt/MYC/ribosome axis. J. Clin. Investig. 2018, 128, 5620–5633. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.C.; Rosen, L.S.; Chugh, R.; Goldman, J.W.; Xu, L.; Kapoun, A.; Brachmann, R.K.; Dupont, J.; Stagg, R.J.; Tolcher, A.W. First-in-human evaluation of the human monoclonal antibody vantictumab (OMP-18R5; anti-Frizzled) targeting the WNT pathway in a phase I study for patients with advanced solid tumors. J. Clin. Oncol. 2013, 31 (Suppl. 15), 2540. [Google Scholar]
- Gurney, A.; Axelrod, F.; Bond, C.J.; Cain, J.; Chartier, C.; Donigan, L.; Fischer, M.; Chaudhari, A.; Ji, M.; Kapoun, A.M. Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 11717–11722. [Google Scholar] [CrossRef]
- Messersmith, W.; Cohen, S.; Shahda, S.; Lenz, H.J.; Weekes, C.; Dotan, E.; Denlinger, C.; O’Neil, B.; Kapoun, A.; Zhang, C. Phase 1b study of WNT inhibitor vantictumab (VAN, human monoclonal antibody) with nab-paclitaxel (Nab-P) and gemcitabine (G) in patients (pts) with previously untreated stage IV pancreatic cancer (PC). Ann. Oncol. 2016, 27 (Suppl. 6). [Google Scholar] [CrossRef]
- Le, P.N.; McDermott, J.D.; Jimeno, A. Targeting the Wnt pathway in human cancers: Therapeutic targeting with a focus on OMP-54F28. Pharmacol. Ther. 2015, 146, 1–11. [Google Scholar] [CrossRef]
- Fischer, M.M.; Cancilla, B.; Yeung, V.P.; Cattaruzza, F.; Chartier, C.; Murriel, C.L.; Cain, J.; Tam, R.; Cheng, C.Y.; Evans, J.W. WNT antagonists exhibit unique combinatorial antitumor activity with taxanes by potentiating mitotic cell death. Sci. Adv. 2017, 3, e1700090. [Google Scholar] [CrossRef]
- Weekes, C.; Berlin, J.; Lenz, H.J.; O’Neil, B.; Messersmith, W.; Cohen, S.; Dendinger, C.; Shahda, S.; Kapoun, A.; Zhang, C. Phase 1b study of WNT inhibitor ipafricept (IPA, decoy receptor for WNT ligands) with nab-paclitaxel (Nab-P) and gemcitabine (G) in patients (pts) with previously untreated stage IV pancreatic cancer (PC). Ann. Oncol. 2016, 27 (Suppl. 6). [Google Scholar] [CrossRef]
- Dotan, E.; Cardin, D.B.; Lenz, H.J.; Messersmith, W.A.; O’Neil, B.; Cohen, S.J.; Denlinger, C.S.; Shahda, S.; Kapoun, A.M.; Brachmann, R.K. Phase Ib study of WNT inhibitor ipafricept (IPA) with nab-paclitaxel (Nab-P) and gemcitabine (G) in patients (pts) with previously untreated stage IV pancreatic cancer (mPC). Am. Soc. Clin. Oncol. 2019. [Google Scholar] [CrossRef]
- Giraudet, A.L.; Cassier, P.A.; Iwao-Fukukawa, C.; Garin, G.; Badel, J.N.; Kryza, D.; Chabaud, S.; Gilles-Afchain, L.; Clapisson, G.; Desuzinges, C. A first-in-human study investigating biodistribution, safety and recommended dose of a new radiolabeled MAb targeting FZD10 in metastatic synovial sarcoma patients. BMC Cancer 2018, 18, 646. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yee, A.; Sirard, C.; Landau, S.; Raje, N.; Mahmood, U. Sodium fluoride PET imaging as a quantitative pharmacodynamic biomarker for bone homeostasis during anti-DKK1 therapy for multiple myeloma. Blood Cancer J. 2017, 7, e615. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.J.; Kahn, M. Safely targeting cancer stem cells via selective catenin coactivator antagonism. Cancer Sci. 2014, 105, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.H.; Chiorean, E.G.; Kwak, E.L.; Lenz, H.J.; Nadler, P.I.; Wood, D.L.; Fujimori, M.; Inada, T.; Kouji, H.; McWilliams, R.R. Final results of a phase Ib dose-escalation study of PRI-724, a CBP/beta-catenin modulator, plus gemcitabine (GEM) in patients with advanced pancreatic adenocarcinoma (APC) as second-line therapy after FOLFIRINOX or FOLFOX. Am. Soc. Clin. Oncol. 2016. [Google Scholar] [CrossRef]
- Grigson, E.R.; Ozerova, M.; Pisklakova, A.; Liu, H.; Sullivan, D.M.; Nefedova, Y. Canonical Wnt pathway inhibitor ICG-001 induces cytotoxicity of multiple myeloma cells in Wnt-independent manner. PLoS ONE 2015, 10, e0117693. [Google Scholar] [CrossRef] [PubMed]
- Ripka, S.; König, A.; Buchholz, M.; Wagner, M.; Sipos, B.; Klöppel, G.; Downward, J.; Gress, T.; Michl, P. WNT5A—target of CUTL1 and potent modulator of tumor cell migration and invasion in pancreatic cancer. Carcinogenesis 2007, 28, 1178–1187. [Google Scholar] [CrossRef]
- Canesin, G.; Evans-Axelsson, S.; Hellsten, R.; Krzyzanowska, A.; Prasad, C.P.; Bjartell, A.; Andersson, T. Treatment with the WNT5A-mimicking peptide Foxy-5 effectively reduces the metastatic spread of WNT5A-low prostate cancer cells in an orthotopic mouse model. PLoS ONE 2017, 12, e0184418. [Google Scholar] [CrossRef]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J. Wnt/β-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef]
- Vankelecom, H.; Van Delm, W.; Gremeaux, L.; Wouters, J.; Allemeersch, J.; Govaere, O.; Roskams, T.; Topal, B. Human pancreatic cancer contains a side population expressing cancer stem cell-associated and prognostic genes. PLoS ONE 2013, 8, e73968. [Google Scholar]
- Chikazawa, N.; Tanaka, H.; Tasaka, T.; Nakamura, M.; Tanaka, M.; Onishi, H.; Katano, M. Inhibition of Wnt signaling pathway decreases chemotherapy-resistant side-population colon cancer cells. Anticancer. Res. 2010, 30, 2041–2048. [Google Scholar] [PubMed]
- Niess, H.; Camaj, P.; Renner, A.; Ischenko, I.; Zhao, Y.; Krebs, S.; Mysliwietz, J.; Jäckel, C.; Nelson, P.J.; Blum, H. Side population cells of pancreatic cancer show characteristics of cancer stem cells responsible for resistance and metastasis. Target. Oncol. 2015, 10, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Quint, K.; Tonigold, M.; Di Fazio, P.; Montalbano, R.; Lingelbach, S.; Rückert, F.; Alinger, B.; Ocker, M.; Neureiter, D. Pancreatic cancer cells surviving gemcitabine treatment express markers of stem cell differentiation and epithelial-mesenchymal transition. Int. J. Oncol. 2012, 41, 2093–2102. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Xie, J. Promising molecular mechanisms responsible for gemcitabine resistance in cancer. Genes Dis. 2015, 2, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Griesmann, H.; Ripka, S.; Pralle, M.; Ellenrieder, V.; Baumgart, S.; Buchholz, M.; Pilarsky, C.; Aust, D.; Gress, T.M.; Michl, P. WNT5A-NFAT signaling mediates resistance to apoptosis in pancreatic cancer. Neoplasia 2013, 15, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Sun, H.H.; Li, N.; Li, H.Y.; Li, X.; Li, Q.; Shen, X.H. WNT5A modulates cell cycle progression and contributes to the chemoresistance in pancreatic cancer cells. Hepatobiliary Pancreat. Dis. Int. 2014, 13, 529–538. [Google Scholar] [CrossRef]
- Nagano, H.; Tomimaru, Y.; Eguchi, H.; Hama, N.; Wada, H.; Kawamoto, K.; Kobayashi, S.; Mori, M.; Doki, Y. MicroRNA-29a induces resistance to gemcitabine through the Wnt/β-catenin signaling pathway in pancreatic cancer cells. Int. J. Oncol. 2013, 43, 1066–1072. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Z.; Li, Y.Y.; Yu, B.H.; Zhang, F.; Li, H.Y. MiR-33a suppresses the nuclear translocation of β-catenin to enhance gemcitabine sensitivity in human pancreatic cancer cells. Tumor Biol. 2015, 36, 9395–9403. [Google Scholar] [CrossRef]
- Cao, J.; Ma, J.; Sun, L.; Li, J.; Qin, T.; Zhou, C.; Cheng, L.; Chen, K.; Qian, W.; Duan, W. Targeting glypican-4 overcomes 5-FU resistance and attenuates stem cell–like properties via suppression of Wnt/β-catenin pathway in pancreatic cancer cells. J. Cell. Biochem. 2018, 119, 9498–9512. [Google Scholar] [CrossRef]
- Kang, M.H.; Wang, J.; Makena, M.R.; Lee, J.S.; Paz, N.; Hall, C.P.; Song, M.M.; Calderon, R.I.; Cruz, R.E.; Hindle, A. Activity of MM-398, nanoliposomal irinotecan (nal-IRI), in Ewing’s family tumor xenografts is associated with high exposure of tumor to drug and high SLFN11 expression. Clin. Cancer Res. 2015, 21, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Wang-Gillam, A.; Hubner, R.A.; Siveke, J.T.; Von Hoff, D.D.; Belanger, B.; de Jong, F.A.; Mirakhur, B.; Chen, L.T. NAPOLI-1 phase 3 study of liposomal irinotecan in metastatic pancreatic cancer: Final overall survival analysis and characteristics of long-term survivors. Eur. J. Cancer 2019, 108, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, J.; Ko, A.H. MM-398 (nanoliposomal irinotecan): Emergence of a novel therapy for the treatment of advanced pancreatic cancer. Future Oncol. 2016, 12, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Skrypek, N.; Vasseur, R.; Vincent, A.; Duchêne, B.; Van Seuningen, I.; Jonckheere, N. The oncogenic receptor ErbB2 modulates gemcitabine and irinotecan/SN-38 chemoresistance of human pancreatic cancer cells via hCNT1 transporter and multidrug-resistance associated protein MRP-2. Oncotarget 2015, 6, 10853. [Google Scholar] [CrossRef] [PubMed]
- Schade, B.; Lesurf, R.; Sanguin-Gendreau, V.; Bui, T.; Deblois, G.; O’Toole, S.A.; Millar, E.K.; Zardawi, S.J.; Lopez-Knowles, E.; Sutherland, R.L. β-Catenin signaling is a critical event in ErbB2-mediated mammary tumor progression. Cancer Res. 2013, 73, 4474–4487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, M.; Huang, H.; Li, L.; Yang, J.; Feng, L.; Gou, J.; Jiang, M.; Peng, L.; Chen, L. Dishevelled1-3 contribute to multidrug resistance in colorectal cancer via activating Wnt/β-catenin signaling. Oncotarget 2017, 8, 115803. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Balibrea, E.; Martínez-Cardús, A.; Ginés, A.; de Porras, V.R.; Moutinho, C.; Layos, L.; Manzano, J.L.; Bugés, C.; Bystrup, S.; Esteller, M. Tumor-related molecular mechanisms of oxaliplatin resistance. Mol. Cancer Ther. 2015, 14, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, A.C.; Tirona, R.G.; Kim, R.B. Pharmacogenetics of ATP-binding Cassette Transporters in Cancer and Chemotherapy1. Mol. Cancer Ther. 2003, 2, 685–698. [Google Scholar]
- Salim, H.; Zong, D.; Hååg, P.; Novak, M.; Mörk, B.; Lewensohn, R.; Lundholm, L.; Viktorsson, K. DKK1 is a potential novel mediator of cisplatin-refractoriness in non-small cell lung cancer cell lines. BMC Cancer 2015, 15, 628. [Google Scholar] [CrossRef]
- Ruan, X.; Liu, A.; Zhong, M.; Wei, J.; Zhang, W.; Rong, Y.; Liu, W.; Li, M.; Qing, X.; Chen, G. Silencing LGR6 Attenuates Stemness and Chemoresistance via Inhibiting Wnt/β-Catenin Signaling in Ovarian Cancer. Mol. Ther. Oncolytics 2019, 14, 94–106. [Google Scholar] [CrossRef]
- Makena, M.R.; Koneru, B.; Nguyen, T.H.; Kang, M.H.; Reynolds, C.P. Reactive oxygen species-mediated synergism of fenretinide and romidepsin in preclinical models of T-cell lymphoid malignancies. Mol. Cancer Ther. 2017, 16, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Makena, M.R.; Nguyen, T.H.; Koneru, B.; Hindle, A.; Chen, W.H.; Verlekar, D.U.; Kang, M.H.; Reynolds, C.P. Vorinostat and Fenretinide Synergize in Preclinical Models of T-cell Lymphoid Malignancies via Reactive Oxygen Species. Am. Assoc. Cancer Res. 2018. [Google Scholar] [CrossRef]
- Gatla, H.R.; Muniraj, N.; Thevkar, P.; Yavvari, S.; Sukhavasi, S.; Makena, M.R. Regulation of Chemokines and Cytokines by Histone Deacetylases and an Update on Histone Decetylase Inhibitors in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1110. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zou, Q.; Sun, M.; Chen, J.; Wang, T.; Bai, Y.; Chen, Z.; Chen, B.; Zhou, M. Reversion of trichostatin A resistance via inhibition of the Wnt signaling pathway in human pancreatic cancer cells. Oncol. Rep. 2014, 32, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Tai, D.; Wells, K.; Arcaroli, J.; Vanderbilt, C.; Aisner, D.L.; Messersmith, W.A.; Lieu, C.H. Targeting the WNT signaling pathway in cancer therapeutics. The Oncol. 2015, 20, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- O’Cearbhaill, R.E.; McMeekin, D.S.; Mantia-Smaldone, G.; Gunderson, C.; Sabbatini, P.; Cattaruzza, F.; Fischer, M.; Kapoun, A.M.; Xu, L.; Dupont, J. Phase 1b of WNT inhibitor ipafricept (IPA, decoy receptor for WNT ligands) with carboplatin (C) and paclitaxel (P) in recurrent platinum-sensitive ovarian cancer (OC). Am. Soc. Clin. Oncol. 2016, 2515. [Google Scholar] [CrossRef]
- Dreyer, S.B.; Chang, D.K.; Bailey, P.; Biankin, A.V. Pancreatic Cancer Genomes: Implications for Clinical Management and Therapeutic Development. Am. Assoc. Cancer Res. 2017, 1638–1646. [Google Scholar] [CrossRef]
- Dou, H.; Shen, R.; Tao, J.; Huang, L.; Shi, H.; Chen, H.; Wang, Y.; Wang, T. Curcumin suppresses the colon cancer proliferation by inhibiting Wnt/β-catenin pathways via miR-130a. Front. Pharmacol. 2017, 8, 877. [Google Scholar] [CrossRef] [PubMed]
- Al-Hendy, A.; Diamond, M.P.; Boyer, T.G.; Halder, S.K. Vitamin D3 inhibits Wnt/β-catenin and mTOR signaling pathways in human uterine fibroid cells. J. Clin. Endocrinol. Metab. 2016, 101, 1542–1551. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Compound | Target | Current Clinical Status |
|---|---|---|
| LGK974/WNT974 | Porcupine inhibitor | Phase I (NCT01351103)—malignancies dependent on Wnt ligands, including pancreatic adenocarcinoma. Phase II (NCT02649530)—metastatic head and neck squamous cell carcinoma |
| ETC-1922159 | Porcupine inhibitor | Phase I (NCT02521844)—advanced solid tumors |
| OMP-18R5 (Vantictumab) | Frizzled receptor | Phase I—solid tumors (NCT01345201), phase I (NCT01973309)—combination with paclitaxel in locally recurrent or metastatic breast cancer, phase 1 (NCT01957007)—combination with docetaxel in non-small cell lung cancer, phase I (NCT02005315)—combination with nab-paclitaxel and gemcitabine in stage IV pancreatic cancer |
| OMP-54 F28 (Ipafricept) | Fzd8-Fc fusion protein | Phase I (NCT01608867)—solid tumors, phase Ib (NCT02069145)—combination with sorafenib in hepatocellular cancer, phase 1(NCT02092363)—combination with paclitaxel and carboplatin in recurrent platinum-sensitive ovarian cancer, phase I (NCT02050178)—combination with Nab-paclitaxel and gemcitabine stage IV pancreatic cancer |
| OTSA-101 | FZD10 mAb | Phase I (NCT01469975)—synovial sarcoma |
| DKN-01 | DKK, dickkopf-related protein | Phase I/II (NCT03645980)—advanced liver cancer, Phase I (NCT01711671)—DKN-01 and lenalidomide/dexamethasone in relapsed or refractory multiple myeloma, Phase I (NCT01457417)—advanced solid tumors, Phase I (NCT02013154)—combination with paclitaxel or pembrolizumab in relapsed or refractory esophagogastric malignancies, Phase I (NCT02375880)—combination with gemcitabine and cisplatin in patients with carcinoma primary to the intra- or extra-hepatic biliary system or gallbladder, Phase II (NCT03395080)—monotherapy or in combination with paclitaxel in recurrent ovarian cancer |
| PRI-724 | β-catenin/CBP | Phase I/II (NCT01606579)—myeloid leukemia, Phase I (NCT01302405)—advanced solid tumors, Phase I (NCT01764477)—combination with gemcitabine in advanced or metastatic pancreatic adenocarcinoma |
| CWP232291 | β-catenin | Phase I/II (NCT03055286 and NCT01398462)—acute myeloid leukemia, Phase I (NCT02426723)—relapsed or refractory myeloma |
| Foxy-5 | Wnt5a | Phase I (NCT02020291)—anti-tumor activity and PK profiles in metastatic breast, colon, or prostate cancer |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ram Makena, M.; Gatla, H.; Verlekar, D.; Sukhavasi, S.; K. Pandey, M.; C. Pramanik, K. Wnt/β-Catenin Signaling: The Culprit in Pancreatic Carcinogenesis and Therapeutic Resistance. Int. J. Mol. Sci. 2019, 20, 4242. https://doi.org/10.3390/ijms20174242
Ram Makena M, Gatla H, Verlekar D, Sukhavasi S, K. Pandey M, C. Pramanik K. Wnt/β-Catenin Signaling: The Culprit in Pancreatic Carcinogenesis and Therapeutic Resistance. International Journal of Molecular Sciences. 2019; 20(17):4242. https://doi.org/10.3390/ijms20174242
Chicago/Turabian StyleRam Makena, Monish, Himavanth Gatla, Dattesh Verlekar, Sahithi Sukhavasi, Manoj K. Pandey, and Kartick C. Pramanik. 2019. "Wnt/β-Catenin Signaling: The Culprit in Pancreatic Carcinogenesis and Therapeutic Resistance" International Journal of Molecular Sciences 20, no. 17: 4242. https://doi.org/10.3390/ijms20174242
APA StyleRam Makena, M., Gatla, H., Verlekar, D., Sukhavasi, S., K. Pandey, M., & C. Pramanik, K. (2019). Wnt/β-Catenin Signaling: The Culprit in Pancreatic Carcinogenesis and Therapeutic Resistance. International Journal of Molecular Sciences, 20(17), 4242. https://doi.org/10.3390/ijms20174242