Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts

 ,
, 

Abstract
1. Introduction
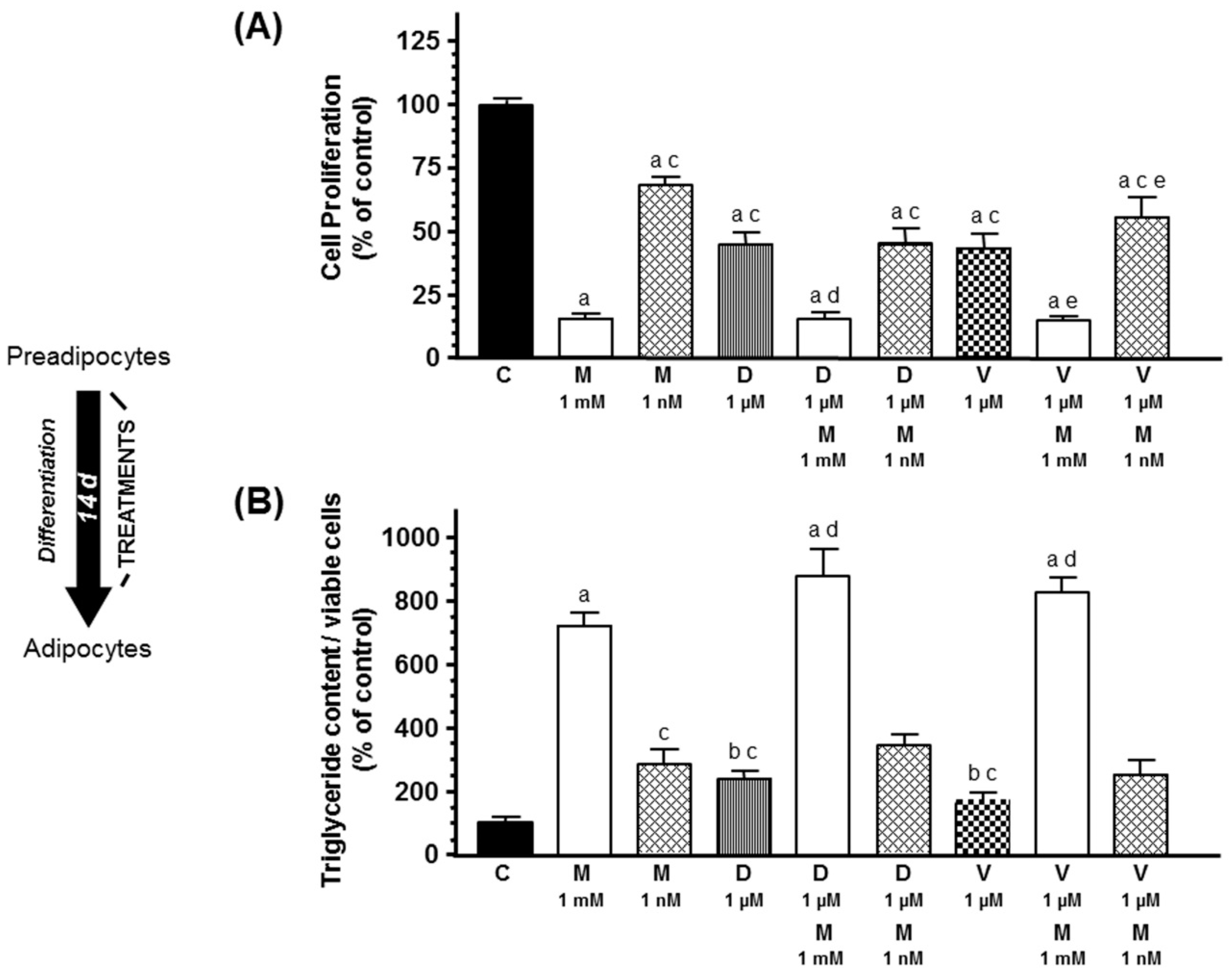
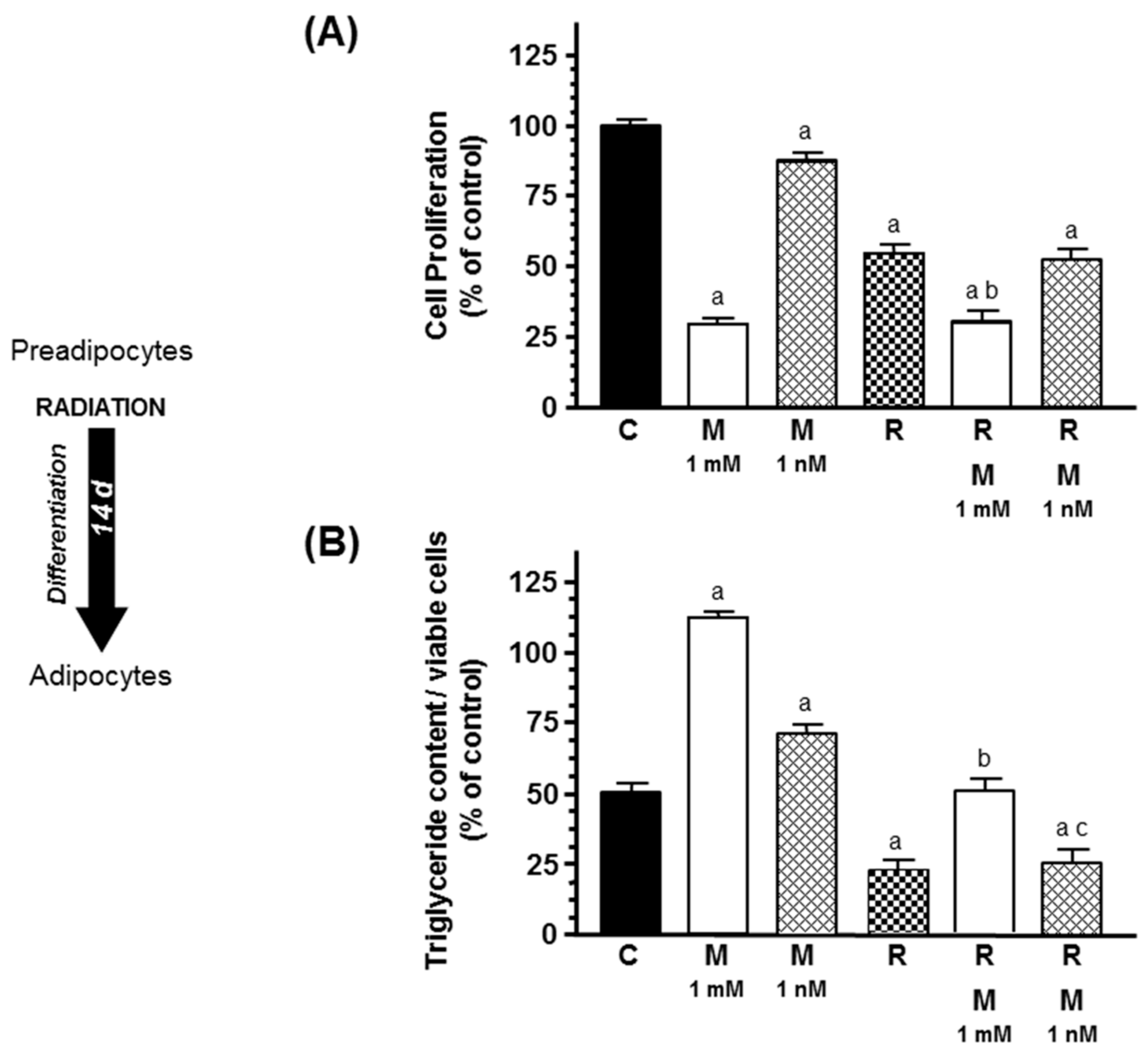
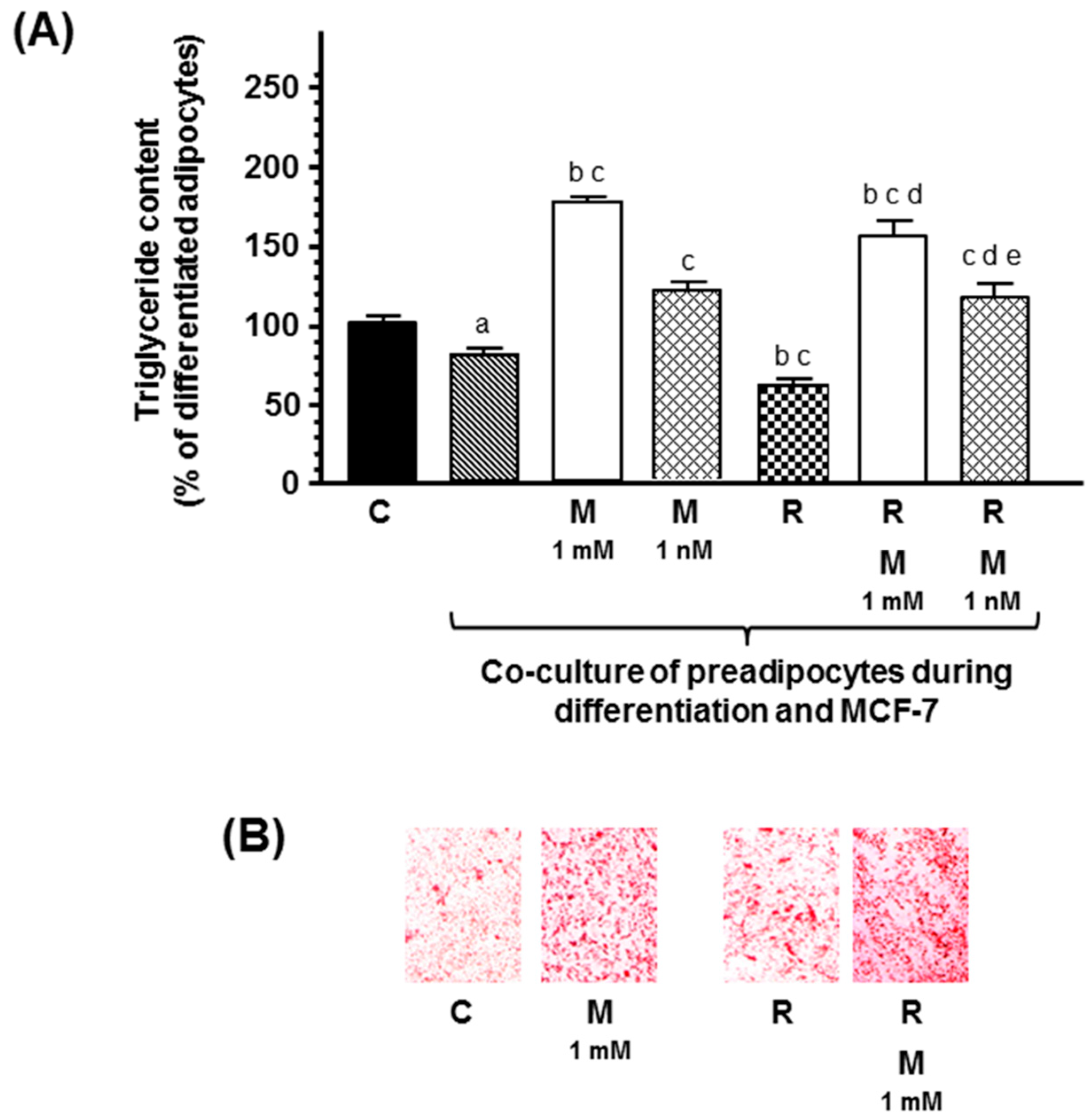
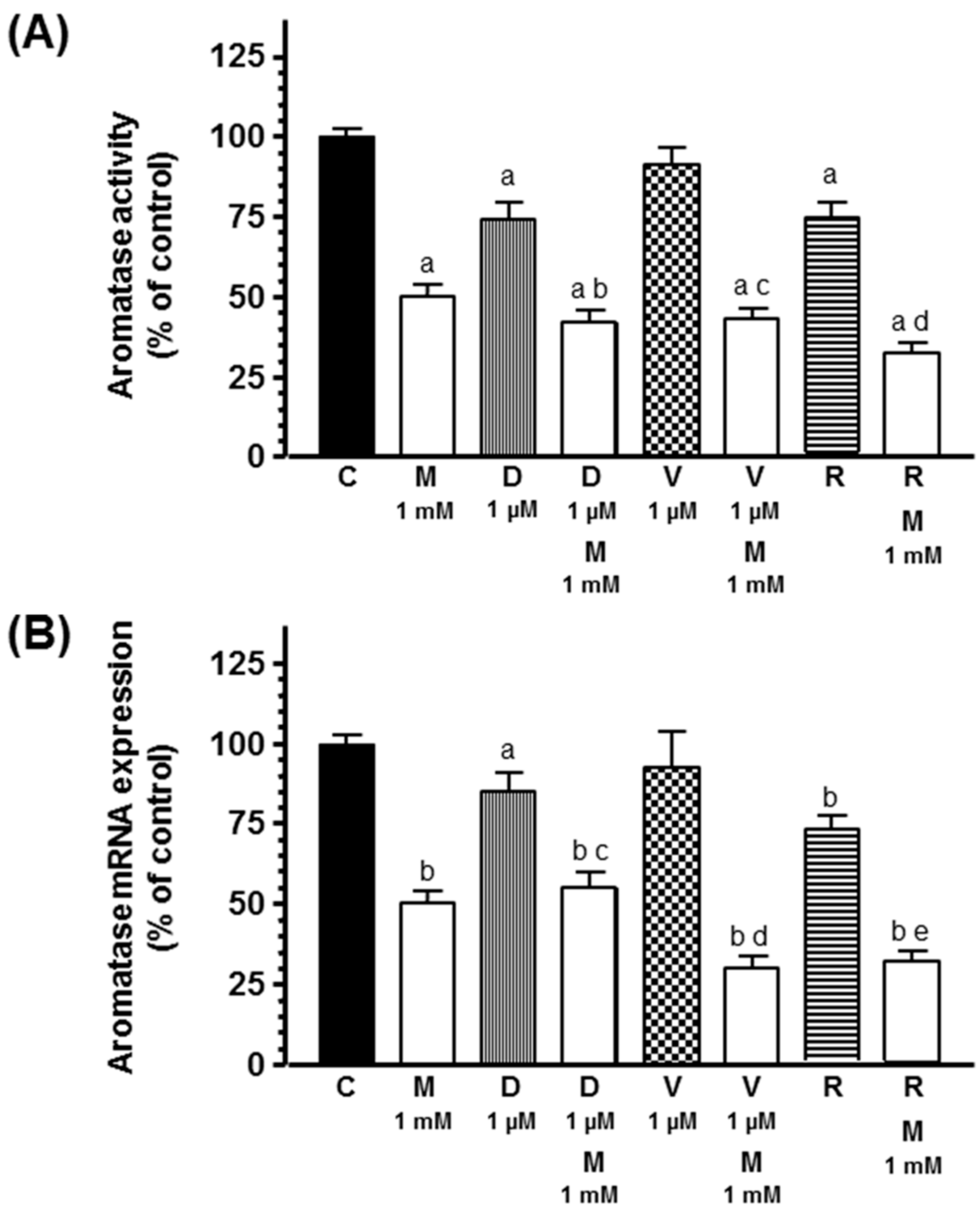
2. Results
2.1. Effects of Melatonin on Docetaxel- and Vinorelbine-Induced Changes on Adipose Differentiation of Human Breast Preadipocytes
2.2. Effects of Melatonin on Radiation-Induced Changes on Adipose Differentiation of Human Breast Preadipocytes
2.3. Effects of Melatonin on Docetaxel-, Vinorelbine-, and Radiation-Induced Changes on C/EBPα and PPARγ Expression in Human Breast Preadipocytes
2.4. Effects of Melatonin on Docetaxel-, Vinorelbine-, and Radiation-Induced Changes on Aromatase Activity and Expression of Human Breast Preadipocytes
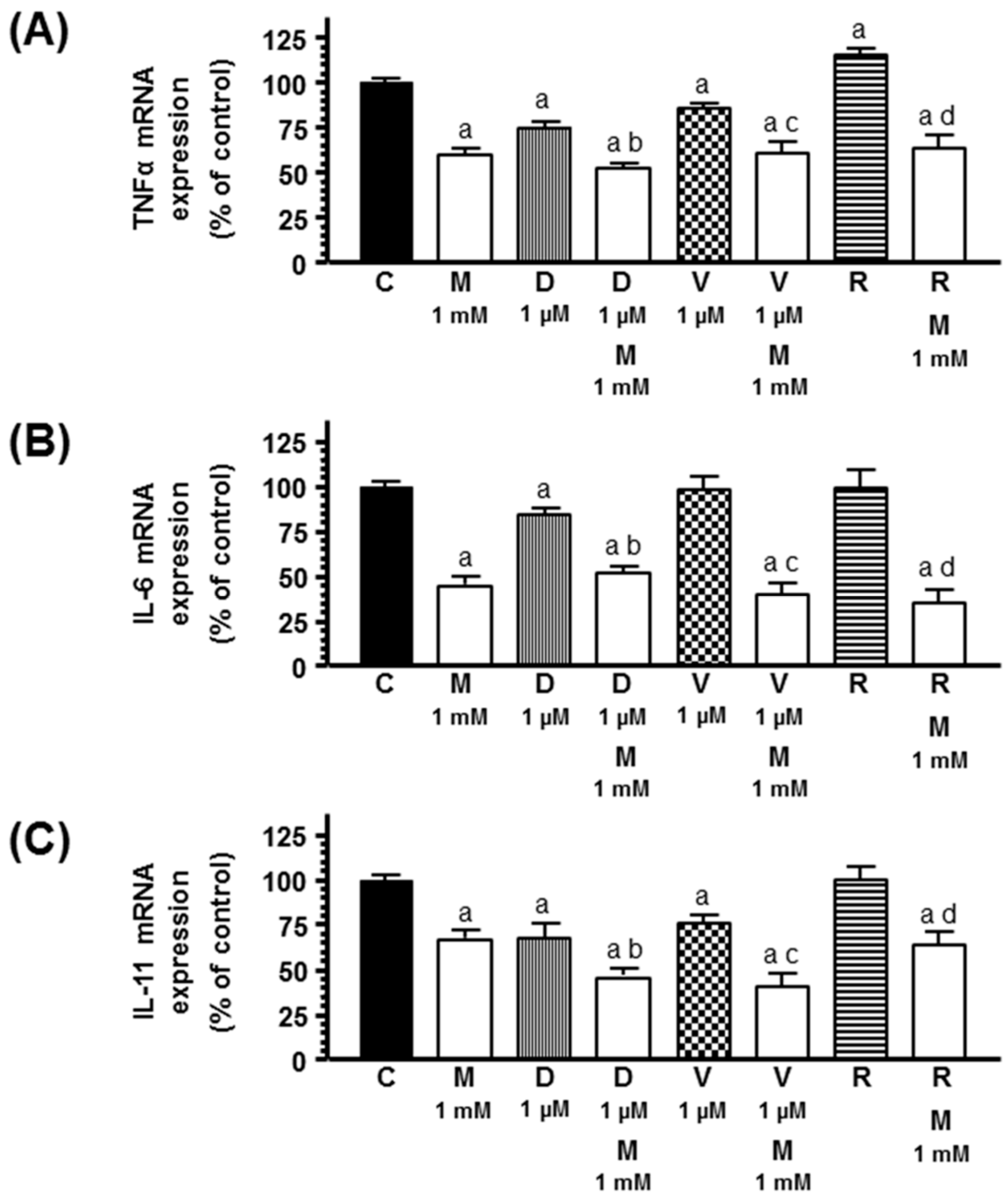
2.5. Effects of Melatonin on Docetaxel-, Vinorelbine-, and Radiation-Induced Changes on Tumor Necrosis Factor α (TNFα), Interleukin 6 (IL-6), and IL-11 mRNA Expression in Human Breast Preadipocytes
2.6. Effects of Melatonin on Docetaxel-, Vinorelbine-, and Radiation-Induced Changes on cyclooxygenase 1 (COX-1) and COX-2 mRNA Expression in Human Breast Preadipocytes
3. Discussion
4. Materials and Methods
4.1. Cells and Culture Conditions
4.2. Co-culture of Human Breast Preadipocites and MCF-7 Cells
4.3. Ionizing Radiation Treatment
4.4. Measurement of Cellular Proliferation
4.5. Quantitation of Triglycerides by Oil Red O Staining
4.6. Measurement of Cellular Aromatase Activity
4.7. Measurement of Specific mRNA Gene Expression
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.Y.; Lehuédé, C.; Laurent, V.; Dirat, B.; Dauvillier, S.; Bochet, L.; Le Gonidec, S.; Escourrou, G.; Valet, P.; Muller, C. Adipose tissue and breast epithelial cells: A dangerous dynamic duo in breast cancer. Cancer Lett. 2012, 324, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Goradel, N.H.; Farhood, B.; Salehi, E.; Solhjoo, S.; Toolee, H.; Kharazinejad, E.; Mortezaee, K. Tumor microenvironment: Interactions and therapy. J. Cell Physiol. 2019, 234, 5700–5721. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Lin, Z.; Imir, G.; Amin, S.; Demura, M.; Yilmaz, B.; Martin, R.; Utsunomiya, H.; Thung, S.; Gurates, B.; et al. Regulation of aromatase expression in estrogen-responsive breast and uterine disease: From bench to treatment. Pharm. Rev. 2005, 57, 359–383. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.I.; Zhou, J.; Sasano, H.; Suzuki, T.; Zeitoun, K.M.; Bulun, S.E. Tumor necrosis factor α and interleukin 11 secreted by malignant breast epithelial cells inhibit adipocyte differentiation by selectively down-regulating CCAAT/enhancer binding protein α and peroxisome proliferator-activated receptor γ: Mechasnism of desmoplastic reaction. Cancer Res. 2001, 61, 2250–2255. [Google Scholar] [PubMed]
- Chen, D.; Reierstad, S.; Lu, M.; Lin, Z.; Ishikawa, H.; Bulun, S.E. Regulation of breast cancer-associated aromatase promoters. Cancer Lett. 2009, 273, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Poglio, S.; Galvani, S.; Bour, S.; André, M.; Prunet-Marcassus, B.; Pénicaud, L.; Casteilla, L.; Cousin, B. Adipose tissue sensitivity to radiation exposure. Am. J. Pathol. 2009, 174, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Barceló, E.J.; Cos, S.; Mediavilla, D.; Martínez-Campa, C.; González, A.; Alonso-González, C. Melatonin-estrogen interactions in breast cancer. J. Pineal Res. 2005, 38, 217–222. [Google Scholar] [CrossRef]
- Cos, S.; González, A.; Martínez-Campa, C.; Mediavilla, M.D.; Alonso-González, C.; Sánchez-Barceló, E.J. Estrogen-signaling pathway: A link between breast cancer and melatonin oncostatic actions. Cancer Detect. Prev. 2006, 30, 118–128. [Google Scholar] [CrossRef]
- Hill, S.M.; Belancio, V.P.; Dauchy, R.T.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.W.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, R183–R204. [Google Scholar] [CrossRef]
- Martínez-Campa, C.; González, A.; Mediavilla, M.D.; Alonso-González, C.; Sánchez-Barceló, E.J.; Cos, S. Melatonin enhances the inhibitory effect of aminoglutethimide on aromatase activity in MCF-7 human breast cancer cells. Breast Cancer Res. Treat. 2005, 94, 249–254. [Google Scholar] [CrossRef]
- Alonso-González, C.; González, A.; Martínez-Campa, C.; Gómez-Arozamena, J.; Cos, S. Melatonin sensitizes human breast cancer cells to ionizing radiation by downregulating proteins involved in double-strand DNA break repair. J. Pineal Res. 2015, 58, 189–197. [Google Scholar] [CrossRef]
- Alonso-González, C.; González, A.; Martínez-Campa, C.; Menéndez-Menéndez, J.; Gómez-Arozamena, J.; García-Vidal, A.; Cos, S. Melatonin enhancement of the radiosensitivity of human breast cancer cells is associated with the modulation of proteins involved in estrogen biosynthesis. Cancer Lett. 2016, 370, 145–152. [Google Scholar] [CrossRef]
- Alonso-González, C.; Menéndez-Menéndez, J.; González-González, A.; González, A.; Cos, S.; Martínez-Campa, C. Melatonin enhances the apoptotic effects and modulates the changes in gene expression induced by docetaxel in MCF-7 human breast cancer cells. Int. J. Oncol. 2018, 52, 560–570. [Google Scholar]
- Najafi, M.; Salehi, E.; Farhood, B.; Nashtaei, M.S.; Goradel, N.H.; Khanlarkhani, N.; Namjoo, Z. Adjuvant chemotherapy with melatonin for targeting human cancers: A review. J. Cell Physiol. 2019, 234, 2356–2372. [Google Scholar] [CrossRef]
- Ramírez-Zacarias, J.L.; Castro-Muñozledo, F.; Kuri-Harcuch, W. Quantitation of adipose conversión and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef]
- Bulun, S.E.; Chen, D.; Lu, M.; Zhao, H.; Cheng, Y.; Demura, M.; Yilmaz, B.; Martin, R.; Utsunomiya, H.; Thung, S.; et al. Aromatase excess in cancers of breast, endometrium and ovary. J. Steroid Biochem. Mol. Biol. 2007, 106, 81–96. [Google Scholar] [CrossRef]
- Guerrero, J.; Tobar, N.; Cáceres, M.; Espinoza, L.; Escobar, P.; Dotor, J.; Smith, P.C.; Martínez, J. Soluble factors derived from tumor mammary cell lines induce a stromal mammary adipose reversion in human and mice adipose cells. Possible role of TGF-β1 and TNF-α. Breast Cancer Res. Treat. 2010, 119, 497–508. [Google Scholar] [CrossRef]
- Cos, S.; Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C. Melatonin modulation of crosstalk among malignant epithelial, endothelial and adipose cells in breast cancer. Oncol. Lett. 2014, 8, 487–492. [Google Scholar] [CrossRef]
- He, J.; Chen, Q.; Gu, H.; Chen, J.; Zhang, E.; Guo, X.; Huang, X.; Yan, H.; He, D.; Yang, Y.; et al. Therapeutic effects of the novel subtype-selective histone deacetylase (HDAC) inhibitor chidamide on myeloma-associated bone disease. Haematologica 2018, 103, 1369–1379. [Google Scholar] [CrossRef]
- Cos, S.; González, A.; Martínez-Campa, C.; Mediavilla, M.D.; Alonso-González, C.; Sánchez-Barceló, E.J. Melatonin as a selective estrogen enzyme modulator. Curr. Cancer Drug Tar. 2008, 8, 691–702. [Google Scholar] [CrossRef]
- Gonzalez, A.; Cos, S.; Martinez-Campa, C.; Alonso-Gonzalez, C.; Sanchez-Mateos, S.; Mediavilla, M.D.; Sanchez-Barcelo, E.J. Selective estrogen enzime modulator (SEEM) actions of melatonin in human breast cancer cells. J. Pineal Res. 2008, 45, 86–92. [Google Scholar] [CrossRef]
- González, A.; Martinez-Campa, C.; Mediavilla, M.D.; Alonso-González, C.; Sánchez-Barceló, E.J.; Cos, S. Inhibitory effects of pharmacological doses of melatonin on aromatase activity and expression in rat glioma cells. Br. J. Cancer 2017, 97, 755–760. [Google Scholar] [CrossRef][Green Version]
- González, A.; Alvarez-García, V.; Martinez-Campa, C.; Alonso-González, C.; Cos, S. Melatonin promotes differentiation of 3T3-L1 fibroblasts. J. Pineal Res. 2012, 52, 12–20. [Google Scholar] [CrossRef]
- Knower, K.C.; To, S.Q.; Takagi, K.; Miki, Y.; Sasano, H.; Simpson, E.R.; Clyne, C.D. Melatonin suppresses aromatase expression and activity in breast cancer associated fibroblasts. Breast Cancer Res. Treat. 2012, 132, 765–771. [Google Scholar] [CrossRef]
- Maldonado, M.D.; Siu, A.W.; Sánchez-Hidalgo, M.; Acuña-Castroviejo, D.; Escames, G. Melatonin and lipid uptake by murine fibroblasts: Clinical implications. Neuro Endocrinol. Lett. 2006, 27, 601–608. [Google Scholar]
- Kato, H.; Tanaka, G.; Masuda, S.; Ogasawara, J.; Sakurai, T.; Kizaki, T.; Ohno, H.; Izawa, T. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3-L1 preadipocytes. J. Pineal Res. 2015, 59, 267–275. [Google Scholar] [CrossRef]
- Yang, W.; Tang, K.; Wang, Y.; Zhang, Y.; Zan, L. Melatonin promotes triacylglycerol accumulation via MT2 receptor during differentiation in bovine intramuscular preadipocytes. Sci. Rep. 2017, 7, 15080. [Google Scholar] [CrossRef]
- Alonso-Vale, M.I.; Peres, S.B.; Vernochet, C.; Farmer, S.R.; Lima, F.B. Adipocyte differentiation is inhibited by melatonin through the regulation of c/EBPβ transcriptional activity. J. Pineal Res. 2009, 47, 221–227. [Google Scholar] [CrossRef]
- Ntambi, J.M.; Kim, Y.C. Adipocyte differentiation and gene expression. J. Nutr. 2000, 130, 3122S–3126S. [Google Scholar] [CrossRef]
- Lane, M.D.; Tang, Q.Q. From multipotent stem cell to adipocyte. Birth Defects Res. A 2005, 73, 476–477. [Google Scholar] [CrossRef]
- Mediavilla, M.D.; Sanchez-Barceló, E.J.; Tan, D.X.; Manchester, L.; Reiter, R.J. Basic mechanisms involved in the anti-cancer effects of melatonin. Curr. Med. Chem. 2010, 17, 4462–4481. [Google Scholar] [CrossRef]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C.; Cos, S. Antiangiogenic effects of melatonin in endothelial cell cultures. Microvascular Res. 2013, 87, 25–33. [Google Scholar] [CrossRef]
- Maestroni, G.J.; Conti, A. Melatonin in human Breast cancer tissue: Association with nuclear grade and estrogen receptor status. Lab. Invest. 1996, 75, 557–561. [Google Scholar]
- Wang, Y.; Zhu, G.; Wang, J.; Chen, J. Irradiation alters the differentiation potential of bone marrow mesenchymal stem cells. Mol. Med. Rep. 2016, 13, 213–223. [Google Scholar] [CrossRef]
- Lee, J.; Lee, J.; Jung, E.; Kim, Y.S.; Roh, K.; Jung, K.H.; Park, D. Ultraviolet A regulates adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells via up-regulation of kruppel-like factor 2. J. Biol. Chem. 2010, 285, 32647–32656. [Google Scholar] [CrossRef]
- Schreder, K.; Rapp, F.; Tsoukala, I.; Rzeznik, V.; Wabitsch, M.; Fischer-Posovszky, P.; Fournier, C. Impact of X-ray exposure on the proliferation and differentiation of human pre-adipocytes. Int. J. Mol. Sci. 2018, 19, 2717. [Google Scholar] [CrossRef]
- Russo, J.; Russo, I.H. The role of estrogen in the initiation of breast cancer. J. Steroid Biochem. Mol. Biol. 2006, 102, 89–96. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Kim, S.J.; Akazawa, K.; Kamigaki, S.; Ueda, S.; Yanagisawa, T.; Inoue, T.; Taguchi, T.; Tamaki, Y.; Noguchi, S. Down-regulation of intratumoral aromatase messenger RNA levels by docetaxel in human breast cancers. Clin. Cancer Res. 2004, 10, 8163–8169. [Google Scholar] [CrossRef]
- Cos, S.; Martínez-Campa, C.; Mediavilla, M.D.; Sánchez-Barceló, E.J. Melatonin modulates aromatase activity in MCF-7 human breast cancer cells. J. Pineal Res. 2005, 38, 136–142. [Google Scholar] [CrossRef]
- Alvarez-García, V.; González, A.; Martinez-Campa, C.; Alonso-González, C.; Cos, S. Melatonin modulates aromatase activity and expression in endothelial cells. Oncol. Rep. 2013, 29, 2058–2064. [Google Scholar] [CrossRef]
- Brueggemeier, R.W.; Richards, J.A.; Petrel, T.A. Aromatase and cyclooxygenases: Enzymes in breast cancer. J. Steroid Biochem. Mol. Biol. 2003, 86, 501–507. [Google Scholar] [CrossRef]
- Liu, X.-H.; Rose, D.P. Differential expression and regulation of cyclooxygenase-1 and -2 in two human breast cancer cell lines. Cancer Res. 1996, 56, 5125–5127. [Google Scholar]
- Erovic, B.M.; Woegerbauer, M.; Pammer, J.; Selzer, E.; Grasl, M.C.; Thurnher, D. Strong evidence for up-regulation of cyclooxigenase-1 in head and neck cancer. Eur. J. Clin. Invest. 2008, 38, 61–66. [Google Scholar] [CrossRef]
- Díaz-Cruz, E.S.; Shapiro, C.L.; Brueggemeier, R.W. Cyclooxygenase inhibitors suppress aromatase expression and activity in breast cancer cells. J. Clin. Endocrinol. Metab. 2005, 90, 2563–2570. [Google Scholar] [CrossRef]
- Howe, L.R. Inflammation and breast cancer. Cyclooxigenase/prostaglandin signaling and breast cancer. Breast Cancer Res. 2007, 9, 210. [Google Scholar] [CrossRef]
- Nakano, T.; Goto, K.; Wakabayashi, I. Docetaxel inhibits cyclooxygenase-2 induction in vascular smooth muscle cells. Eur. J. Pharmacol. 2011, 654, 150–154. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Marmo, T.P.; Dixon, D.A.; Dannenberg, A.J. Regulation of cyclooxygenase-2 mRNA stability by taxanes: Evidence for involvement of p38, MAPKAPK-2 and HuR. J. Biol. Chem. 2003, 278, 37637–37647. [Google Scholar] [CrossRef]
- Martinez Campa, C.; González, A.; Mediavilla, M.D.; Alonso-González, C.; Alvarez-García, V.; Sánchez-Barceló, E.J.; Cos, S. Melatonin inhibits aromatase promoter expression by regulating cyclooxygenases expression and activity in breast cancer cells. Br. J. Cancer 2009, 101, 1613–1619. [Google Scholar] [CrossRef]
- Torti, F.M.; Torti, S.V.; Larrick, J.W.; Ringold, G.M. Modulation of adipocyte differentiation by tumor necrosis factor and transforming growth factor β. J. Cell Biol. 1989, 108, 1105–1113. [Google Scholar] [CrossRef]
- Keller, D.C.; Du, X.X.; Srour, E.F.; Hoffman, R.; Williams, D.A. Interleukin-11 inhibits adipogenesis and stimulates myelopoiesis in human long-term marrow cultures. Blood 1993, 82, 1428–1435. [Google Scholar]
- Rubin, G.L.; Zhao, Y.; Kalus, A.M.; Simpson, E.R. Peroxisome proliferator-activated receptor γ ligands inhibit estrogen biosynthesis in human breast adipose tissue: Possible implications for breast cancer therapy. Cancer Res. 2000, 60, 1604–1608. [Google Scholar]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martinez-Campa, C.; Cos, S. Melatonin interferes in the desmoplastic reaction in breast cancer by regulating cytokine production. J. Pineal Res. 2012, 52, 282–290. [Google Scholar] [CrossRef]
- Tsavaris, N.; Kosmas, C.; Vadiaka, M.; Kanelopoulos, P.; Boulamatsis, D. Immune changes in patients with advanced breast cancer undergoing chemotherapy with taxanes. Br. J. Cancer 2002, 87, 21–27. [Google Scholar] [CrossRef]
- Meng, G.; Tang, X.; Yang, Z.; Benesch, M.G.K.; Marshall, A.; Murray, D.; Hemmings, D.G.; Wuest, F.; McMullen, T.P.W.; Brindley, D.N. Implications for breast cancer treatment from increased autotaxin production in adipose tissue after radiotherapy. FASEB J. 2017, 31, 4064–4077. [Google Scholar] [CrossRef]
- Farhood, B.; Goradel, N.H.; Mortezaee, K.; Khanlarkhani, N.; Salehi, E.; Nashtaei, M.S.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Shabeeb, D.; Musa, A.E.; et al. Melatonin as an adjuvant in radiotherapy for radioprotection and radiosensitization. Clin. Transl. Oncol. 2019, 21, 268–279. [Google Scholar] [CrossRef]
- Martínez-Campa, C.; Menéndez-Menéndez, J.; Alonso-González, C.; González, A.; Álvarez-García, V.; Cos, S. What is known about melatonin, chemotherapy and altered gene expression in breast cancer. Oncol. Lett. 2017, 13, 2003–2014. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ackerman, G.E.; Smith, M.E.; Mendelson, C.R.; MacDonald, P.C.; Simpson, E.R. Aromatization of androstenedione by human adipose tissue stromal cells in monolayer culture. J. Clin. Endocrinol. Metab. 1981, 53, 412–417. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔC(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Sequence | Concentration (nM) |
|---|---|---|
| hARO fw | 5′- GTCGTGGACTTGGTCATGC -3′ | 100 |
| hARO rv | 5′- CGAGTCTGTGCATCCTTCC -3′ | 100 |
| hC/EBPα fw | 5′- AGGTGCTGGAGCTGACCAGT -3′ | 200 |
| hC/EBPα rv | 5′- AAGCCTCGAGATCCGGCGAC -3′ | 200 |
| COX-1 fw | 5′- ACCCGCACGGGCTATTCCGGC -3′ | 200 |
| COX-1 rv | 5′- AGGCGCATGAGCATCTCTCGG -3′ | 200 |
| COX-2 fw | 5′- ATGTATGAGTGTGGGATTTGA -3′ | 200 |
| COX-2 rv | 5′- TCCAAAATCCCTTGAAGTGGG- 3′ | 200 |
| hIL-6 fw | 5′- AGGAGACTTGCCTGGTGAAA -3′ | 200 |
| hIL-6 rv | 5′- CAGGGGTGGTTATTGCATCT -3′ | 200 |
| hIL-11 fw | 5′- GCTGGTTTCGAACTCCTGAC -3′ | 200 |
| hIL-11 rv | 5′- CAGGGTGACTTGTGGAACCT -3′ | 200 |
| hpII fw | 5′- CTCTGAAGCAACAGGAGCTATAGA -3′ | 100 |
| hpII rv | 5′- CAGGCACGATGCTGGTGATG -3′ | 100 |
| hpI.3 fw | 5′- GGGCTTCCTTGTTTTGACTGTAA -3′ | 200 |
| hpI.3 rv | 5′- AGAGGGGGCAATTTAGAGTCTGTT -3′ | 200 |
| hPPARγ fw | 5′- GATGCACTGCCTATGAGCACTT -3′ | 400 |
| hPPARγ rv | 5′- AGAGGTCCACAGAGCTGATTCC -3′ | 400 |
| hS14 fw | 5′- TCCTGCGAGTGCTGTCAGAG -3′ | 100 |
| hS14 rv | 5′- TCACCGCCCTACACATCAAAC -3′ | 100 |
| hTNFα fw | 5′- TGGGGTTTGTGAAACTGTGA -3′ | 50 |
| hTNFα rv | 5′- GTTCCTGCACATTCCCTCTC -3′ | 50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-González, A.; García Nieto, E.; González, A.; Sánchez-Fernández, C.; Alonso-González, C.; Menéndez-Menéndez, J.; Gómez-Arozamena, J.; Cos, S.; Martínez-Campa, C. Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts. Int. J. Mol. Sci. 2019, 20, 3935. https://doi.org/10.3390/ijms20163935
González-González A, García Nieto E, González A, Sánchez-Fernández C, Alonso-González C, Menéndez-Menéndez J, Gómez-Arozamena J, Cos S, Martínez-Campa C. Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts. International Journal of Molecular Sciences. 2019; 20(16):3935. https://doi.org/10.3390/ijms20163935
Chicago/Turabian StyleGonzález-González, Alicia, Enrique García Nieto, Alicia González, Cristina Sánchez-Fernández, Carolina Alonso-González, Javier Menéndez-Menéndez, José Gómez-Arozamena, Samuel Cos, and Carlos Martínez-Campa. 2019. "Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts" International Journal of Molecular Sciences 20, no. 16: 3935. https://doi.org/10.3390/ijms20163935
APA StyleGonzález-González, A., García Nieto, E., González, A., Sánchez-Fernández, C., Alonso-González, C., Menéndez-Menéndez, J., Gómez-Arozamena, J., Cos, S., & Martínez-Campa, C. (2019). Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts. International Journal of Molecular Sciences, 20(16), 3935. https://doi.org/10.3390/ijms20163935