The Activation Status of the TGF-β Transducer Smad2 Is Associated with a Reduced Survival in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Search Results
2.2. Study and Patient Characteristics
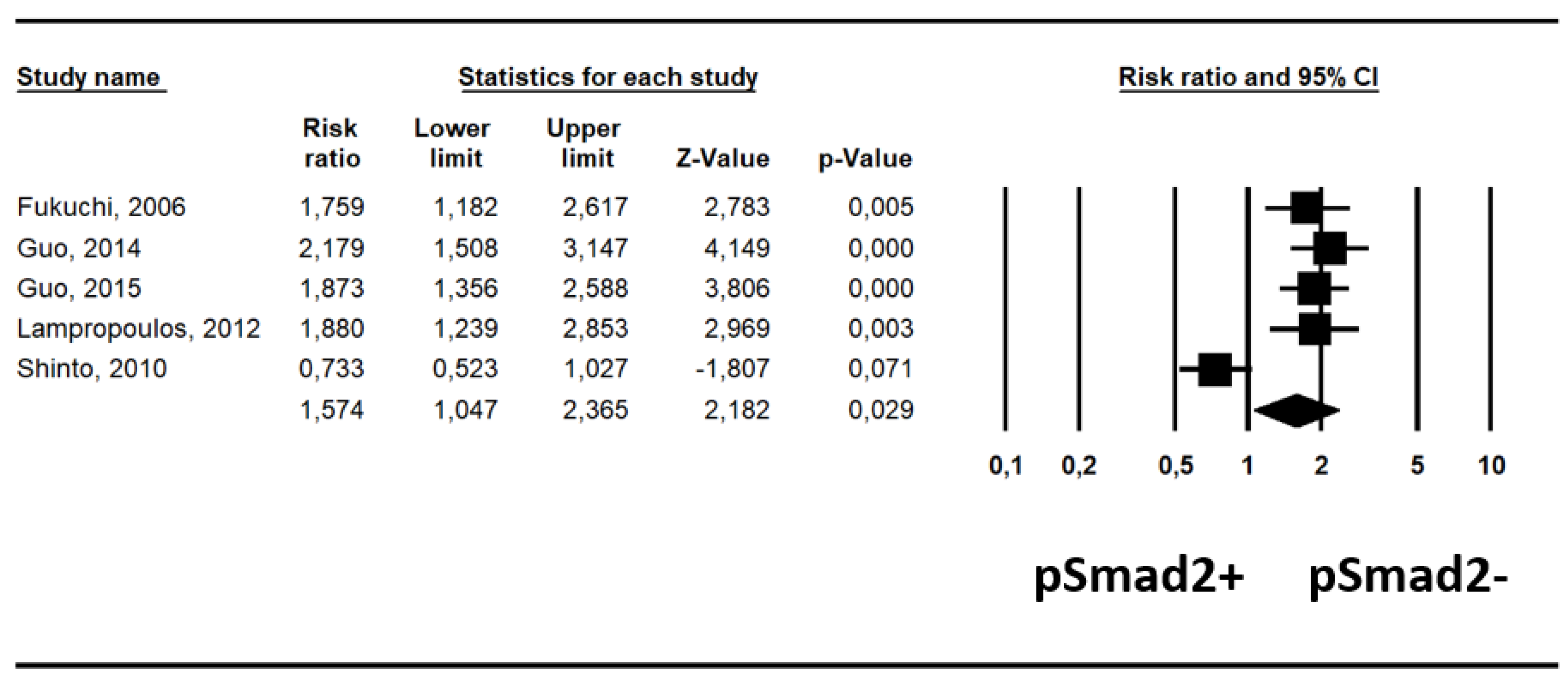
2.3. Association between pSmad2- and pSmad2+ and Survival
2.4. Publication Bias and Meta-Regression Analyses
3. Discussion
4. Materials and Methods
4.1. Inclusion and Exclusion Criteria
4.2. Data Sources and Literature Search Strategy
4.3. Study Selection
4.4. Data Extraction
4.5. Outcomes
4.6. Assessment of Study Quality
4.7. Data Synthesis and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TGF-β | Transforming growth factor beta |
| IHC | immunohistochemistry |
| TNM | Tumor Nodes Metastasis staging system |
| G | Grading |
| MOOSE | Meta-analyses Of Observational Studies in Epidemiology guidelines |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analyses |
References
- Ooshima, A.; Park, J.; Kim, S.J. Phosphorylation status at Smad3 linker region modulates transforming growth factor-β-induced epithelial-mesenchymal transition and cancer progression. Cancer Sci. 2019, 110, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Cytokine & Growth Factor Reviews Smad phospho-isoforms direct context-dependent TGF-b signaling. Cytokine Growth Factor Rev. 2013, 24, 385–399. [Google Scholar] [PubMed]
- Wrana, J.L. Regulation of Smad activity. Cell 2000, 100, 189–192. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.; Feng, X.H. Smads: Transcriptional activators of TGF-beta responses. Cell 1998, 95, 737–740. [Google Scholar] [CrossRef]
- Matsuzaki, K. Smad phosphoisoform signaling specificity: The right place at the right time. Carcinogenesis 2011, 32, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Matsuzaki, K.; Murata, M.; Yamaguchi, T.; Suwa, K.; Okazaki, K. Clinico-Pathological Importance of TGF-β/Phospho-Smad Signaling during Human Hepatic Fibrocarcinogenesis. Cancers 2018, 10, 183. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Gifford, C.C.; Samarakoon, R.; Higgins, P.J. Deregulation of negative controls on TGF-β1 signaling in tumor progression. Cancers 2018, 10, 159. [Google Scholar] [CrossRef] [PubMed]
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFβ in the tumour microenvironment. Nat. Rev. Cancer 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Wu, C.-J.; Chu, G.C.; Xiao, Y.; Ho, D.; Zhang, J.; Perry, S.R.; Labrot, E.S.; Wu, X.; Lis, R.; et al. SMAD4-dependent barrier constrains prostate cancer growth and metastatic progression. Nature 2011, 470, 269–273. [Google Scholar] [CrossRef]
- Katz, L.H.; Likhter, M.; Jogunoori, W.; Belkin, M.; Ohshiro, K.; Mishra, L. TGF-β signaling in liver and gastrointestinal cancers. Cancer Lett. 2016, 379, 166–172. [Google Scholar] [CrossRef]
- Achyut, B.R.; Yang, L. Transforming growth factor-β in the gastrointestinal and hepatic tumor microenvironment. Gastroenterology 2011, 141, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Fleming, N.I.; Jorissen, R.N.; Mouradov, D.; Christie, M.; Sakthianandeswaren, A.; Palmieri, M.; Day, F.; Li, S.; Tsui, C.; Lipton, L.; et al. SMAD2, SMAD3 and SMAD4 Mutations in Colorectal Cancer. Cancer Res. 2013, 73, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; von Deimling, A.; Brandes, A.; Carpentier, A.; Kesari, S.; Sepulveda-Sanchez, J.; Wheeler, H.; Chinot, O.; Cher, L.; Steinbach, J.; et al. Biomarker and Histopathology Evaluation of Patients with Recurrent Glioblastoma Treated with Galunisertib, Lomustine, or the Combination of Galunisertib and Lomustine. Int. J. Mol. Sci. 2017, 18, 995. [Google Scholar] [CrossRef] [PubMed]
- Bruna, A.; Darken, R.S.; Rojo, F.; Ocaña, A.; Peñuelas, S.; Arias, A.; Paris, R.; Tortosa, A.; Mora, J.; Baselga, J.; et al. High TGFβ-Smad Activity Confers Poor Prognosis in Glioma Patients and Promotes Cell Proliferation Depending on the Methylation of the PDGF-B Gene. Cancer Cell 2007, 11, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Gutgold, N.; Davidson, B.; Catane, L.J.; Holth, A.; Hellesylt, E.; Tropé, C.G.; Dørum, A.; Reich, R. TGFβ splicing and canonical pathway activation in high-grade serous carcinoma. Virchows Arch. 2017, 470, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xing, P.; Chen, Y.; Zou, L.; Zhang, Y.; Li, F.; Lu, X. High p-Smad2 expression in stromal fibroblasts predicts poor survival in patients with clinical stage I to IIIA non-small cell lung cancer. World J. Surg. Oncol. 2014, 12, 328. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moustakas, A.; Pardali, K.; Gaal, A.; Heldin, C.H. Mechanisms of TGF-beta signaling in regulation of cell growth and differentiation. Immunol. Lett. 2002, 82, 85–91. [Google Scholar] [CrossRef]
- Fukuchi, M.; Nakajima, M.; Miyazaki, T.; Masuda, N.; Osawa, H.; Manda, R.; Tsukada, K.; Kato, H.; Kuwano, H. Lack of activated Smad2 in transforming growth factor-beta signaling is an unfavorable prognostic factor in patients with esophageal squamous cell carcinoma. J. Surg. Oncol. 2006, 94, 51–56. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, M.; Shen, S.; Guo, Y.; Kuang, G.; Yang, Z.; Dong, Z. Aberrant methylation and decreased expression of the TGF-β/Smad target gene FBXO32 in esophageal squamous cell carcinoma. Cancer 2014, 120, 2412–2423. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, M.; Guo, Y.; Shen, S.; Guo, X.; Dong, Z. FBXO32, a new TGF-β/Smad signaling pathway target gene, is epigenetically inactivated in gastric cardia adenocarcinoma. Neoplasma 2015, 62, 646–657. [Google Scholar] [CrossRef][Green Version]
- Lampropoulos, P.; Zizi-Sermpetzoglou, A.; Rizos, S.; Kostakis, A.; Nikiteas, N.; Papavassiliou, A.G. Prognostic significance of transforming growth factor beta (TGF-β) signaling axis molecules and E-cadherin in colorectal cancer. Tumour Biol. 2012, 33, 1005–1014. [Google Scholar] [CrossRef]
- Shinto, O.; Yashiro, M.; Toyokawa, T.; Nishii, T.; Kaizaki, R.; Matsuzaki, T.; Noda, S.; Kubo, N.; Tanaka, H.; Doi, Y.; et al. Phosphorylated Smad2 in Advanced Stage Gastric Carcinoma. BMC Cancer 2010, 10, 652. [Google Scholar] [CrossRef]
- Voorneveld, P.W.; Jacobs, R.J.; De Miranda, N.F.C.C.; Morreau, H.; Van Noesel, C.J.M.; Offerhaus, G.J.A.; Kodach, L.L.; Hardwick, J.C.H. Evaluation of the prognostic value of pSMAD immunohistochemistry in colorectal cancer. Eur. J. Cancer Prev. 2013, 22, 420–424. [Google Scholar] [CrossRef]
- Muro-Cacho, C.A.; Rosario-Ortiz, K.; Livingston, S.; Muñoz-Antonia, T. Defective transforming growth factor beta signaling pathway in head and neck squamous cell carcinoma as evidenced by the lack of expression of activated Smad2. Clin. Cancer Res. 2001, 7, 1618–1626. [Google Scholar]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef]
- Prunier, C.; Ferrand, N.; Frottier, B.; Pessah, M.; Atfi, A. Mechanism for mutational inactivation of the tumor suppressor Smad2. Mol. Cell. Biol. 2001, 21, 3302–3313. [Google Scholar] [CrossRef]
- Osawa, H.; Shitara, Y.; Shoji, H.; Mogi, A.; Kuwano, H.; Hagiwara, K.; Takenoshita, S. Mutation analysis of transforming growth factor beta type II receptor, Smad2, Smad3 and Smad4 in esophageal squamous cell carcinoma. Int. J. Oncol. 2000, 17, 723–728. [Google Scholar] [CrossRef]
- Luchini, C.; Capelli, P.; Scarpa, A. Pancreatic Ductal Adenocarcinoma and Its Variants. Surg. Pathol. Clin. 2016, 9, 547–560. [Google Scholar] [CrossRef]
- Liao, X.; Hao, Y.; Zhang, X.; Ward, S.; Houldsworth, J.; Polydorides, A.D.; Harpaz, N. Clinicopathological characterization of SMAD4-mutated intestinal adenocarcinomas: A case-control study. PLoS ONE 2019, 14, e0212142. [Google Scholar] [CrossRef]
- Riva, G.; Pea, A.; Pilati, C.; Fiadone, G.; Lawlor, R.T.; Scarpa, A.; Luchini, C. Histo-molecular oncogenesis of pancreatic cancer: From precancerous lesions to invasive ductal adenocarcinoma. World J. Gastrointest. Oncol. 2018, 10, 317–327. [Google Scholar] [CrossRef]
- Luchini, C.; Pea, A.; Lionheart, G.; Mafficini, A.; Nottegar, A.; Veronese, N.; Chianchiano, P.; Brosens, L.A.; Noë, M.; Offerhaus, G.J.A.; et al. Pancreatic undifferentiated carcinoma with osteoclast-like giant cells is genetically similar to, but clinically distinct from, conventional ductal adenocarcinoma. J. Pathol. 2017, 243, 148–154. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Luchini, C.; Parcesepe, P.; Mafficini, A.; Nottegar, A.; Parolini, C.; Veronese, N.; Remo, A.; Manfrin, E. Specific expression patterns of epithelial to mesenchymal transition factors in gestational molar disease. Placenta 2015, 36, 1318–1324. [Google Scholar] [CrossRef]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-mesenchymal transition in cancer: Complexity and opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef]
- Lawlor, R.T.; Veronese, N.; Nottegar, A.; Malleo, G.; Smith, L.; Demurtas, J.; Cheng, L.; Wood, L.D.; Silvestris, N.; Salvia, R.; et al. Prognostic Role of High-Grade Tumor Budding in Pancreatic Ductal Adenocarcinoma: A Systematic Review and Meta-Analysis with a Focus on Epithelial to Mesenchymal Transition. Cancers 2019, 11, 113. [Google Scholar] [CrossRef]
- Du, F.; Liu, H.; Lu, Y.; Zhao, X.; Fan, D. Epithelial-to-Mesenchymal Transition: Liaison between Cancer Metastasis and Drug Resistance. Crit. Rev. Oncog. 2017, 22, 275–282. [Google Scholar] [CrossRef]
- Cappellesso, R.; Luchini, C.; Veronese, N.; Lo Mele, M.; Rosa-Rizzotto, E.; Guido, E.; De Lazzari, F.; Pilati, P.; Farinati, F.; Realdon, S.; et al. Tumor budding as a risk factor for nodal metastasis in pT1 colorectal cancers: A meta-analysis. Hum. Pathol. 2017, 65, 62–70. [Google Scholar] [CrossRef]
- Yu, C.; Liu, Y.; Huang, D.; Dai, Y.; Cai, G.; Sun, J.; Xu, T.; Tian, Y.; Zhang, X. TGF-β1 mediates epithelial to mesenchymal transition via the TGF-β/Smad pathway in squamous cell carcinoma of the head and neck. Oncol. Rep. 2011, 25, 1581–1587. [Google Scholar]
- Kim, A.N.; Jeon, W.-K.; Lim, K.-H.; Lee, H.-Y.; Kim, W.J.; Kim, B.-C. Fyn mediates transforming growth factor-beta1-induced down-regulation of E-cadherin in human A549 lung cancer cells. Biochem. Biophys. Res. Commun. 2011, 407, 181–184. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Steed, H.; Davidge, S.; Fu, Y. TGFβ and EGF synergistically induce a more invasive phenotype of epithelial ovarian cancer cells. Biochem. Biophys. Res. Commun. 2010, 401, 376–381. [Google Scholar] [CrossRef]
- Wendt, M.K.; Allington, T.M.; Schiemann, W.P. Mechanisms of the epithelial-mesenchymal transition by TGF-beta. Future Oncol. 2009, 5, 1145–1168. [Google Scholar] [CrossRef]
- Wieser, R. The transforming growth factor-beta signaling pathway in tumorigenesis. Curr. Opin. Oncol. 2001, 13, 70–77. [Google Scholar] [CrossRef]
- Pardali, K.; Moustakas, A. Actions of TGF-beta as tumor suppressor and pro-metastatic factor in human cancer. Biochim. Biophys. Acta 2007, 1775, 21–62. [Google Scholar]
- Pidugu, V.K.; Wu, M.M.; Yen, A.H.; Pidugu, H.B.; Chang, K.W.; Liu, C.J.; Lee, T.C. IFIT1 and IFIT3 promote oral squamous cell carcinoma metastasis and contribute to the anti-tumor effect of gefitinib via enhancing p-EGFR recycling. Oncogene 2019, 38, 3232–3247. [Google Scholar] [CrossRef]
- Luchini, C.; Veronese, N.; Pea, A.; Sergi, G.; Manzato, E.; Nottegar, A.; Solmi, M.; Capelli, P.; Scarpa, A. Extranodal extension in N1-adenocarcinoma of the pancreas and papilla of Vater: A systematic review and meta-analysis of its prognostic significance. Eur. J. Gastroenterol. Hepatol. 2016, 28, 205–209. [Google Scholar] [CrossRef]
- Luchini, C.; Nottegar, A.; Pea, A.; Solmi, M.; Stubbs, B.; Capelli, P.; Sergi, G.; Manzato, E.; Fassan, M.; Wood, L.D.; et al. Extranodal extension is an important prognostic parameter for both colonic and rectal cancer. Ann. Oncol. 2016, 27, 955–956. [Google Scholar] [CrossRef]
- Veronese, N.; Fassan, M.; Wood, L.D.; Stubbs, B.; Solmi, M.; Capelli, P.; Pea, A.; Nottegar, A.; Sergi, G.; Manzato, E.; et al. Extranodal extension of nodal metastases is a poor prognostic indicator in gastric cancer: A systematic review and meta-analysis. J. Gastrointest. Surg. 2016, 20, 1692–1698. [Google Scholar] [CrossRef]
- Veronese, N.; Luchini, C.; Nottegar, A.; Kaneko, T.; Sergi, G.; Manzato, E.; Solmi, M.; Scarpa, A. Prognostic impact of extra-nodal extension in thyroid cancer: A meta-analysis. J. Surg. Oncol. 2015, 112, 828–833. [Google Scholar] [CrossRef]
- Luchini, C.; Wood, L.D.; Cheng, L.; Nottegar, A.; Stubbs, B.; Solmi, M.; Capelli, P.; Pea, A.; Sergi, G.; Manzato, E.; et al. Extranodal extension of lymph node metastasis is a marker of poor prognosis in oesophageal cancer: A systematic review with meta-analysis. J. Clin. Pathol. 2016. [Google Scholar] [CrossRef]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Stroup, D.F.; Berlin, J.A.; Morton, S.C.; Olkin, I.; Williamson, G.D.; Rennie, D.; Moher, D.; Becker, B.J.; Sipe, T.A.; Thacker, S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. Meta-analysis Of Observational Studies in Epidemiology (MOOSE) group. JAMA 2000, 283, 2008–2012. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonradomised Studies in Meta-Analyses. Available online: http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 6 June 2019).
- Luchini, C.; Stubbs, B.; Solmi, M.; Veronese, N. Assessing the quality of studies in meta-analyses: Advantages and limitations of the Newcastle Ottawa Scale. World J. Meta-Anal. 2017, 5, 80. [Google Scholar] [CrossRef]
- DerSimonian, R.; Laird, N. Meta-analysis in clinical trials revisited. Contemp. Clin. Trials 2015, 45, 139–145. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef]
- Egger, M.; Davey Smith, G.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | N Studies | Risk Ratio (95% CI) | p Value | Heterogeneity (I2%); tau2 | Egger Test ± SE (p Value) |
|---|---|---|---|---|---|
| Unadjusted | 5 | 1.58 (1.05–2.37) | 0.029 | 84%, p < 0.0001 | 8.62 ± 12.51 (p = 0.54) |
| Adjusted | 5 | 1.65 (1.24–2.18) | <0.001 | 4%; p = 0.38 | −2.28 ± 2.83 (p = 0.48) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girolami, I.; Veronese, N.; Smith, L.; Caruso, M.G.; Reddavide, R.; Leandro, G.; Demurtas, J.; Nottegar, A. The Activation Status of the TGF-β Transducer Smad2 Is Associated with a Reduced Survival in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2019, 20, 3831. https://doi.org/10.3390/ijms20153831
Girolami I, Veronese N, Smith L, Caruso MG, Reddavide R, Leandro G, Demurtas J, Nottegar A. The Activation Status of the TGF-β Transducer Smad2 Is Associated with a Reduced Survival in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences. 2019; 20(15):3831. https://doi.org/10.3390/ijms20153831
Chicago/Turabian StyleGirolami, Ilaria, Nicola Veronese, Lee Smith, Maria G. Caruso, Rosa Reddavide, Gioacchino Leandro, Jacopo Demurtas, and Alessia Nottegar. 2019. "The Activation Status of the TGF-β Transducer Smad2 Is Associated with a Reduced Survival in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis" International Journal of Molecular Sciences 20, no. 15: 3831. https://doi.org/10.3390/ijms20153831
APA StyleGirolami, I., Veronese, N., Smith, L., Caruso, M. G., Reddavide, R., Leandro, G., Demurtas, J., & Nottegar, A. (2019). The Activation Status of the TGF-β Transducer Smad2 Is Associated with a Reduced Survival in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences, 20(15), 3831. https://doi.org/10.3390/ijms20153831