Gadolinium Chloride Restores the Function of the Gap Junctional Intercellular Communication between Hepatocytes in a Liver Injury

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
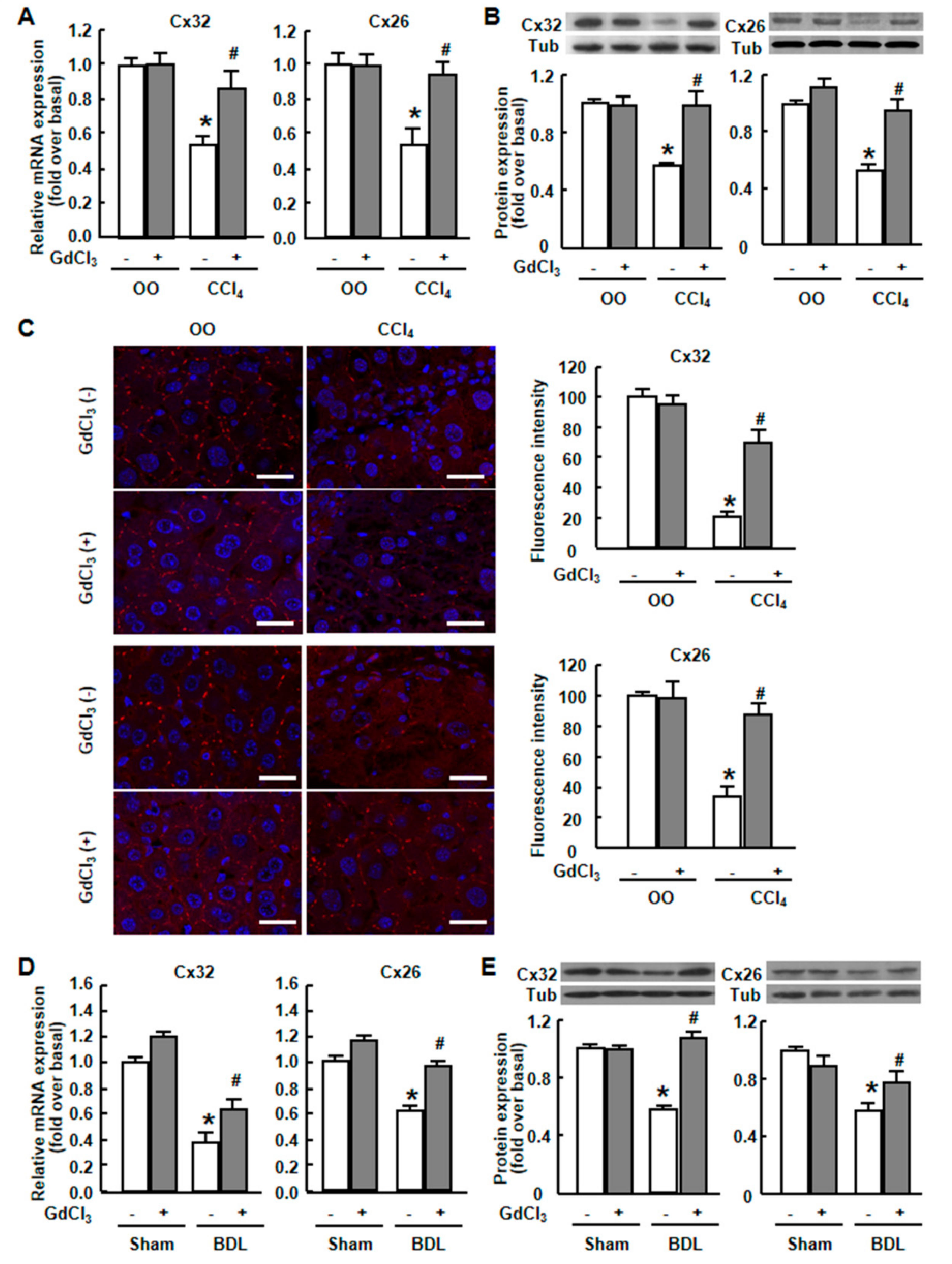
2.1. Cx32 and Cx26 Were Markedly Down-Regulated in Injured Liver
2.2. GdCl3 Prevented the Down-Regulation of Connexins and Restored the Cx32/Cx26 Plaques in Injured Mouse Liver
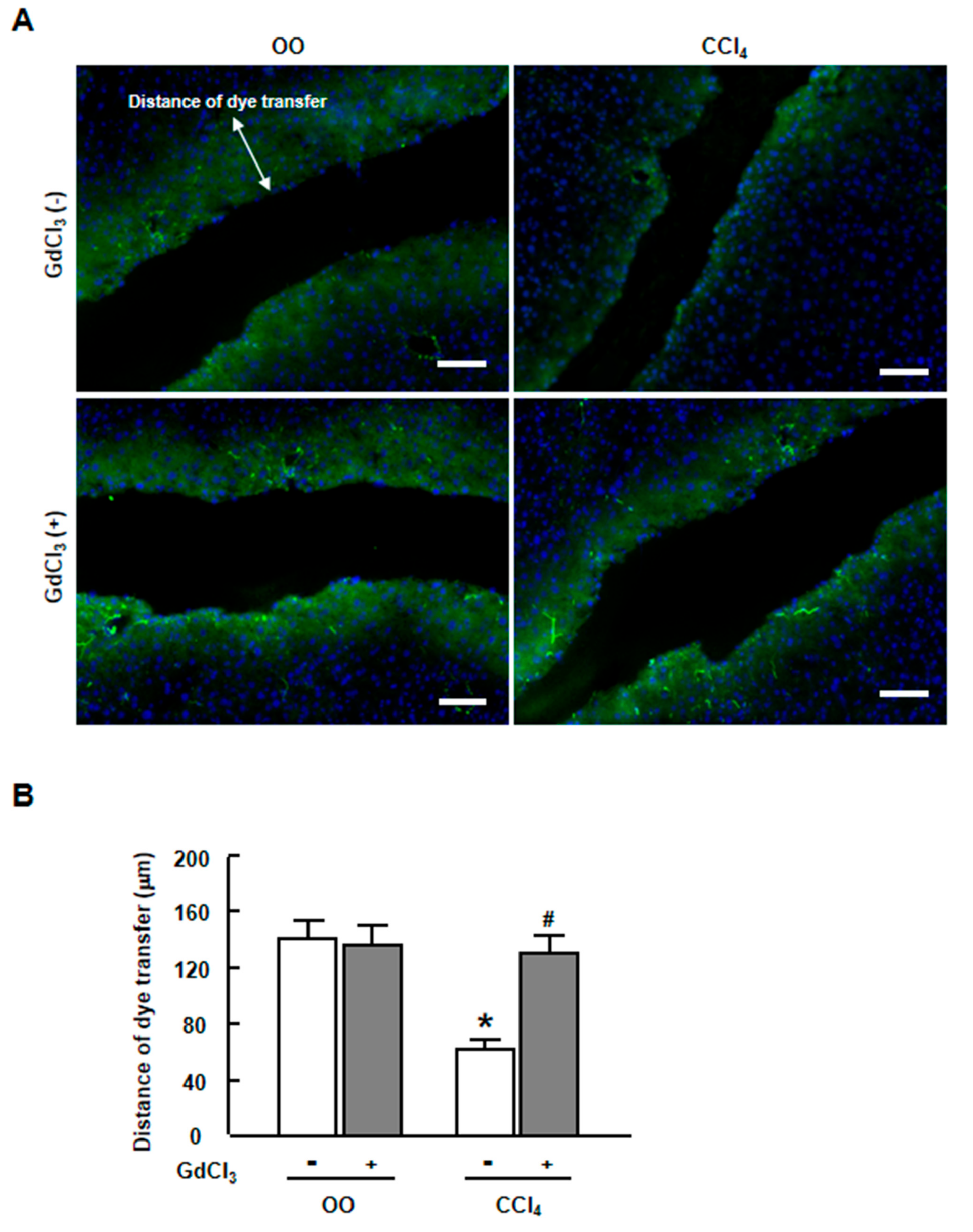
2.3. GdCl3 Restored the Functional GJIC in Injured Liver of CCl4-Treated Mice
2.4. GdCl3 Prevented the Down-Regulation of Cx32 and Cx26 Induced by CCl4 or LPS in Primary Hepatocytes
2.5. GdCl3 Protected Hepatocytes and Contributed to the Attenuation of Liver Injury
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Mouse Models of Liver Injury
4.3. Mouse Primary Hepatocyte Isolation and Culture
4.4. Real-time RT-PCR
4.5. Immunofluorescence Staining
4.6. Quantitative Analysis of Liver Fibrosis and Inflammation
4.7. Scrape Loading/Dye Transfer assay
4.8. Western Blot Analysis
4.9. ALT and AST Quantification
4.10. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Sánchez, A.; Castro, C.; Flores, D.L.; Gutiérrez, E.; Baldi, P. Gap Junction Channels of Innexins and Connexins: Relations and Computational Perspectives. Int. J. Mol. Sci. 2019, 20, 2476. [Google Scholar] [CrossRef] [PubMed]
- Chanson, M.; Watanabe, M.; O’Shaughnessy, E.M.; Zoso, A.; Martin, P.E. Connexin Communication Compartments and Wound Repair in Epithelial Tissue. Int. J. Mol. Sci. 2018, 19, 1354. [Google Scholar] [CrossRef] [PubMed]
- Oshima, A. Structure and closure of connexin gap junction channels. FEBS Lett. 2014, 588, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Decrock, E.; Cogliati, B.; Oliveira, A.G.; Marques, P.E.; Dagli, M.L.Z.; Menezes, G.B.; Mennecier, G.; Leybaert, L.; Vanhaecke, T.; et al. Connexin and pannexin (hemi)channels in the liver. Front. Physiol. 2014, 4, 405. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Watanabe, S.; Hirose, M.; Miyazaki, A.; Sato, N. Dimethylsulfoxide maintains intercellular communication by preserving the gap junctional protein connexin32 in primary cultured hepatocyte doublets from rats. J. Gastroenterol. Hepatol. 1997, 12, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Monvoisin, A.; Vix, J.; Mesnil, M.; Thuringer, D.; Debiais, F.; Cronier, L. Connexins, important players in the dissemination of prostate cancer cells. Biochim. Biophys. Acta Biomembr. 2018, 1860, 202–215. [Google Scholar] [CrossRef]
- Roy, S.; Kim, D.; Lim, R. Cell-cell communication in diabetic retinopathy. Vis. Res. 2017, 139, 115–122. [Google Scholar] [CrossRef]
- Vinken, M.; Vanhaecke, T.; Papeleu, P.; Snykers, S.; Henkens, T.; Rogiers, V. Connexins and their channels in cell growth and cell death. Cell. Signal. 2006, 18, 592–600. [Google Scholar] [CrossRef]
- Maes, M.; Cogliati, B.; Yanguas, S.C.; Willebrords, J.; Vinken, M. Roles of connexins and pannexins in digestive homeostasis. Cell. Mol. Life Sci. 2015, 72, 2809–2821. [Google Scholar] [CrossRef]
- Vinken, M.; Papeleu, P.; Snykers, S.; De Rop, E.; Henkens, T.; Chipman, J.K.; Rogiers, V.; Vanhaecke, T. Involvement of Cell Junctions in Hepatocyte Culture Functionality. Crit. Rev. Toxicol. 2006, 36, 299–318. [Google Scholar] [CrossRef]
- Hussain, A.; Das, S.S.; Babu, S.; Pal, D.; Das, S.J. Interaction of arsenic with gap junction protein connexin 43 alters gap junctional intercellular communication. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Vinken, M. Connexin-based signaling and drug-induced hepatotoxicity. J. Clin. Transl. Res. 2017, 3, 189–198. [Google Scholar] [PubMed]
- Hayashi, N.; George, J.; Shiroeda, H.; Saito, T.; Toshikuni, N.; Tsuchishima, M.; Arisawa, T.; Tsutsumi, M. Irsogladine maleate for the treatment of recurrent aphthous stomatitis in hepatitis C virus patients on pegylated-interferon and ribavirin: A pilot study. J. Gastroenterol. Hepatol. 2013, 28, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Yanguas, S.C.; Willebrords, J.; Cogliati, B.; Vinken, M. Connexin and pannexin signaling in gastrointestinal and liver disease. Transl. Res. 2015, 166, 332–343. [Google Scholar] [CrossRef]
- Nakata, Y.; Iwai, M.; Kimura, S.; Shimazu, T. Prolonged decrease in hepatic connexin32 in chronic liver injury induced by carbon tetrachloride in rats. J. Hepatol. 1996, 25, 529–537. [Google Scholar] [CrossRef]
- Miyashita, T.; Takeda, A.; Iwai, M.; Shimazu, T. Single administration of hepatotoxic chemicals transiently decreases the gap-junction-protein levels of connexin 32 in rat liver. Eur. J. Biochem. 1991, 196, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Cowles, C.; Mally, A.; Chipman, J. Different mechanisms of modulation of gap junction communication by non-genotoxic carcinogens in rat liver in vivo. Toxicology 2007, 238, 49–59. [Google Scholar] [CrossRef]
- Maes, M.; McGill, M.R.; Da Silva, T.C.; Abels, C.; Lebofsky, M.; De Araújo, C.M.M.; Tiburcio, T.; Pereira, I.V.A.; Willebrords, J.; Yanguas, S.C.; et al. Involvement of connexin43 in acetaminophen-induced liver injury. Biochim. Biophys. Acta 2016, 1862, 1111–1121. [Google Scholar] [CrossRef]
- Vinken, M.; Decrock, E.; Leybaert, L.; Bultynck, G.; Himpens, B.; Vanhaecke, T.; Rogiers, V. Non-channel functions of connexins in cell growth and cell death. Biochim. Biophys. Acta 2012, 1818, 2002–2008. [Google Scholar] [CrossRef] [PubMed]
- Berger, F.; Kubik-Huch, R.A.; Niemann, T.; Schmid, H.R.; Poetzsch, M.; Froehlich, J.M.; Beer, J.H.; Thali, M.J.; Kraemer, T. Gadolinium Distribution in Cerebrospinal Fluid after Administration of a Gadolinium-based MR Contrast Agent in Humans. Radiology 2018, 288, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Bode, F.; Katchman, A.; Woosley, R.L.; Franz, M.R. Gadolinium Decreases Stretch-Induced Vulnerability to Atrial Fibrillation. Circulation 2000, 101, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Nicolosi, A.C.; Strande, J.L.; Hsu, A.; Fu, X.; Su, J.; Gross, G.J.; Baker, J.E. Gadolinium limits myocardial infarction in the rat: Dose-response, temporal relations and mechanisms. J. Mol. Cell. Cardiol. 2008, 44, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Head, I.S.; Yeung, E.W.; Allen, D.G. Gadolinium reduces short-term stretch-induced muscle damage in isolated mdx mouse muscle fibres. J. Physiol. 2003, 552, 449–458. [Google Scholar]
- Koop, D.R.; Klopfenstein, B.; Iimuro, Y.; Thurman, R.G. Gadolinium Chloride Blocks Alcohol-Dependent Liver Toxicity in Rats Treated Chronically with Intragastric Alcohol Despite the Induction of CYP2E1. Mol. Pharmacol. 1997, 51, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Sakaida, I.; Hironaka, K.; Terai, S.; Okita, K. Gadolinium chloride reverses dimethylnitrosamine (DMN)-induced rat liver fibrosis with increased matrix metalloproteinases (MMPs) of Kupffer cells. Life Sci. 2003, 72, 943–959. [Google Scholar] [CrossRef]
- Yang, L.; Yang, L.; Dong, C.; Li, L. The class D scavenger receptor CD68 contributes to mouse chronic liver injury. Immunol. Res. 2018, 66, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Guo, W.; Fang, H.; Cao, S.; Yan, B.; Chen, S.; Zhang, K.; Zhang, S. Kupffer cell depletion by gadolinium chloride aggravates liver injury after brain death in rats. Mol. Med. Rep. 2018, 17, 6357–6362. [Google Scholar] [CrossRef]
- Harstad, E.B.; Klaassen, C.D. Gadolinium Chloride Pretreatment Prevents Cadmium Chloride-Induced Liver Damage in Both Wild-Type and MT-Null Mice. Toxicol. Appl. Pharmacol. 2002, 180, 178–185. [Google Scholar] [CrossRef]
- Hirasawa, F.; Kawagoe, M.; Wang, J.S.; Arany, S.; Zhou, X.P.; Kumagai, A.; Koizumi, Y.; Koyota, S.; Sugiyama, T. Gadolinium chloride suppresses styrene-induced cytochrome P450s expression in rat liver. Biomed. Res. 2007, 28, 323–330. [Google Scholar] [CrossRef]
- Eskandari, S.; Zampighi, G.; Leung, D.; Wright, E.; Loo, D. Inhibition of Gap Junction Hemichannels by Chloride Channel Blockers. J. Membr. Boil. 2002, 185, 93–102. [Google Scholar] [CrossRef]
- Nakashima, Y.; Ono, T.; Yamanoi, A.; El-Assal, O.N.; Kohno, H.; Nagasue, N. Expression of gap junction protein connexin32 in chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma. J. Gastroenterol. 2004, 39, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Chanson, M.; Derouette, J.P.; Roth, I.; Foglia, B.; Scerri, I.; Dudez, T.; Kwak, B.R. Gap junctional communication in tissue inflammation and repair. Biochim. Biophys. Acta 2005, 1711, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Green, L.M.; Lazarus, J.P.; LaBUE, M.; Shah, M.M. Reduced cell-cell communication in a spontaneous murine model of autoimmune thyroid disease. Endocrinology 1995, 136, 3611–3618. [Google Scholar] [CrossRef] [PubMed]
- Hu, V.W.; Xie, H.Q. Interleukin-1 alpha suppresses gap junction-mediated intercellular communication in human endothelial cells. Exp. Cell Res. 1994, 213, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Mensink, A.; De Haan, L.H.; Lakemond, C.M.; A Koelman, C.; Koeman, J.H. Inhibition of gap junctional intercellular communication between primary human smooth muscle cells by tumor necrosis factor alpha. Carcinogenesis 1995, 16, 2063–2067. [Google Scholar] [CrossRef]
- Chandross, K.J. Nerve injury and inflammatory cytokines modulate gap junctions in the peripheral nervous system. Glia 1998, 24, 21–31. [Google Scholar] [CrossRef]
- Bolaños, J.P.; Medina, J.M. Induction of nitric oxide synthase inhibits gap junction permeability in cultured rat astrocytes. J. Neurochem. 1996, 66, 2091–2099. [Google Scholar] [CrossRef]
- Temme, A.; Traub, O.; Willecke, K. Downregulation of connexin32 protein and gap-junctional intercellular communication by cytokine-mediated acute-phase response in immortalized mouse hepatocytes. Cell Tissue Res. 1998, 294, 345–350. [Google Scholar] [CrossRef]
- Graham, S.V.; Jiang, J.X.; Mesnil, M. Connexins and Pannexins: Important Players in Tumorigenesis, Metastasis and Potential Therapeutics. Int. J. Mol. Sci. 2018, 19, 1645. [Google Scholar] [CrossRef]
- Huang, L.; Guo, Y.; Cao, D.; Liu, X.; Zhang, L.; Cao, K.; Hu, T.; Qi, Y.; Xu, C. Effects of Helicobacter pylori on the expression levels of GATA-3 and connexin 32 and the GJIC function in gastric epithelial cells and their association by promoter analysis. Oncol. Lett. 2018, 16, 1650–1658. [Google Scholar] [CrossRef]
- Adding, L.C.; Bannenberg, G.L.; E Gustafsson, L. Basic experimental studies and clinical aspects of gadolinium salts and chelates. Cardiovasc. Drug Rev. 2001, 19, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, S.; Taguchi, K.; Maeda, H.; Watanabe, H.; Ishima, Y.; Otagiri, M.; Maruyama, T. Kupffer cell inactivation by carbon monoxide bound to red blood cells preserves hepatic cytochrome P450 via anti-oxidant and anti-inflammatory effects exerted through the HMGB1/TLR-4 pathway during resuscitation from hemorrhagic shock. Biochem. Pharmacol. 2015, 97, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Thinnes, F.P.; Walter, G.; Hellmann, K.P.; Hellmann, T.; Merker, R.; Kiafard, Z.; Eben-Brunnen, J.; Schwarzer, C.; Gotz, H.; Hilschmann, N. Gadolinium as an opener of the outwardly rectifying Cl(-) channel (ORCC). Is there relevance for cystic fibrosis therapy? Pflugers Arch. 2001, 443 (Suppl. 1), S111–S116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Yang, R.; Xiao, M.; Guan, M.J.; Zhao, N.; Zeng, T. Kupffer cells activation promoted binge drinking-induced fatty liver by activating lipolysis in white adipose tissues. Toxicology 2017, 390, 53–60. [Google Scholar] [CrossRef]
- Vinken, M.; Doktorova, T.; Decrock, E.; Leybaert, L.; Vanhaecke, T.; Rogiers, V. Gap junctional intercellular communication as a target for liver toxicity and carcinogenicity. Crit. Rev. Biochem. Mol. Boil. 2009, 44, 201–222. [Google Scholar] [CrossRef]
- Kang, K.S.; Kang, B.C.; Lee, B.J.; Che, J.H.; Li, G.X.; Trosko, J.E.; Lee, Y.S. Preventive effect of epicatechin and ginsenoside Rb2 on the inhibition of gap junctional intercellular communication by TPA and H2O2. Cancer Lett. 2000, 152, 97–106. [Google Scholar] [CrossRef]
- Sai, K. Prevention of the down-regulation of gap junctional intercellular communication by green tea in the liver of mice fed pentachlorophenol. Carcinogenesis 2000, 21, 1671–1676. [Google Scholar] [CrossRef]
- Yang, L.; Yue, S.; Yang, L.; Liu, X.; Han, Z.; Zhang, Y.; Li, L. Sphingosine kinase/sphingosine 1-phosphate (S1P)/S1P receptor axis is involved in liver fibrosis-associated angiogenesis. J. Hepatol. 2013, 59, 114–123. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Dong, C.; Tian, L.; Ji, X.; Yang, L.; Li, L. Gadolinium Chloride Restores the Function of the Gap Junctional Intercellular Communication between Hepatocytes in a Liver Injury. Int. J. Mol. Sci. 2019, 20, 3748. https://doi.org/10.3390/ijms20153748
Yang L, Dong C, Tian L, Ji X, Yang L, Li L. Gadolinium Chloride Restores the Function of the Gap Junctional Intercellular Communication between Hepatocytes in a Liver Injury. International Journal of Molecular Sciences. 2019; 20(15):3748. https://doi.org/10.3390/ijms20153748
Chicago/Turabian StyleYang, Le, Chengbin Dong, Lei Tian, Xiaofang Ji, Lin Yang, and Liying Li. 2019. "Gadolinium Chloride Restores the Function of the Gap Junctional Intercellular Communication between Hepatocytes in a Liver Injury" International Journal of Molecular Sciences 20, no. 15: 3748. https://doi.org/10.3390/ijms20153748
APA StyleYang, L., Dong, C., Tian, L., Ji, X., Yang, L., & Li, L. (2019). Gadolinium Chloride Restores the Function of the Gap Junctional Intercellular Communication between Hepatocytes in a Liver Injury. International Journal of Molecular Sciences, 20(15), 3748. https://doi.org/10.3390/ijms20153748