R-Loci Arrangement Versus Downy and Powdery Mildew Resistance Level: A Vitis Hybrid Survey

 , ,
, , 
Abstract
1. Introduction
2. Results and Discussion
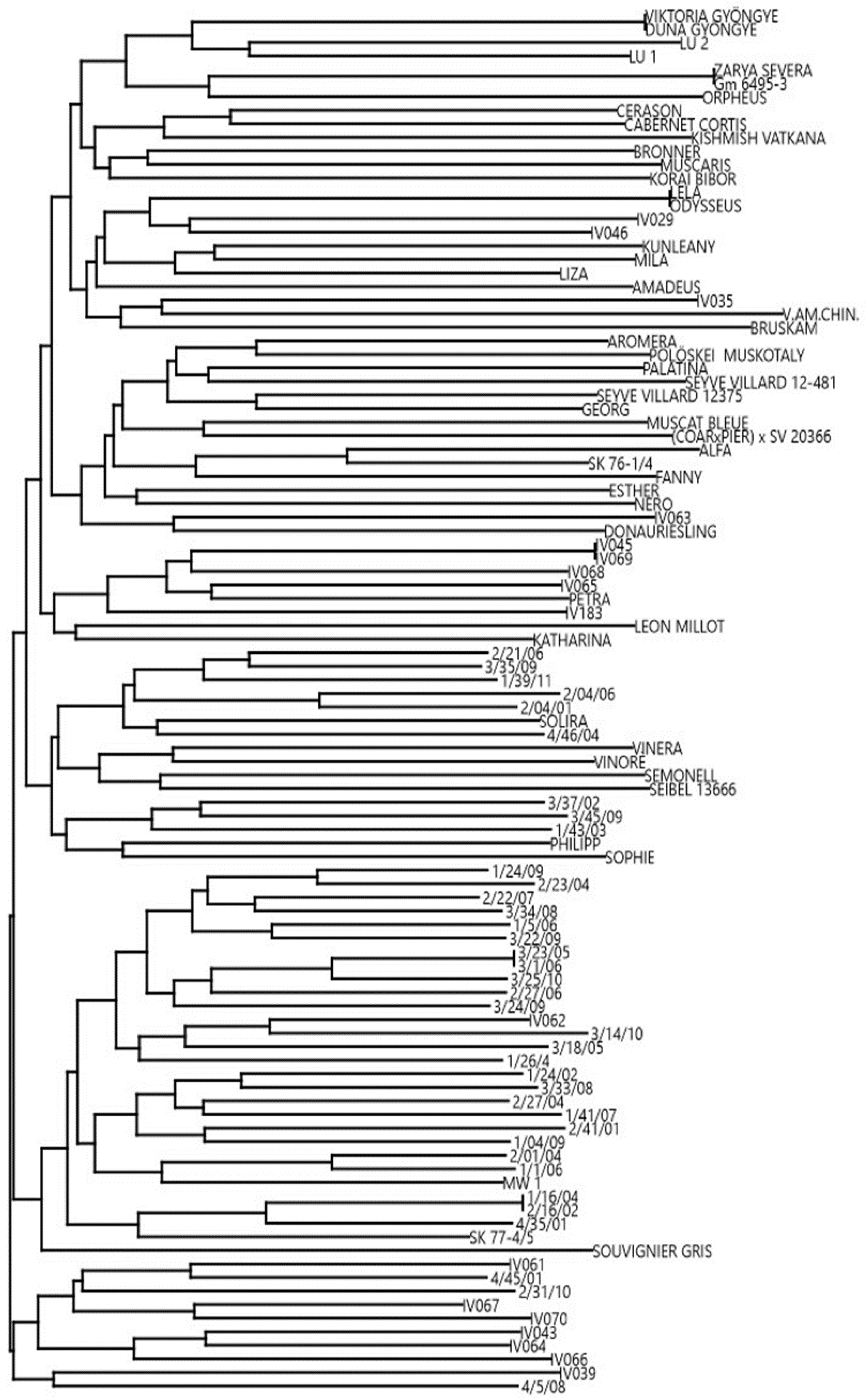
2.1. Fingerprinting
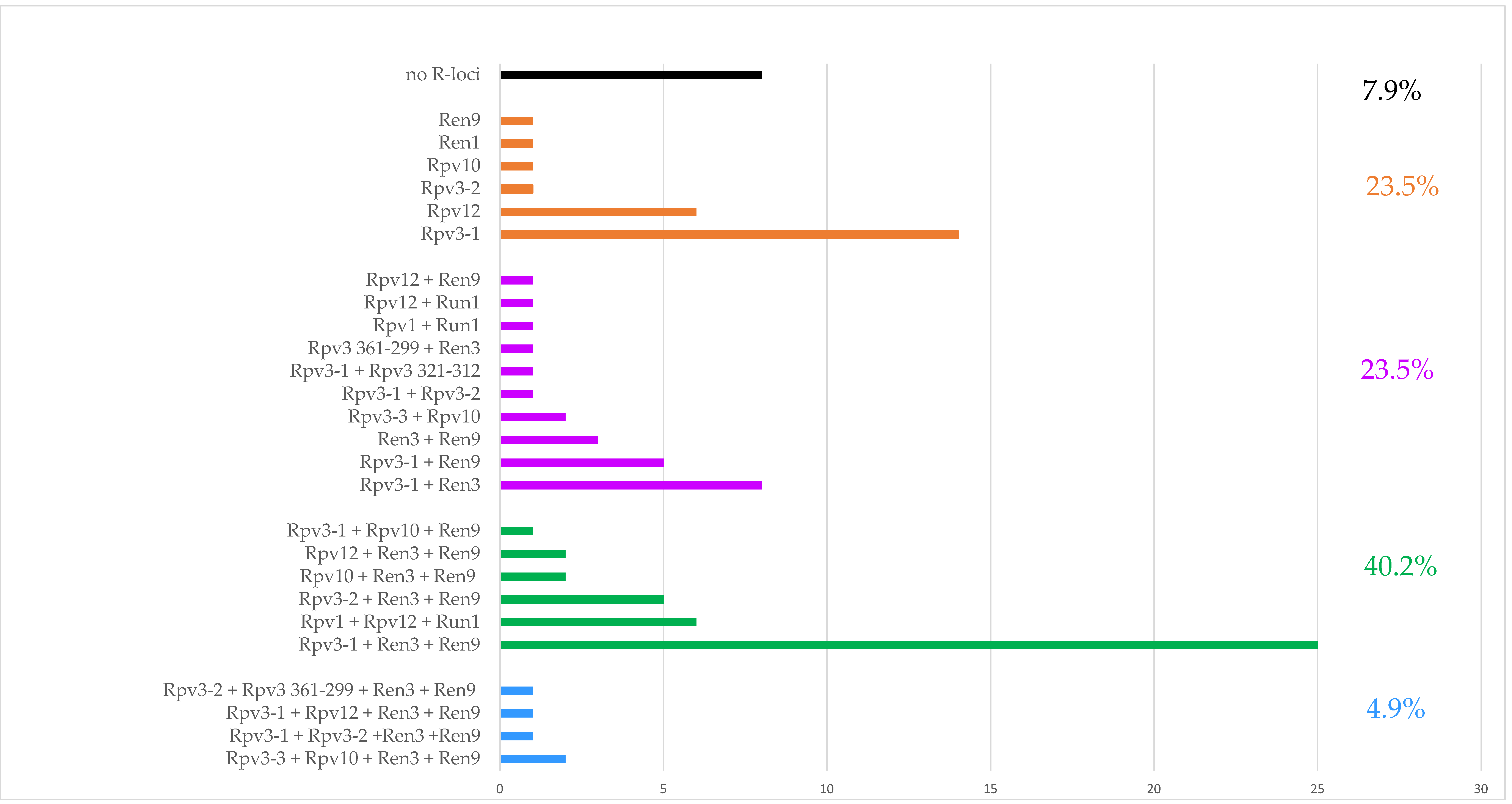
2.2. R-Loci Characterization
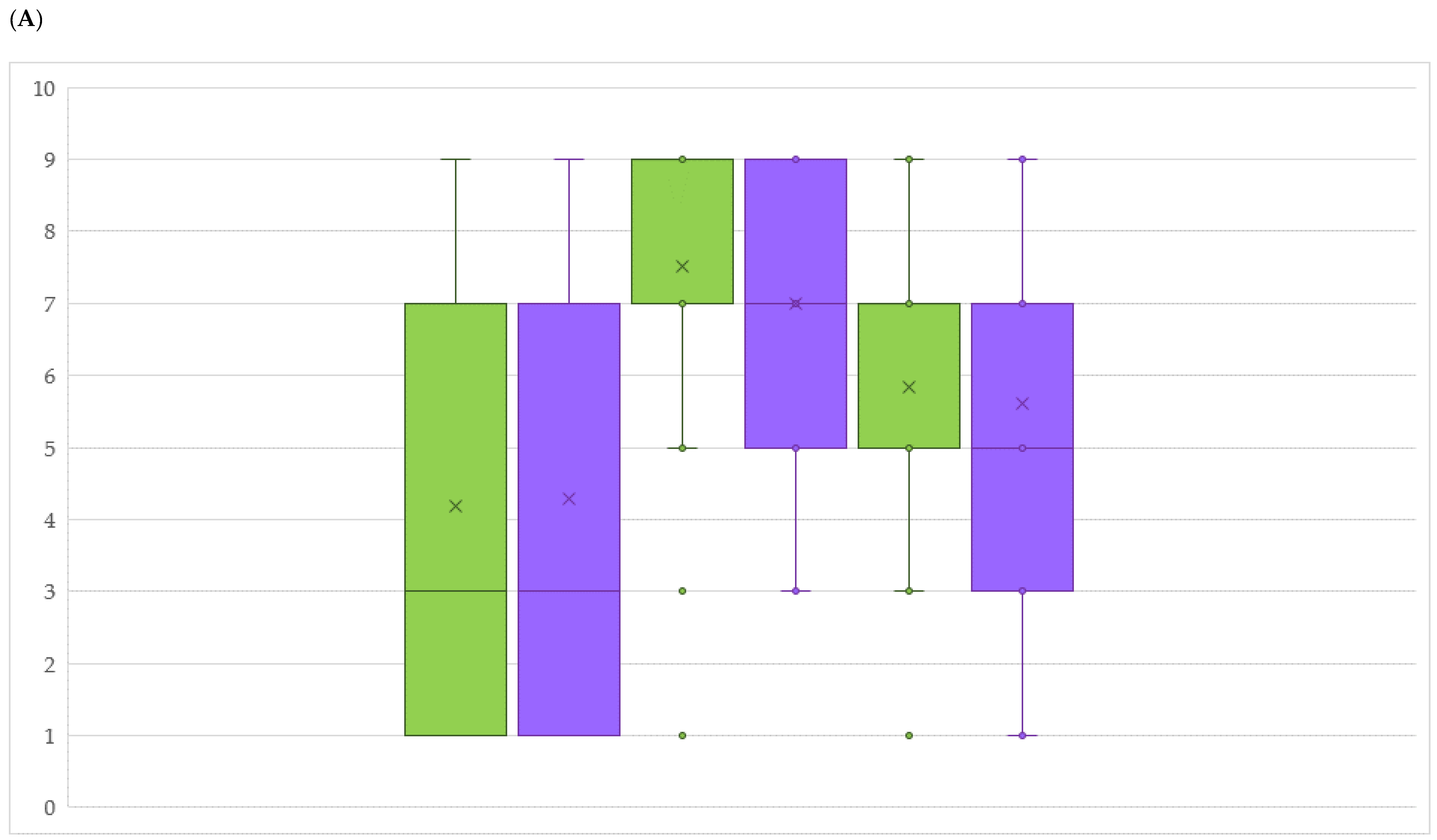
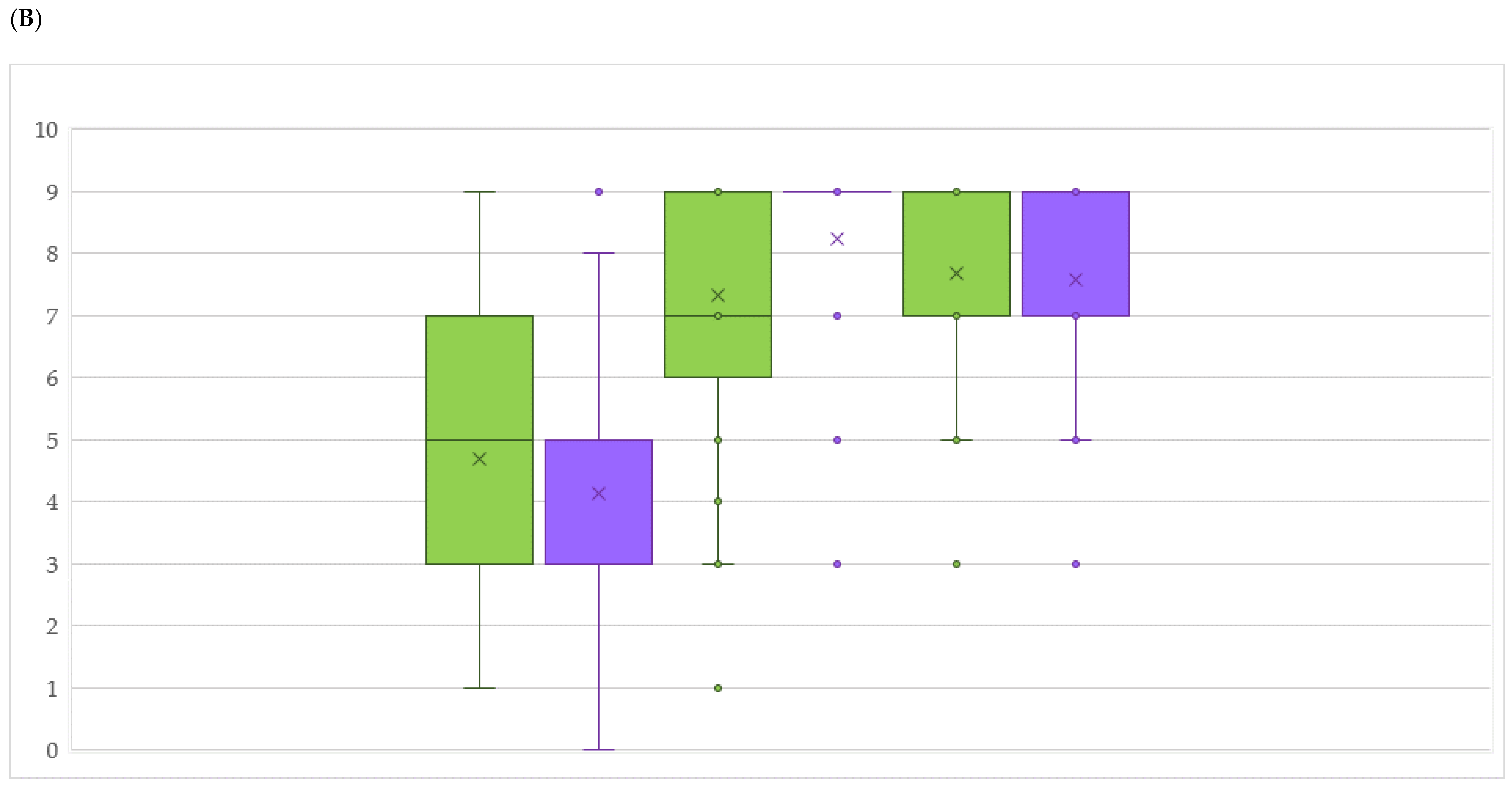
2.3. DM and PM Resistance Evaluation in an Untreated Field
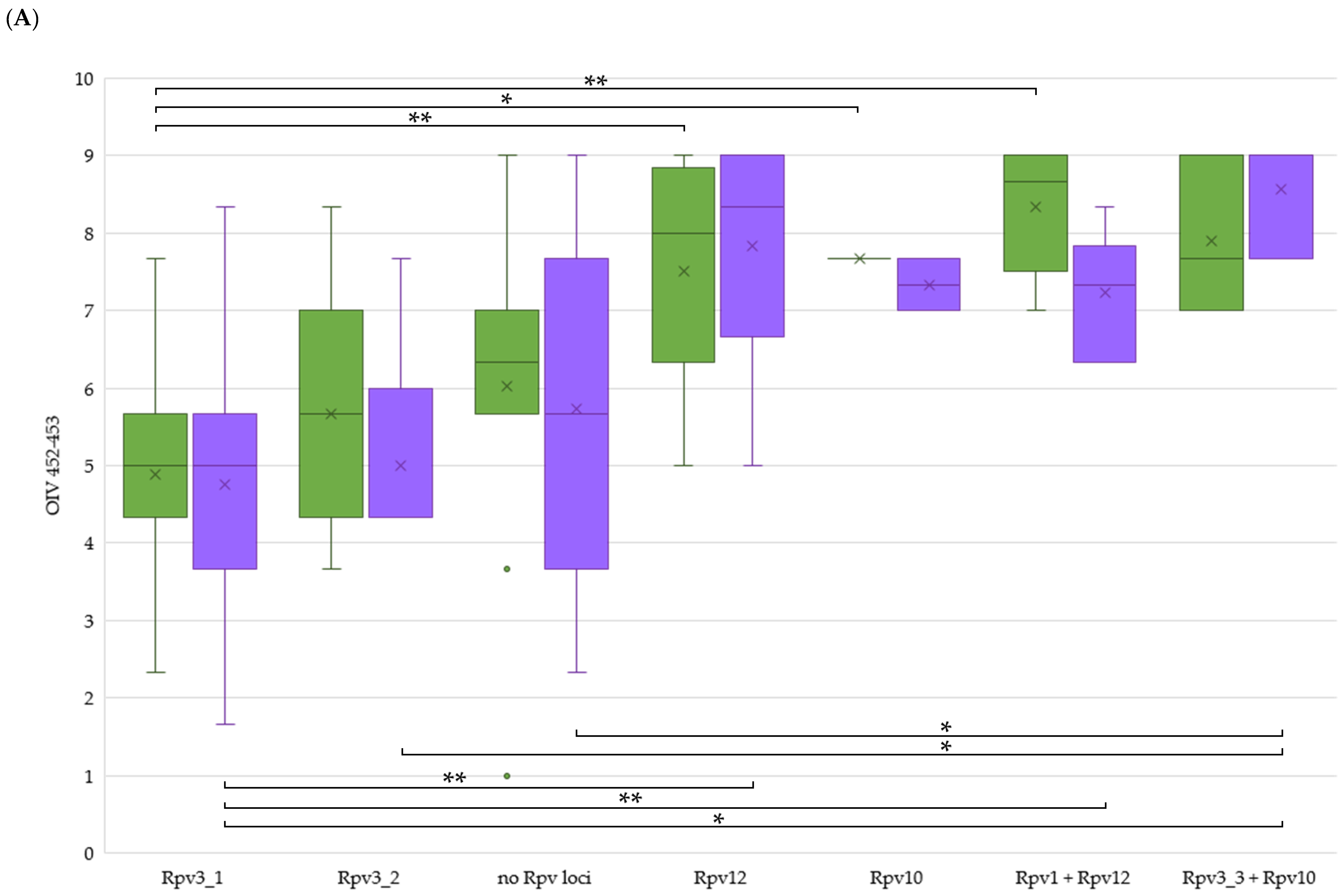
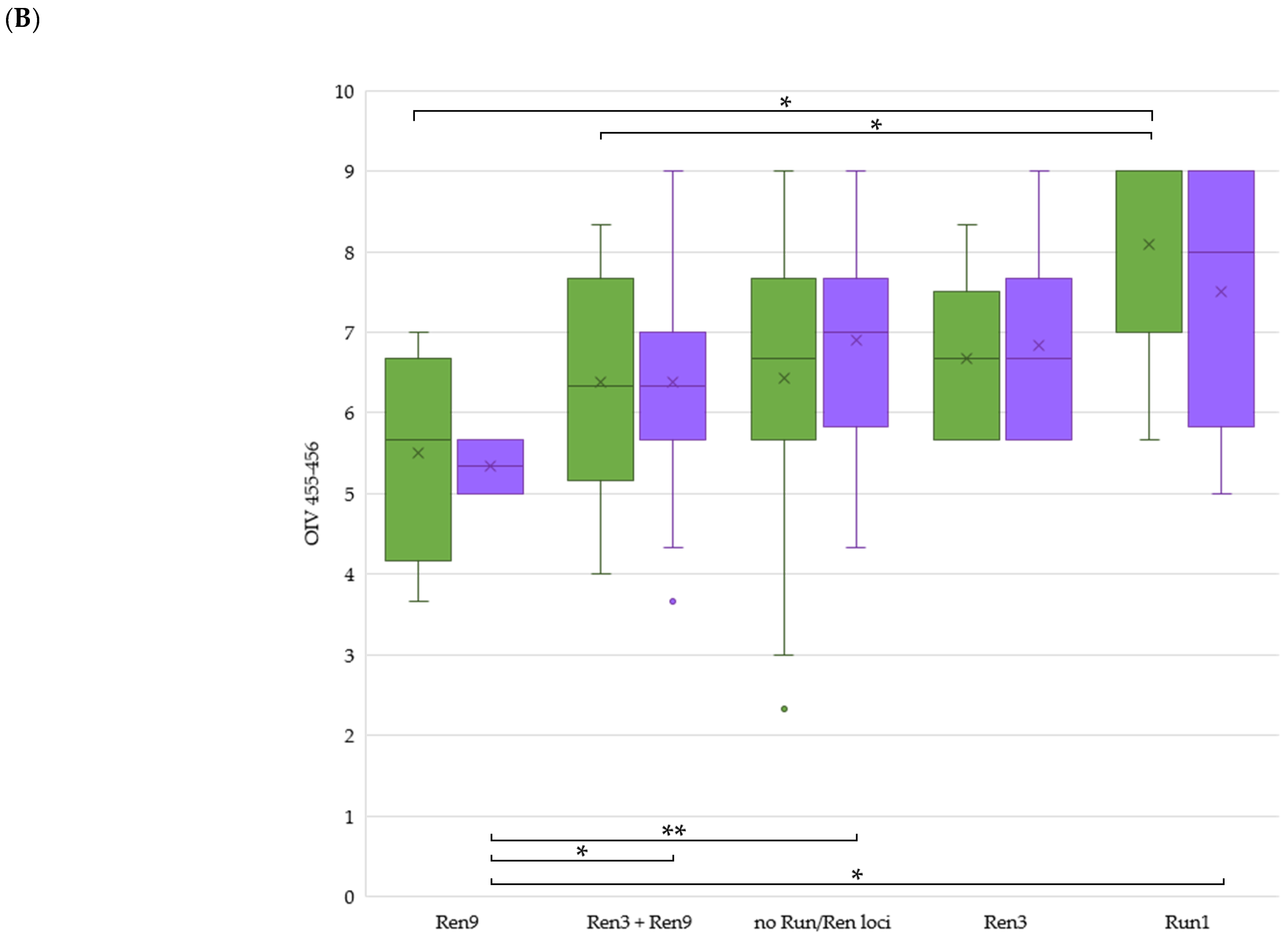
2.4. Comparison between Mildew Resistance in Untreated Field and the Arrangement of Rpv or Run/Ren Loci
3. Materials and Methods
3.1. Genetic Material and DNA Isolation
3.2. Trueness-to-Type Analysis
3.3. R-Loci Analysis
3.4. DM and PM Symptom Phenotyping in Field
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Agrios, G. Plant Pathology, 3rd ed.; Academic Press: New York, NY, USA, 1988. [Google Scholar]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Dodds, P.N.; Lawrence, G.J.; Pryor, A.; Ellis, J.G. Genetic Analysis and Evolution of Plant Disease Resistance Genes. Mol. Plant Pathol. 2000, 4, 92–112. [Google Scholar] [CrossRef]
- Table of Loci for Traits in Grapevine Relevant for Breeding and Genetics. Available online: http://www.vivc.de/docs/dataonbreeding/20181001_Table of Loci for Traits in Grapevine.pdf (accessed on 30 October 2018).
- Pauquet, J.; Bouquet, A.; This, P.; Adam-Blondon, A.F. Establishment of a local map of AFLP markers around the powdery mildew resistance gene Run1 in grapevine and assessment of their usefulness for marker assisted selection. Theor. Appl. Genet. 2001, 103, 1201–1210. [Google Scholar] [CrossRef]
- Barker, C.L.; Donald, T.; Pauquet, J.; Ratnaparkhe, M.B.; Bouquet, A.; Adam-Blondon, A.F.; Thomas, M.R.; Dry, I. Genetic and physical mapping of the grapevine powdery mildew resistance gene, Run1, using a bacterial artificial chromosome library. Theor. Appl. Genet. 2005, 111, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hausmann, L.; Eibach, R.; Welter, L.J.; Töpfer, R.; Zyprian, E.M. A framework map from grapevine V3125 (Vitis vinifera ‘Schiava grossa’ × ’Riesling’) × rootstock cultivar “Börner” (Vitis riparia × Vitis cinerea) to localize genetic determinants of phylloxera root resistance. Theor. Appl. Genet. 2009, 119, 1039–1051. [Google Scholar] [CrossRef]
- Feechan, A.; Anderson, C.; Torregrosa, L.; Jermakow, A.; Mestre, P.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; Walker, A.R.; Cadle-Davidson, L.; Reisch, B.; et al. Genetic dissection of a TIR-NB-LRR locus from the wild North American grapevine species Muscadinia rotundifolia identifies paralogous genes conferring resistance to major fungal and oomycete pathogens in cultivated grapevine. Plant. J. 2013, 76, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Maul, E.; Töpfer, R. VIVC-Vitis International Variety Catalogue. Available online: http://www.vivc.de/ (accessed on 31 May 2019).
- Alleweldt, G.; Possingham, J.V. Progress in grapevine breeding. Theor. Appl. Genet. 1988, 75, 669–673. [Google Scholar] [CrossRef]
- Raddova, J.; Stefkova, A.; Radek, S.; Baranek, M. Genetic analysis of Vitis interspecific hybrids occuring in vineyards of the Czech Republic. Pak. J. Bot. 2016, 48, 681–688. [Google Scholar]
- Töpfer, R.; Hausmann, L.; Harst, M.; Maul, E.; Zyprian, E.; Eibach, R. New Horizons for Grapevine Breeding. Fruit Veg. Cereal Sci. Biotechnol. 2011, 5, 79–100. [Google Scholar]
- Pedneault, K.; Provost, C. Fungus resistant grape varieties as a suitable alternative for organic wine production: Benefits, limits, and challenges. Sci. Hortic. (Amsterdam). 2016, 208, 57–77. [Google Scholar] [CrossRef]
- Council Regulation (EC) No. 1493/1999 of 17 May 1999 on the Common Organization of the Market in Wine. Available online: https://eur-lex.europa.eu/eli/reg/1999/1493/2009-01-20 (accessed on 28 June 2019).
- European Food Safety Authority (EFSA) Annual Report 2003. Available online: https://www.efsa.europa.eu/sites/default/files/corporate_publications/files/ar03en.pdf (accessed on 28 June 2019).
- Council Regulation (EC) No 491/2009 of 25 May 2009 amending Regulation (EC) No 1234/2007 Establishing a Common Organisation of Agricultural Markets and on Specific Provisions for Certain Agricultural Products (Single CMO Regulation). Available online: https://eur-lex.europa.eu/eli/reg/2009/491/oj (accessed on 28 June 2019).
- Sivčev, B.V.; Sivčev, I.L.; Ranković Vasić, Z.Z. Natural process and use of natural matters in organic viticulture. J. Agric. Sci. 2010, 55, 195–215. [Google Scholar] [CrossRef][Green Version]
- PIWI International. Available online: https://www.piwi-international.de/de/ (accessed on 8 May 2019).
- Peressotti, E.; Wiedemann-Merdinoglu, S.; Delmotte, F.; Bellin, D.; Di Gaspero, G.; Testolin, R.; Merdinoglu, D.; Mestre, P. Breakdown of resistance to grapevine downy mildew upon limited deployment of a resistant variety. BMC Plant Biol. 2010, 10, 147. [Google Scholar] [CrossRef] [PubMed]
- Malav, A.K.; Indu; Chandrawat, K.S. Gene Pyramiding: An Overview. Int. J. Curr. Res. Biosci. Plant Biol. 2016, 3, 22–28. [Google Scholar] [CrossRef]
- Delmotte, F.; Bourguet, D.; Franck, P.; Guillemaud, T.; Reboud, X.; Vacher, C.; Walker, A.-S. Combining Selective Pressures to Enhance the Durability of Disease Resistance Genes. Front. Plant Sci. 2016, 7, 1916. [Google Scholar] [CrossRef]
- Eibach, R.; Zyprian, E.; Welter, L.; Töpfer, R. The use of molecular markers for pyramiding resistance genes in grapevine breeding. J. Grapevine Res. 2007, 46, 120–124. [Google Scholar]
- Di Gaspero, G.; Copetti, D.; Coleman, C.; Castellarin, S.D.; Eibach, R.; Kozma, P.; Lacombe, T.; Gambetta, G.; Zvyagin, A.; Cindrić, P.; et al. Selective sweep at the Rpv3 locus during grapevine breeding for downy mildew resistance. Theor. Appl. Genet. 2012, 124, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Mahanil, S.; Ramming, D.; Cadle-Davidson, M.; Owens, C.; Garris, A.; Myles, S.; Cadle-Davidson, L. Development of marker sets useful in the early selection of Ren4 powdery mildew resistance and seedlessness for table and raisin grape breeding. Theor. Appl. Genet. 2012, 124, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.; Tenscher, A.C.; Ramming, D.W.; Walker, M.A. Using a limited mapping strategy to identify major QTLs for resistance to grapevine powdery mildew (Erysiphe necator) and their use in marker-assisted breeding. Theor. Appl. Genet. 2011, 122, 1059–1073. [Google Scholar] [CrossRef]
- Pap, D.; Riaz, S.; Dry, I.B.; Jermakow, A.; Tenscher, A.C.; Cantu, D.; Oláh, R.; Walker, M.A. Identification of two novel powdery mildew resistance loci, Ren6 and Ren7, from the wild Chinese grape species Vitis piasezkii. BMC Plant Biol. 2016, 16, 170. [Google Scholar] [CrossRef] [PubMed]
- Foria, S.; Magris, G.; Morgante, M.; Di Gaspero, G. The genetic background modulates the intensity of Rpv3-dependent downy mildew resistance in grapevine. Plant Breed. 2018, 137, 220–228. [Google Scholar] [CrossRef]
- Barba, P.; Cadle-Davidson, L.; Harriman, J.; Glaubitz, J.C.; Brooks, S.; Hyma, K.; Reisch, B. Grapevine powdery mildew resistance and susceptibility loci identified on a high-resolution SNP map. Theor. Appl. Genet. 2014, 127, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Zyprian, E.; Šimon, S.; Schwander, F.; Töpfer, R. Efficiency of Single Nucleotide Polymorphisms to improve a genetic map of complex pedigree grapevines. Vitis-J. Grapevine Res. 2015, 54, 29–32. [Google Scholar]
- Mammadov, J.; Aggarwal, R.; Buyyarapu, R.; Kumpatla, S. SNP markers and their impact on plant breeding. Int. J. Plant Genom. 2012, 2012, 728398. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fresnedo-Ramírez, J.; Wang, M.; Cote, L.; Schweitzer, P.; Barba, P.; Takacs, E.M.; Clark, M.; Luby, J.; Manns, D.C.; et al. A next-generation marker genotyping platform (AmpSeq) in heterozygous crops: A case study for marker-assisted selection in grapevine. Hortic. Res. 2016, 3, 16002. [Google Scholar] [CrossRef] [PubMed]
- Welter, L.J.; Göktürk-Baydar, N.; Akkurt, M.; Maul, E.; Eibach, R.; Töpfer, R.; Zyprian, E.M. Genetic mapping and localization of quantitative trait loci affecting fungal disease resistance and leaf morphology in grapevine (Vitis vinifera L). Mol. Breed. 2007, 20, 359–374. [Google Scholar] [CrossRef]
- van Heerden, C.J.; Burger, P.; Vermeulen, A.; Prins, R. Detection of downy and powdery mildew resistance QTL in a ‘Regent’ × ‘RedGlobe’ population. Euphytica 2014, 200, 281–295. [Google Scholar] [CrossRef]
- Bellin, D.; Peressotti, E.; Merdinoglu, D.; Wiedemann-Merdinoglu, S.; Adam-Blondon, A.-F.; Cipriani, G.; Morgante, M.; Testolin, R.; Di Gaspero, G. Resistance to Plasmopara viticola in grapevine ‘Bianca’ is controlled by a major dominant gene causing localised necrosis at the infection site. Theor. Appl. Genet. 2009, 120, 163–176. [Google Scholar] [CrossRef]
- Casagrande, K.; Falginella, L.; Castellarin, S.D.; Testolin, R.; Di Gaspero, G. Defence responses in Rpv3-dependent resistance to grapevine downy mildew. Planta 2011, 234, 1097–1109. [Google Scholar] [CrossRef]
- Zyprian, E.; Ochßner, I.; Schwander, F.; Šimon, S.; Hausmann, L.; Bonow-Rex, M.; Moreno-Sanz, P.; Grando, M.S.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; et al. Quantitative trait loci affecting pathogen resistance and ripening of grapevines. Mol. Genet. Genom. 2016, 291, 1573–1594. [Google Scholar] [CrossRef]
- Vezzulli, S.; Malacarne, G.; Masuero, D.; Vecchione, A.; Dolzani, C.; Goremykin, V.; Mehari, Z.H.; Banchi, E.; Velasco, R.; Stefanini, M.; et al. The Rpv3-3 Haplotype and Stilbenoid Induction Mediate Downy Mildew Resistance in a Grapevine Interspecific Population. Front. Plant Sci. 2019, 10, 234. [Google Scholar] [CrossRef]
- Delmotte, F.; Mestre, P.; Schneider, C.; Kassemeyer, H.-H.; Kozma, P.; Richart-Cervera, S.; Rouxel, M.; Delière, L. Rapid and multiregional adaptation to host partial resistance in a plant pathogenic oomycete: Evidence from European populations of Plasmopara viticola, the causal agent of grapevine downy mildew. Infect. Genet. Evol. 2014, 27, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Venuti, S.; Copetti, D.; Foria, S.; Falginella, L.; Hoffmann, S.; Bellin, D.; Cindrić, P.; Kozma, P.; Scalabrin, S.; Morgante, M.; et al. Historical Introgression of the Downy Mildew Resistance Gene Rpv12 from the Asian Species Vitis amurensis into Grapevine Varieties. PLoS ONE 2013, 8, e61228. [Google Scholar] [CrossRef] [PubMed]
- Schwander, F.; Eibach, R.; Fechter, I.; Hausmann, L.; Zyprian, E.; Töpfer, R. Rpv10: A new locus from the Asian Vitis gene pool for pyramiding downy mildew resistance loci in grapevine. Theor. Appl. Genet. 2012, 124, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Bouquet, A. Contribution á L’étude de L’espèce Muscadinia Rotundifolia (Michx) Small et de Ses Hybrides Avec Vitis Vinifera L.-Applications en Sélection. Dissertation. Ph.D. Thesis, University of Bordeaux, Bordeaux, France, 1983. [Google Scholar]
- Bouquet, A. Introduction dans l’espèce Vitis vinifera L. d’un caractère de résistance à l’oidium (Uncinula necator Schw. Burr.) issu de l’espèce Muscadinia rotundifolia (Michx.) Small. Vignevini 1986, 12, 141–146. [Google Scholar]
- Kozma, P.; Kiss, E.; Hoffmann, S.; Galbács, Z.; Dula, T. Using the powdery mildew resistant Muscadinia rotundifolia and Vitis vinifera ‘Kishmish vatkana’ for breeding new cultivars. Acta Hortic. 2009, 827, 559–564. [Google Scholar] [CrossRef]
- Fischer, B.M.; Salakhutdinov, I.; Akkurt, M.; Eibach, R.; Edwards, K.J.; Töpfer, R.; Zyprian, E.M. Quantitative trait locus analysis of fungal disease resistance factors on a molecular map of grapevine. Theor. Appl. Genet. 2004, 108, 501–515. [Google Scholar] [CrossRef]
- Akkurt, M.; Welter, L.; Maul, E.; Töpfer, R.; Zyprian, E. Development of SCAR markers linked to powdery mildew (Uncinula necator) resistance in grapevine (Vitis vinifera L. and Vitis sp.). Mol. Breed. 2007, 19, 103–111. [Google Scholar] [CrossRef]
- Zendler, D.; Schneider, P.; Töpfer, R.; Zyprian, E. Fine mapping of Ren3 reveals two loci mediating hypersensitive response against Erysiphe necator in grapevine. Euphytica 2017, 213, 68. [Google Scholar] [CrossRef]
- Goyal, N.; Bhatia, G.; Sharma, S.; Garewal, N.; Upadhyay, A.; Upadhyay, S.K.; Singh, K. Genome-wide characterization revealed role of NBS-LRR genes during powdery mildew infection in Vitis vinifera. Genomics 2019, in press. [Google Scholar] [CrossRef]
- Katula-Debreceni, D.; Lencsés, A.K.; Szőke, A.; Veres, A.; Hoffmann, S.; Kozma, P.; Kovács, L.G.; Heszky, L.; Kiss, E. Marker-assisted selection for two dominant powdery mildew resistance genes introgressed into a hybrid grape population. Sci. Hortic. (Amsterdam) 2010, 126, 448–453. [Google Scholar] [CrossRef]
- Di Gaspero, G.; Cattonaro, F. Application of genomics to grapevine improvement. Aust. J. Grape Wine Res. 2010, 16, 122–130. [Google Scholar] [CrossRef]
- Merdinoglu, D.; Schneider, C.; Prado, E.; Wiedemann-Merdinoglu, S.; Mestre, P. Breeding for durable resistance to downy and powdery mildew in grapevine. OENO One 2018, 52, 203–209. [Google Scholar] [CrossRef]
- Klekowski, E. Genetic load and its causes in long-lived plants. Trees 1988, 2, 195–203. [Google Scholar] [CrossRef]
- King, J.L. The gene interaction component of the genetic load. Genetics 1966, 53, 403–413. [Google Scholar] [PubMed]
- Savary, S.; Delbac, L.; Rochas, A.; Taisant, G.; Willocquet, L. Analysis of Nonlinear Relationships in Dual Epidemics, and its Application to the Management of Grapevine Downy and Powdery Mildews. Phytopathology 2009, 99, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Caffi, T.; Giosuè, S.; Bugiani, R. A mechanistic model simulating primary infections of downy mildew in grapevine. Ecol. Model. 2008, 212, 480–491. [Google Scholar] [CrossRef]
- Boso, S.; Gago, P.; Santiago, J.L.; Martínez, M.C. Variation in Sensitivity of Different Grapevine Genotypes to Erysiphe necator Growing under Unfavourable Climatic Conditions. S. Afr. J. Enol. Vitic. 2018, 39, 100–105. [Google Scholar] [CrossRef]
- Keller, M.; Rogiers, S.Y.; Schultz, H.R. Nitrogen and ultraviolet radiation modify grapevines’ susceptibility to powdery mildew Cold hardiness in grapevines View project. Vitis 2003, 42, 87–94. [Google Scholar]
- Pacifico, D.; Gaiotti, F.; Giusti, M.; Tomasi, D. Performance of interspecific grapevine varieties in north-east Italy. Agric. Sci. 2013, 4, 91–101. [Google Scholar] [CrossRef]
- Vezzulli, S.; Vecchione, A.; Stefanini, M.; Zulini, L. Downy mildew resistance evaluation in 28 grapevine hybrids promising for breeding programs in Trentino region (Italy). Eur. J. Plant Pathol. 2018, 150, 485–495. [Google Scholar] [CrossRef]
- Pavloušek, P. Evaluation of foliar resistance of grapevine genetic resources to downy mildew (Plasmopara viticola). Acta Univ. Agric. Silvic. Mendel. Brun. 2012, 60, 191–198. [Google Scholar] [CrossRef]
- Lisek, J. Yielding and healthiness of selected grape cultivars for processing in central Poland. J. Fruit Ornam. Plant Res. 2010, 18, 265–272. [Google Scholar]
- Krizsics Cziskász, A.; Kozma, P. Characterization of fungus resistant grape varieties and candidate varieties in Pécs area. Acta Hortic. 2003, 763–765. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.-L.; Moury, B.; Caffier, V.; Montarry, J.; Kerlan, M.-C.; Fournet, S.; Durel, C.-E.; Delourme, R. Quantitative Resistance to Plant Pathogens in Pyramiding Strategies for Durable Crop Protection. Front. Plant Sci. 2017, 8, 1838. [Google Scholar] [CrossRef]
- Qiu, W.; Feechan, A.; Dry, I. Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease. Hortic. Res. 2015, 2, 15020. [Google Scholar] [CrossRef]
- Fodor, A.; Segura, V.; Denis, M.; Neuenschwander, S.; Fournier-Level, A.; Chatelet, P.; Homa, F.A.A.; Lacombe, T.; This, P.; Le Cunff, L. Genome-Wide Prediction Methods in Highly Diverse and Heterozygous Species: Proof-of-Concept through Simulation in Grapevine. PLoS ONE 2014, 9, e110436. [Google Scholar] [CrossRef]
- This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.S.; Eisenheld, C.; Ferreira-Monteiro, F.; et al. Development of a standard set of microsatellite reference alleles for identification of grape cultivars. Theor. Appl. Genet. 2004, 109, 1448–1458. [Google Scholar] [CrossRef]
- Maul, E.; Sudharma, K.N.; Kecke, S.; Marx, G.; Müller, C.; Audeguin, L.; Boselli, M.; Boursiquot, J.M.; Bucchetti, B.; Cabello, F.; et al. The European Vitis Database (www.eu-vitis.de): A technical innovation through an online uploading and interactive modification system. Vitis 2012, 51, 79–85. [Google Scholar]
- Migliaro, D.; Peressotti, E.; Dolzani, C.; Bettinelli, P.; Banchi, E.; Riaz, S.; Cappellin, L.; Zini, E.; Walker, M.A.; Reisch, B.I.; et al. Unravelling the knot of grapevine hybrid kinship: A 50 SSR- and 12 R-loci-based genetic diversity analysis. in preparation.
- Vezzulli, S.; Dolzani, C.; Migliaro, D.; Banchi, E.; Stedile, T.; Zatelli, A.; Dallaserra, M.; Clementi, S.; Dorigatti, C.; Velasco, R.; et al. The FEM grapevine breeding program for downy and powdery mildew resistances: Towards a green viticulture. In Proceedings of the XII International Conference on Grapevine Breeding and Genetics, Bordeaux, France, 15–20 July 2018; Volume 2018, p. 37. [Google Scholar]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Name | InnoVitis Code | TTT | Rpv1 | Rpv3-1 299-279 | Rpv3-2 null-297 | Rpv3-3 null-271 | Rpv3 321-312 | Rpv3 361-299 | Rpv3 299-314 | Rpv3 null-287 | Rpv10 | Rpv12 | Run1 | Ren1 | Ren3 | Ren9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1/04/09 | IV130 | not available | Ren3 | Ren9 | ||||||||||||
| 1/1/06 | IV109 | not available | Rpv3-1 299-279 | Ren3 | ||||||||||||
| 1/16/04 | IV090 | not available | Rpv3-1 299-279 | Ren9 | ||||||||||||
| 1/24/02 | IV091 | not available | Rpv3-1 299-279 | Ren9 | ||||||||||||
| 1/24/09 | IV097 | not available | Rpv3-1 299-279 | Ren3 | ||||||||||||
| 1/26/4 | IV132 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 1/39/11 | IV119 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 1/41/07 | IV104 | not available | Rpv3-1 299-279 | Ren3 | ||||||||||||
| 1/43/03 | IV101 | not available | Rpv3-1 299-279 | |||||||||||||
| 1/5/06 | IV112 | not available | Rpv3-2 null-297 | Ren3 | Ren9 | |||||||||||
| 2/01/04 | IV102 | not available | Rpv3-1 299-279 | Ren3 | ||||||||||||
| 2/04/01 | IV131 | not available | Rpv3-2 null-297 | Ren3 | Ren9 | |||||||||||
| 2/04/06 | IV108 | not available | Rpv3-1 299-279 | |||||||||||||
| 2/16/02 | IV117 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 2/21/06 | IV088 | not available | Rpv3-1 299-279 | |||||||||||||
| 2/22/07 | IV078 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 2/23/04 | IV120 | not available | Rpv3-1 299-279 | |||||||||||||
| 2/27/04 | IV074 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 2/27/06 | IV077 | not available | Rpv3-2 null-297 | Ren3 | Ren9 | |||||||||||
| 2/31/10 | IV098 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 2/41/01 | IV086 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/1/06 | IV076 | not available | Rpv3-1 299-279 | |||||||||||||
| 3/14/10 | IV096 | not available | Rpv3-1 299-279 | Ren9 | ||||||||||||
| 3/18/05 | IV085 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/22/09 | IV133 | not available | Rpv3-1 299-279 | |||||||||||||
| 3/23/05 | IV075 | not available | Rpv3-1 299-279 | |||||||||||||
| 3/24/09 | IV125 | not available | Rpv3-1 299-279 | |||||||||||||
| 3/25/10 | IV083 | not available | Rpv3-1 299-279 | |||||||||||||
| 3/33/08 | IV114 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/34/08 | IV107 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/35/09 | IV106 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/37/02 | IV072 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| 3/45/09 | IV134 | not available | Rpv3-1 299-279 | |||||||||||||
| 4/35/01 | IV129 | not available | Ren3 | Ren9 | ||||||||||||
| 4/45/01 | IV126 | not available | Rpv3-1 299-279 | Ren9 | ||||||||||||
| 4/46/04 | IV100 | not available | Rpv3-1 299-279 | |||||||||||||
| 4/5/08 | IV110 | not available | Rpv3-1 299-279 | |||||||||||||
| ALFA | IV042 | putative | Rpv3-1 299-279 | |||||||||||||
| AMADEUS | IV184 | putative | ||||||||||||||
| V. AMURENSIS CHINENSIS | IV055 | putative | Rpv10 | |||||||||||||
| AROMERA | IV004 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| BRONNER | IV001 | yes | Rpv3-3 null-271 | Rpv10 | Ren3 | Ren9 | ||||||||||
| BRUSKAM | IV038 | putative | ||||||||||||||
| CABERNET CORTIS | IV051 | putative | Rpv3-3 null-271 | Rpv10 | Ren3 | Ren9 | ||||||||||
| CERASON | IV028 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| (COARNA N. × PIERELLE) × SV 20366 | IV037 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| DONAURIESLING | IV185 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| DUNA GYÖNGYE | IV032 | no | Rpv3-1 299-279 | Ren3 | ||||||||||||
| ESTHER | IV013 | yes | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| FANNY | IV011 | yes | Rpv3 361-299 | Ren3 | ||||||||||||
| GEORG | IV163 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| GM 6495-3 | IV213 | yes | Rpv3-3 null-271 | Rpv10 | ||||||||||||
| InnoVitis 029 | IV029 | not available | Rpv12 | Ren9 | ||||||||||||
| InnoVitis 035 | IV035 | not available | ||||||||||||||
| InnoVitis 039 | IV039 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| InnoVitis 043 | IV043 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 045 | IV045 | not available | Rpv12 | Ren3 | Ren9 | |||||||||||
| InnoVitis 046 | IV046 | not available | Rpv3-1 299-279 | Rpv12 | Ren3 | Ren9 | ||||||||||
| InnoVitis 061 | IV061 | not available | Rpv12 | Run1 | ||||||||||||
| InnoVitis 062 | IV062 | not available | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| InnoVitis 063 | IV063 | not available | ||||||||||||||
| InnoVitis 064 | IV064 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 065 | IV065 | not available | Rpv1 | Run1 | ||||||||||||
| InnoVitis 066 | IV066 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 067 | IV067 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 068 | IV068 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 069 | IV069 | not available | Rpv12 | Ren3 | Ren9 | |||||||||||
| InnoVitis 070 | IV070 | not available | Rpv1 | Rpv12 | Run1 | |||||||||||
| InnoVitis 183 | IV183 | not available | ||||||||||||||
| KATHARINA | IV164 | yes | Rpv3-1 299-279 | Rpv3-2 null-297 | ||||||||||||
| KISHMISH VATKANA | IV034 | yes | Ren1 | |||||||||||||
| KORAI BIBOR | IV186 | putative | Ren9 | |||||||||||||
| KUNLEANY | IV030 | yes | Rpv12 | |||||||||||||
| LELA | IV187 | putative | Rpv12 | |||||||||||||
| LEON MILLOT | IV021 | yes | ||||||||||||||
| LIZA | IV188 | putative | Rpv12 | |||||||||||||
| LU 1 | IV040 | putative | Rpv3-1 299-279 | Rpv10 | Ren9 | |||||||||||
| LU 2 | IV036 | putative | Rpv10 | Ren3 | Ren9 | |||||||||||
| MILA | IV189 | putative | Rpv12 | |||||||||||||
| MUSCARIS | IV008 | putative | Rpv10 | Ren3 | Ren9 | |||||||||||
| MUSCAT BLEU | IV010 | yes | Rpv3-1 299-279 | Rpv3 321-312 | ||||||||||||
| MW 1 | IV027 | putative | Rpv3-1 299-279 | Rpv3-2 null-297 | Ren3 | Ren9 | ||||||||||
| NERO | IV016 | yes | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| ODYSSEUS | IV190 | no | Rpv12 | |||||||||||||
| ORPHEUS | IV191 | yes | ||||||||||||||
| PALATINA | IV009 | yes | Rpv3-1 299-279 | |||||||||||||
| PETRA | IV195 | yes | Rpv12 | |||||||||||||
| PHILIPP | IV161 | yes | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| PÖLÖSKEI MUSKOTALY | IV012 | yes | Rpv3-1 299-279 | Ren3 | ||||||||||||
| SEIBEL 13666 | IV053 | yes | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| SEMONELL | IV041 | not available | ||||||||||||||
| SEYVE VILLARD 12375 | IV047 | yes | Rpv3-1 299-279 | Rpv3 361-299 | Ren3 | Ren9 | ||||||||||
| SEYVE VILLARD 12-481 | IV054 | putative | Rpv3-1 299-279 | Ren9 | ||||||||||||
| SK 76-1/4 | IV196 | putative | Rpv3-2 null-297 | Ren3 | Ren9 | |||||||||||
| SK 77-4/5 | IV197 | putative | Ren3 | Ren9 | ||||||||||||
| SOLIRA | IV023 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| SOPHIE | IV162 | yes | Rpv3-1 299-279 | Ren3 | ||||||||||||
| SOUVIGNIER GRIS | IV044 | putative | Rpv3-2 null-297 | Ren3 | Ren9 | |||||||||||
| VIKTORIA GYÖNGYE | IV031 | no | Rpv3-1 299-279 | Ren3 | ||||||||||||
| VINERA | IV017 | putative | Rpv3-2 null-297 | |||||||||||||
| VINORÈ | IV025 | putative | Rpv3-1 299-279 | Ren3 | Ren9 | |||||||||||
| ZARYA SEVERA | IV165 | no | Rpv3-3 null-271 | Rpv10 |
| Symbol | Resistance Trait | Chromosome | Associated Marker | Resistant Alleles/Haplotype | Reference Genotype |
|---|---|---|---|---|---|
| Rpv1 | Plasmopara viticola | 12 | VMC1g3.2 | 122 | VRH30-82-1-42 |
| VMC8g09 | 160 | ||||
| Rpv3-1 | Plasmopara viticola | 18 | UDV305 | 299 | Villard Blanc |
| (= Rpv3 299-279) | UDV737 | 279 | |||
| Rpv3-2 | UDV305 | null | Seyval | ||
| (= Rpv3 null-297) | UDV737 | 297 | |||
| Rpv3321-312 | UDV305 | 321 | Chancellor | ||
| UDV737 | 312 | ||||
| Rpv3-3(= Rpv3null-271) | UDV305 | null | Seyval | ||
| UDV737 | 271 | ||||
| Rpv3361-299 | UDV305 | 361 | Villard Blanc | ||
| UDV737 | 299 | ||||
| Rpv3299-314 | UDV305 | 299 | Couderc 13 | ||
| UDV737 | 314 | ||||
| Rpv3null-287 | UDV305 | null | Chancellor | ||
| UDV737 | 287 | ||||
| Rpv10 | Plasmopara viticola | 9 | GF09-44 | 230 | Severnyi |
| GF09-46 | 416 | ||||
| GF09-47 | 299 | ||||
| Rpv12 | Plasmopara viticola | 14 | UDV340 | 197 | Zarya Severa |
| UDV345 | 236 | ||||
| UDV 360 | 227 | ||||
| Rpv14 | Plasmopara viticola | 5 | GF05-13 | 228 | Börner |
| UDV111 | 114 | ||||
| Run1 | Erysiphe necator | 12 | VMC4f3.1 | 186 | VRH3082-1-42 |
| VMC8g9 | 160 | ||||
| Sc34-8 | 216 | ||||
| Sc35-2 | 238 | ||||
| Run2.1 | Erysiphe necator | 18 | VMC7f2 | 193 | Magnolia |
| UDV108 | 202 | ||||
| Run2.2 | Erysiphe necator | 18 | VMC7f2 | 195 | Trayshed |
| UDV108 | 220 | ||||
| Ren1 | Erysiphe necator | 13 | Sc47-18 | 249 | Kishmish vatkana |
| SC08_0071_014 | 143 | ||||
| Ren3 | Erysiphe necator | 15 | GF15-42 | 199 | Regent |
| ScORGF15-02 | 242 | ||||
| Ren9 | Erysiphe necator | 15 | CenGen6 | 287 | Regent |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zini, E.; Dolzani, C.; Stefanini, M.; Gratl, V.; Bettinelli, P.; Nicolini, D.; Betta, G.; Dorigatti, C.; Velasco, R.; Letschka, T.; et al. R-Loci Arrangement Versus Downy and Powdery Mildew Resistance Level: A Vitis Hybrid Survey. Int. J. Mol. Sci. 2019, 20, 3526. https://doi.org/10.3390/ijms20143526
Zini E, Dolzani C, Stefanini M, Gratl V, Bettinelli P, Nicolini D, Betta G, Dorigatti C, Velasco R, Letschka T, et al. R-Loci Arrangement Versus Downy and Powdery Mildew Resistance Level: A Vitis Hybrid Survey. International Journal of Molecular Sciences. 2019; 20(14):3526. https://doi.org/10.3390/ijms20143526
Chicago/Turabian StyleZini, Elena, Chiara Dolzani, Marco Stefanini, Verena Gratl, Paola Bettinelli, Daniela Nicolini, Giulia Betta, Cinzia Dorigatti, Riccardo Velasco, Thomas Letschka, and et al. 2019. "R-Loci Arrangement Versus Downy and Powdery Mildew Resistance Level: A Vitis Hybrid Survey" International Journal of Molecular Sciences 20, no. 14: 3526. https://doi.org/10.3390/ijms20143526
APA StyleZini, E., Dolzani, C., Stefanini, M., Gratl, V., Bettinelli, P., Nicolini, D., Betta, G., Dorigatti, C., Velasco, R., Letschka, T., & Vezzulli, S. (2019). R-Loci Arrangement Versus Downy and Powdery Mildew Resistance Level: A Vitis Hybrid Survey. International Journal of Molecular Sciences, 20(14), 3526. https://doi.org/10.3390/ijms20143526