Hypoxia Modulates Effects of Fatty Acids on NES2Y Human Pancreatic β-cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Effect of Hypoxia and Fatty Acids on HIF1α Expression
2.2. Modulation of the Effects of Fatty Acids on Cell Growth and Viability by Hypoxia
2.3. Modulation of the Effect of Fatty Acids on the Activation of Caspases by Hypoxia
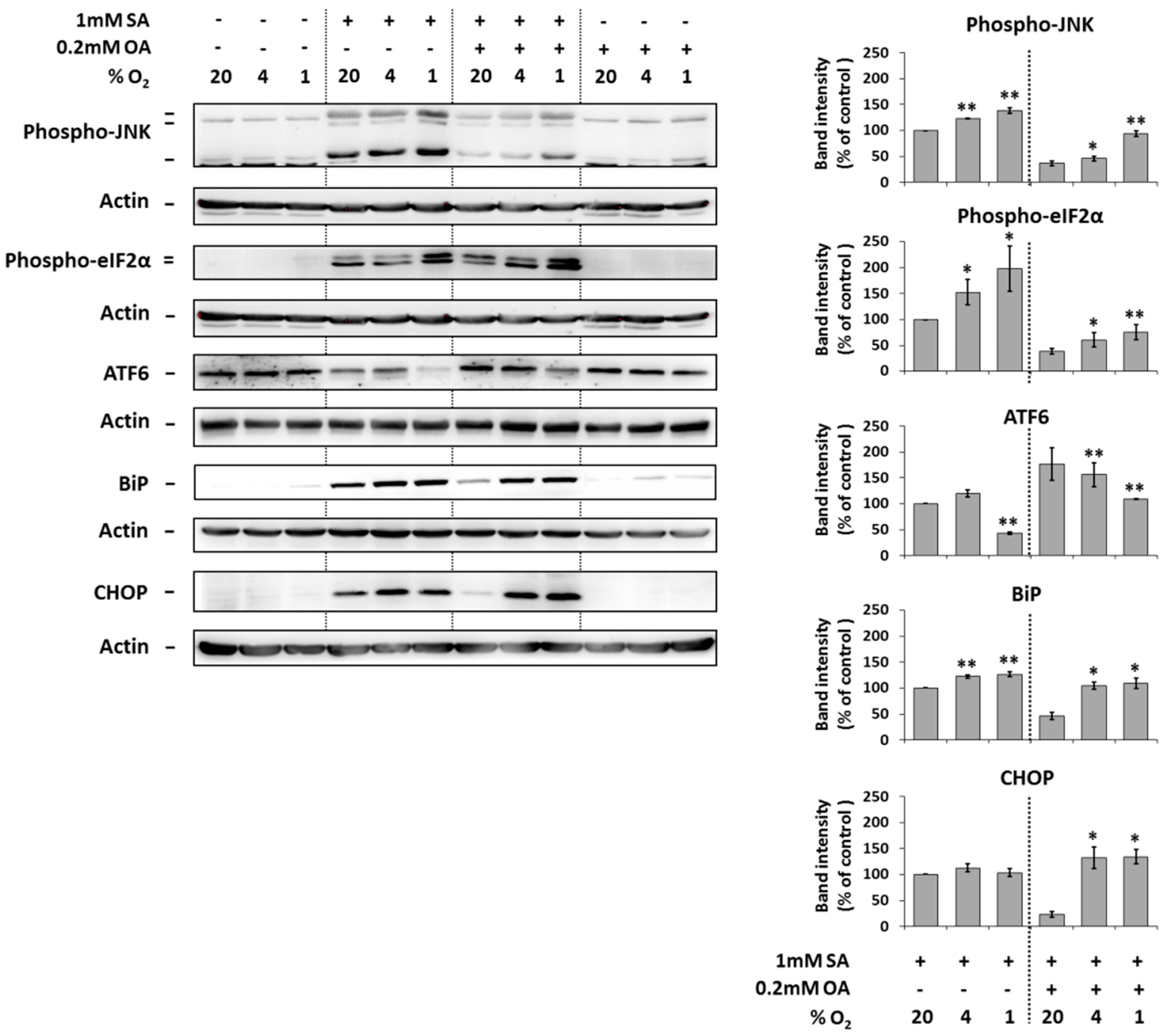
2.4. Modulation of the Effect of Fatty Acids on ER Stress Signaling by Hypoxia
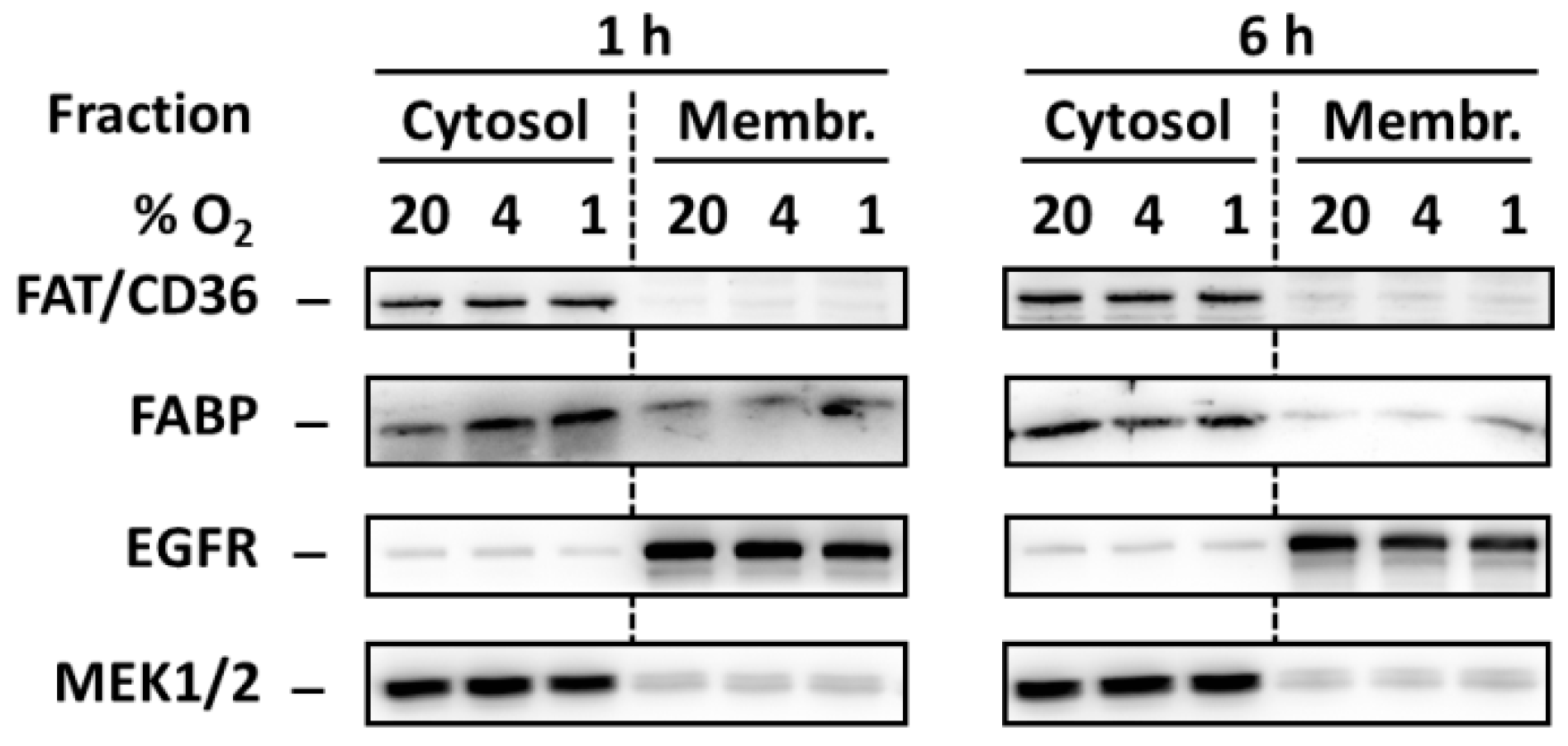
2.5. Effect of Hypoxia on Redistribution of FA Transporters FAT/CD36 and FABP
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cells and Culture Conditions
4.3. Assessment of the Effect of Hypoxia on Cell Growth and Viability
4.4. Western Blot Analysis
4.5. Preparation of Membrane-Enriched Fractions
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ATF6 | Activating transcription factor 6 |
| BiP | Immunoglobulin heavy chain-binding protein |
| CHOP | CCAAT-enhancer-binding protein (C/EBP) homologous protein |
| ER | Endoplasmic reticulum |
| FABP | Fatty acid-binding protein |
| FAT/CD36 | Fatty acid translocase/cluster of differentiation 36 |
| FAs | Fatty acids |
| HIF1α | Hypoxia-inducible factor 1α |
| IRE1α | Inositol-requiring protein 1α |
| JNK | c-Jun N-terminal kinase |
| PARP | Poly ADP-ribose polymerase |
| PERK | Protein kinase RNA (PKR)-like ER kinase |
| SA | Stearic acid |
| SAS | Sleep apnea syndrome |
| T2DM | Type 2 diabetes mellitus |
| OA | Oleic acid |
References
- Fürstova, V.; Kopska, T.; James, R.F.; Kovar, J. Comparison of the effect of individual saturated and unsaturated fatty acids on cell growth and death induction in the human pancreatic β-cell line NES2Y. Life Sci. 2008, 82, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Welters, H.J.; Tadayyon, M.; Scarpello, J.H.; Smith, S.A.; Morgan, N.G. Mono-unsaturated fatty acids protect against beta-cell apoptosis induced by saturated fatty acids, serum withdrawal or cytokine exposure. FEBS Lett. 2004, 560, 103–108. [Google Scholar] [CrossRef]
- Diakogiannaki, E.; Welters, H.J.; Morgan, N.G. Differential regulation of the endoplasmic reticulum stress response in pancreatic beta-cells exposed to long-chain saturated and monounsaturated fatty acids. J. Endocrinol. Jun 2008, 197, 553–563. [Google Scholar] [CrossRef]
- Nemcova-Fürstova, V.; James, R.F.L.; Kovar, J. Inhibitory effect of unsaturated fatty acids on saturated fatty acids-induced apoptosis in human pancreatic β-cells: activation of caspases and ER stress induction. Cell. Physiol. Biochem. 2011, 27, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Brandhorst, H.; Hossain, H.; Bierhaus, A.; Chen, C.; Bretzel, R.G.; Linn, T. Activation of NFκB dependent apoptotic pathway in pancreatic islet cells by hypoxia. Islets 2009, 1, 19–25. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zheng, X.; Zheng, X.; Wang, X.; Ma, Z.; Sunkari, G.V.; Botusan, I.; Takeda, T.; Björklund, A.; Inoue, M.; Catrina, S.B.; et al. Strong hypoxia induces apoptosis of pancreatic beta-cell by activation of the unfolded protein response and upregulation of CHOP. Cell Death Dis. 2012, 3, e322. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, Q.; Tan, J.; Li, L.; An, X.; Lei, P. Intermittent hypoxia-induced rat pancreatic β-cell apoptosis and protective effects of antioxidant intervention. Nutr. Diabetes 2014, 4, e131. [Google Scholar] [CrossRef] [PubMed]
- Jonas, J.C.; Sharma, A.; Hasenkamp, W.; Ilkova, H.; Patanè, G.; Laybutt, R.; Bonner-Weir, S.; Weir, G.C. Chronic hyperglycemia triggers loss of pancreatic b cell differentiation in an animal model of diabetes. J. Biol. Chem. 1999, 274, 14112–14121. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Meshinchi, S.; Dias-Leme, C.; Raffin, D.; Johnson, J.D.; Treutelaar, M.K.; Burant, C.F. Islet microvasculature in islet hyperplasia and failure in a model of type 2 diabetes. Diabetes 2006, 55, 2965–2973. [Google Scholar] [CrossRef]
- Valencia-Flores, M.; Orea, A.; Castano, V.A.; Resendiz, M.; Rosales, M.; Rebollar, V.; Santiago, V.; Gallegos, J.; Campos, R.M.; Gonzalez, J.; et al. Prevalence of sleep apnea and electrocardiographic disturbances in morbidly obese patients. Obes. Res. 2000, 8, 262–269. [Google Scholar] [CrossRef]
- Lecomte, P.; Criniere, L.; Fagot-Campagna, A.; Druet, C.; Fuhrman, C. Under diagnosis of obstructive sleep apnoea syndrome in patients with type 2 diabetes in France: ENTRED 2007. Diabetes Metab. 2013, 39, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Heffner, J.E.; Rozenfeld, Y.; Kai, M.; Stephens, E.A.; Brown, L.K. Prevalence of diagnosed sleep apnea among patients with type 2 diabetes in primary care. Chest 2012, 141, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Fredheim, J.M.; Rollheim, J.; Omland, T.; Hofsø, D.; Røislien, J.; Vegsgaard, K.; Hjelmesæth, J. Type 2 diabetes and pre-diabetes are associated with obstructive sleep apnea in extremely obese subjects: A cross-sectional study. Cardiovasc. Diabetol. 2011, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Biden, T.J.; Boslem, E.; Chu, K.Y.; Sue, N. Lipotoxic endoplasmic reticulum stress, β cell failure, and type 2 diabetes mellitus. Trends Endocrinol. Metab. 2014, 25, 1043–2760. [Google Scholar] [CrossRef] [PubMed]
- Bensellam, M.; Duvillie, B.; Rybachuk, G.; Laybutt, D.R.; Magnan, C.; Guiot, Y.; Pouysségur, J.; Jonas, J.C. Glucoseinduced O2 consumption activates hypoxia inducible factors 1 and 2 in rat insulin-secreting pancreatic beta-cells. PLoS ONE 2012, 7, e29807. [Google Scholar]
- Hetz, C.; Martinon, F.; Rodriguez, D.; Glimcher, L.H. The unfolded protein response: integrating stress signals through the stress sensor IRE1α. Physiol. Rev. 2011, 91, 1219–1243. [Google Scholar] [CrossRef]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef]
- Chabowski, A.; Górski, J.; Calles-Escandon, J.; Tandon, N.N.; Bonen, A. Hypoxia-induced fatty acid transporter translocation increases fatty acid transport and contributes to lipid accumulation in the heart. FEBS Lett. 2006, 580, 3617–3623. [Google Scholar] [CrossRef]
- Hyder, A.; Zenhom, M.; Klapper, M.; Herrmann, J.; Schrezenmeir, J. Expression of fatty acid binding proteins 3 and 5 genes in rat pancreatic islets and INS-1E cells: Regulation by fatty acids and glucose. Islets 2010, 2, 174–184. [Google Scholar] [CrossRef]
- Dalgaard, L.T.; Thams, P.; Gaarn, L.W.; Jensen, J.; Lee, Y.C.; Nielsen, J.H. Suppression of FAT/CD36 mRNA by human growth hormone in pancreatic beta-cells. Biochem. Biophys. Res. Commun. 2011, 410, 345–350. [Google Scholar] [CrossRef]
- Walford, G.A.; Moussignac, R.L.; Scribner, A.W.; Loscalzoand, J.; Leopold, J.A. Hypoxia Potentiates Nitric Oxide-mediated Apoptosis in Endothelial Cells via Peroxynitrite-induced Activation of Mitochondria-dependent and -independent Pathways. J. Biol. Chem. 2004, 279, 4425–4432. [Google Scholar] [CrossRef] [PubMed]
- Bullova, P.; Cougnoux, A.; Abunimer, L.; Kopacek, J.; Pastorekova, S.; Pacak, K. Hypoxia potentiates the cytotoxic effect of piperlongumine in pheochromocytoma models. Oncotarget 2016, 7, 40531–40545. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Inoue, M.; Yoshizawa, T.; Yamagata, K. Moderate hypoxia induces β-cell dysfunction with HIF-1-independent gene expression changes. PLoS ONE 2014, 9, e114868. [Google Scholar] [CrossRef] [PubMed]
- Qiao, N.; Xu, C.; Zhu, Y.X.; Cao, Y.; Liu, D.C.; Han, X. Ets-1 as an early response gene against hypoxia-induced apoptosis in pancreatic β-cells. Cell Death Dis. 2015, 6, e1650. [Google Scholar] [CrossRef]
- Oslowski, C.M.; Urano, F. The binary switch that controls the life and death decisions of ER stressed beta cells. Curr. Opin. Cell Biol. 2010, 23, 207–2015. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C.; Ellgaard, L. The human PDI family: Versatility packed into a single fold. Biochim. Biophys. Acta 2008, 1783, 535–548. [Google Scholar] [CrossRef]
- Sramek, J.; Nemcova-Fürstova, V.; Kovar, J. Kinase signaling in apoptosis induced by saturated fatty acids in pancreatic β-cells. Int. J. Mol. Sci. 2016, 17, 1400. [Google Scholar] [CrossRef]
- Martinez, S.C.; Tanabe, K.; Cras-Méneur, C.; Abumrad, N.A.; Bernal-Mizrachi, E.; Permutt, M.A. Inhibition of FoxO1 protects pancreatic islet β-cells against fatty acid and endoplasmic reticulum stress-induced apoptosis. Diabetes 2008, 57, 846–859. [Google Scholar] [CrossRef]
- Macfarlane, W.M.; Cragg, H.; Docherty, H.M.; Read, M.L.; James, R.F.; Aynsley-Green, A.; Docherty, K. Impaired expression of transcription factor IUF1 in a pancreatic beta-cell line derived from a patient with persistent hyperinsulinaemic hypoglycaemia of infancy (nesidioblastosis). FEBS Lett. 1997, 413, 304–308. [Google Scholar] [CrossRef]
- Musilkova, J.; Kovar, J. Additive stimulatory effect of extracellular calcium and potassium on non-transferrin ferric iron uptake by HeLa and K562 cells. Biochim. Biophys. Acta 2001, 1514, 117–126. [Google Scholar] [CrossRef]
- Polak, J.; Studer-Rabeler, K.; McHugh, H.; Hussain, M.A.; Shimoda, L.A. System for exposing cultured cells to intermittent hypoxia utilizing gas permeable cultureware. Gen. Physiol. Biophys. 2015, 34, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Weiszenstein, M.; Pavlikova, N.; Elkalaf, M.; Halada, P.; Seda, O.; Trnka, J.; Kovar, J.; Polak, J. The effect of pericellular oxygen levels on proteomic profile and lipogenesis in 3T3-L1 differentiated preadipocytes cultured on gas-permeable cultureware. PLoS ONE 2016, 11, e0152382. [Google Scholar] [CrossRef] [PubMed]
- Kovar, J.; Franek, F. Growth-stimulating effect of transferrin on a hybridoma cell line relation to transferrin iron-transporting function. Exp. Cell Res. 1989, 182, 358–369. [Google Scholar] [CrossRef]
- Cnop, M.; Hannaert, J.C.; Hoorens, A.; Eizirik, D.L.; Pipeleers, D.G. Inverse relationship between cytotoxicity of free fatty acids in pancreatic islet cells and cellular triglyceride accumulation. Diabetes 2001, 50, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Nemcova-Fürstova, V.; Balusikova, K.; Sramek, J.; James, R.F.; Kovar, J. Caspase-2 and JNK Activated by Saturated Fatty Acids are Not Involved in Apoptosis Induction but Modulate ER Stress in Human Pancreatic β-cells. Cell. Physiol. Biochem. 2013, 31, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Sramek, J.; Nemcova-Fürstova, V.; Balusikova, K.; Daniel, P.; Jelinek, M.; James, R.F.; Kovar, J. P38 MAPK is activated but does not play a key role during apoptosis induction by saturated fatty Acid in human pancreatic β-cells. Int. J. Mol. Sci. 2016, 17, 159. [Google Scholar] [CrossRef] [PubMed]
- Sramek, J.; Nemcova-Fürstova, V.; Pavlikova, N.; Kovar, J. Effect of Saturated Stearic Acid on MAP Kinase and ER Stress Signaling Pathways during Apoptosis Induction in Human Pancreatic β-Cells Is Inhibited by Unsaturated Oleic Acid. Int. J. Mol. Sci. 2017, 18, 2313. [Google Scholar] [CrossRef] [PubMed]
- Lagerstedt, S.A.; Hinrichs, D.R.; Batt, S.M.; Magera, M.J.; Rinaldo, P.; McConnell, J.P. Quantitative determination of plasma c8-c26 total fatty acids for the biochemical diagnosis of nutritional and metabolic disorders. Mol. Genet. Metab. 2001, 73, 38–45. [Google Scholar] [CrossRef]
- Abdelmagid, S.A.; Clarke, S.E.; Nielsen, D.E.; Badawi, A.; El-Sohemy, A.; Mutch, D.M.; Ma, D.W.L. Comprehensive profiling of plasma fatty acid concentrations in young healthy Canadian adults. PLoS ONE 2015, 10, e0116195. [Google Scholar] [CrossRef]
- Carlsson, P.O.; Liss, P.; Andersson, A.; Jansson, L. Measurements of oxygen tension in native and transplanted rat pancreatic islets. Diabetes 1998, 47, 1027–1032. [Google Scholar] [CrossRef]
- Buchwald, P. A local glucose-and oxygen concentration-based insulin secretion model for pancreatic islets. Theor. Biol. Med. Model 2011, 8, 20. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šrámek, J.; Němcová-Fürstová, V.; Polák, J.; Kovář, J. Hypoxia Modulates Effects of Fatty Acids on NES2Y Human Pancreatic β-cells. Int. J. Mol. Sci. 2019, 20, 3441. https://doi.org/10.3390/ijms20143441
Šrámek J, Němcová-Fürstová V, Polák J, Kovář J. Hypoxia Modulates Effects of Fatty Acids on NES2Y Human Pancreatic β-cells. International Journal of Molecular Sciences. 2019; 20(14):3441. https://doi.org/10.3390/ijms20143441
Chicago/Turabian StyleŠrámek, Jan, Vlasta Němcová-Fürstová, Jan Polák, and Jan Kovář. 2019. "Hypoxia Modulates Effects of Fatty Acids on NES2Y Human Pancreatic β-cells" International Journal of Molecular Sciences 20, no. 14: 3441. https://doi.org/10.3390/ijms20143441
APA StyleŠrámek, J., Němcová-Fürstová, V., Polák, J., & Kovář, J. (2019). Hypoxia Modulates Effects of Fatty Acids on NES2Y Human Pancreatic β-cells. International Journal of Molecular Sciences, 20(14), 3441. https://doi.org/10.3390/ijms20143441