Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice

Abstract
1. Introduction
2. Toxic Effects of Cadmium Exposure on Rice
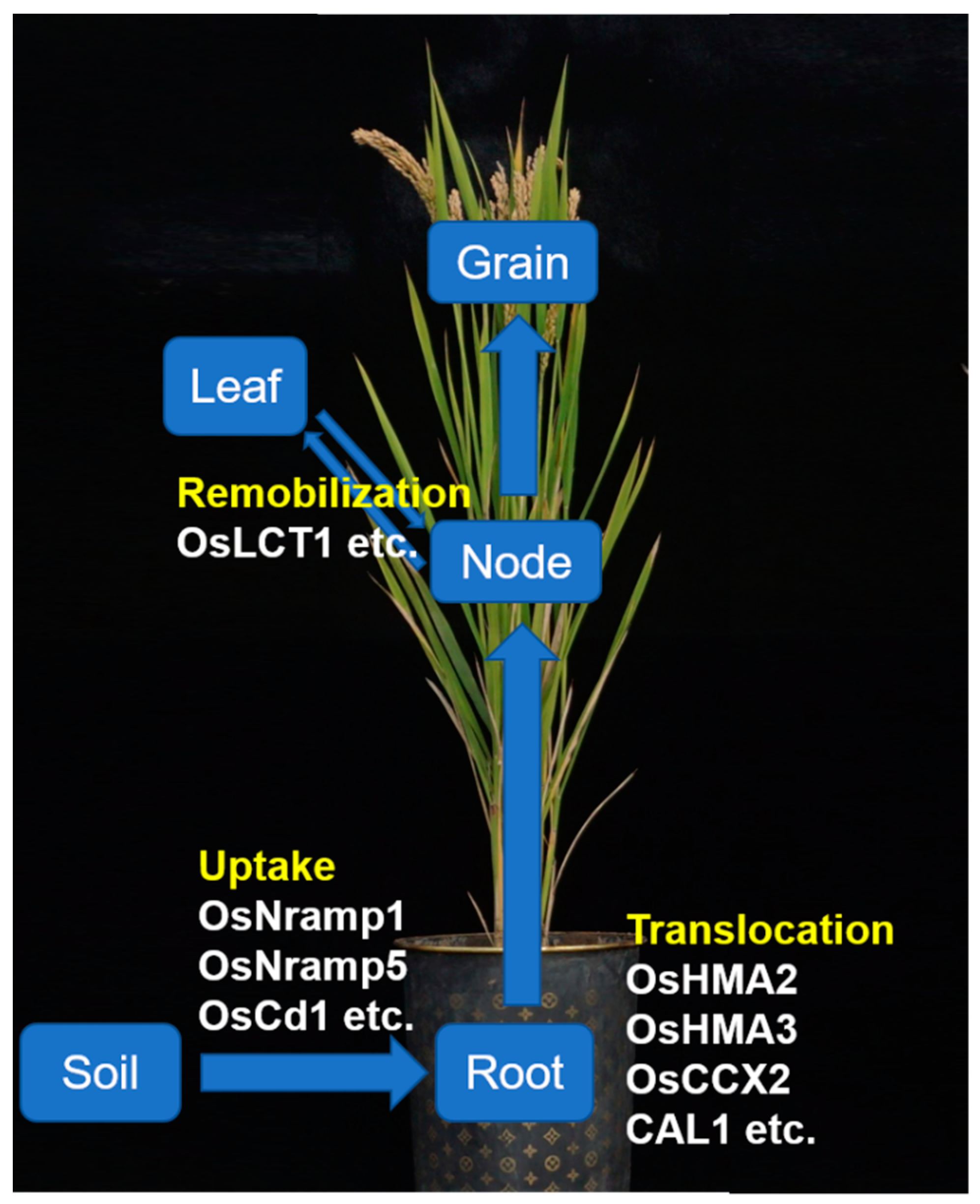
3. Uptake and Transport Pathway of Cd in Rice
3.1. Functional Analysis of Related Genes
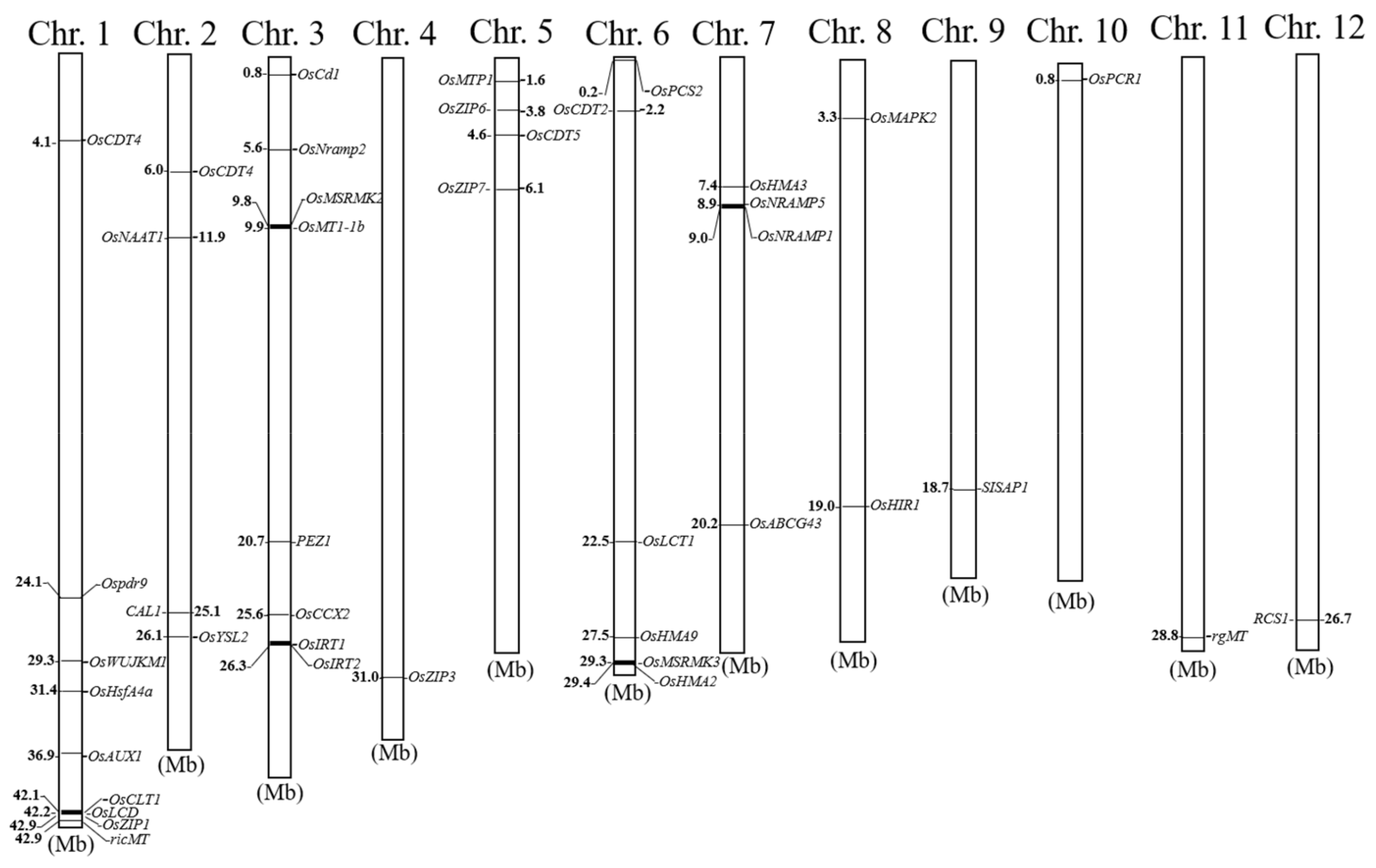
3.2. Location of Related QTLs
4. Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- He, S.Y.; He, Z.L.; Yang, X.E.; Stoffella, P.J.; Baligar, V.C. Soil biogeochemistry, plant physiology, and phytoremediation of cadmium-contaminated soils. Adv. Agron. 2015, 134, 135–225. [Google Scholar]
- Song, W.E.; Chen, S.B.; Liu, J.F.; Chen, L.; Song, N.N.; Li, N.; Liu, B. Variation of Cd concentration in various rice cultivars and derivation of cadmium toxicity thresholds for paddy soil by species-sensitivity distribution. J. Integr. Agric. 2015, 14, 1845–1854. [Google Scholar] [CrossRef]
- Liu, F.; Liu, X.N.; Ding, C.; Wu, L. The dynamic simulation of rice growth parameters under cadmium stress with the assimilation of multi-period spectral indices and crop model. Field Crop. Res. 2015, 183, 225–234. [Google Scholar] [CrossRef]
- Zhao, K.L.; Fu, W.J.; Ye, Z.Q.; Zhang, C.S. Contamination and spatial variation of heavy metals in the soil-rice system in Nanxun County, Southeastern China. Int. J. Environ. Res. Public Health. 2015, 12, 1577–1594. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.P.; Deng, J.W.; Zhang, H.M.; Ma, Y.H.; Cao, D.J.; Ma, R.X.; Liu, R.J.; Liu, C.; Liang, Y.G. Effects of cadmium on bioaccumulation and biochemical stress response in rice (Oryza sativa L.). Ecotox. Environ. Safe. 2015, 122, 392–398. [Google Scholar] [CrossRef]
- Xue, D.W.; Jiang, H.; Deng, X.X.; Zhang, X.Q.; Wang, H.; Xu, X.B.; Hu, J.; Zeng, D.L.; Guo, L.B.; Qian, Q. Comparative proteomic analysis provides new insights into cadmium accumulation in rice grain under cadmium stress. J. Hazard. Mater. 2014, 280, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.; Rafiq, M.T.; Li, T.; Liu, D.; He, Z.; Stoffella, P.J.; Sun, K.; Xiaoe, Y. Uptake of cadmium by rice grown on contaminated soils and its bioavailability/toxicity in human cell lines (Caco-2/HL-7702). J. Agric. Food Chem. 2015, 63, 3599–3608. [Google Scholar] [CrossRef]
- Godt, J.; Scheidig, F.; Grosse-Siestrup, C.; Esche, V.; Brandenburg, P.; Reich, A.; Groneberg, D.A. The toxicity of cadmium and resulting hazards for human health. J. Occup Med. Toxicol. 2006, 1, 22. [Google Scholar] [CrossRef]
- Satarug, S.; Baker, J.R.; Urbenjapol, S.; Haswell-Elkins, M.; Reilly, P.E.; Williams, D.J.; Moore, M.R. A global perspective on cadmium pollution and toxicity in non-occupationally exposed population. Toxicol Lett. 2003, 137, 65–83. [Google Scholar] [CrossRef]
- Horiguchi, H.; Teranishi, H.; Niiya, K.; Aoshima, K.; Katoh, T.; Sakuragawa, N.; Kasuya, M. Hypoproduction of erythropoietin contributes to anemia in chronic cadmium intoxication — clinical-study on itai-itai disease in Japan. Arch. Toxicol. 1994, 68, 632–636. [Google Scholar] [CrossRef]
- Tsukahara, T.; Ezaki, T.; Moriguchi, J.; Furuki, K.; Shimbo, S.; Matsuda-Inoguchi, N.; Ikeda, M. Rice as the most influential source of cadmium intake among general Japanese population. Sci. Total Environ. 2003, 305, 41–51. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Y.; Mao, W.; Sui, H.; Yong, L.; Yang, D.; Jiang, D.; Zhang, L.; Gong, Y. Dietary cadmium exposure assessment among the Chinese population. PLoS ONE 2017, 12, e0177978. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, X.; Wang, P.; Wang, Z.; Li, M.; Zhao, F.J. Dietary cadmium intake from rice and vegetables and potential health risk: A case study in Xiangtan, southern China. Sci. Total Environ. 2018, 639, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Lee, S.H.; Lee, D.G.; Lee, H.; Lee, S.W.; Bahk, J.D.; Lee, B.H. Physiological and protein profiles alternation of germinating rice seedlings exposed to acute cadmiumtoxicity. C. R. Biol. 2007, 330, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.; Qi, X.; Huang, L.; Ye, Z. Identification of rice cultivars with low brown rice mixed cadmium and lead contents and their interactions with the micronutrients iron, zinc, nickel and manganese. J. Environ. Sci. 2012, 24, 1790–1798. [Google Scholar] [CrossRef]
- Li, S.; Yu, J.; Zhu, M.; Zhao, F.; Luan, S. Cadmium impairs ion homeostasis by altering K+ and Ca2+ channel activities in rice root hair cells. Plant Cell Environ. 2012, 35, 1998–2013. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, X.; Li, K.; Wu, M.; Zhang, R.; Zhang, L.; Chen, G. Photosynthetic responses of Oryza sativa L. seedlings to cadmium stress: Physiological, biochemical and ultrastructural analyses. BioMetals 2014, 27, 389–401. [Google Scholar] [CrossRef]
- Kanu, A.S.; Ashraf, U.; Bangura, A.; Yang, D.M.; Ngaujah, A.S.; Tang, X. Cadmium (Cd) Stress in Rice; Phyto-Availability, Toxic Effects, and Mitigation Measures-A Critical Review. IOSR-JESTFT 2017, 11, 07–23. [Google Scholar]
- Liu, J.; Li, K.; Xu, J.; Liang, J.; Lu, X.; Yang, J.; Zhu, Q. Interaction of Cd and five mineral nutrients for uptake and accumulation in different rice cultivars and genotypes. Field Crop. Res. 2003, 83, 271–281. [Google Scholar] [CrossRef]
- Liu, J.G.; Liang, J.S.; Li, K.Q.; Zhang, Z.J.; Yu, B.Y.; Lu, X.L.; Yang, J.C.; Zhu, Q.S. Correlations between cadmium and mineral nutrients in absorption and accumulation in various genotypes of rice under cadmium stress. Chemosphere 2003, 52, 1467–1473. [Google Scholar] [CrossRef]
- Rascio, N.; Dalla Vecchia, F.; La Rocca, N.; Barbato, R.; Pagliano, C.; Raviolo, M.; Gonnelli, C.; Gabbrielli, R. Metal accumulation and damage in rice (cv. Vialone nano) seedlings exposed to cadmium. Environ. Exp. Bot. 2008, 62, 267–278. [Google Scholar] [CrossRef]
- Yu, H.; Wang, J.; Fang, W.; Yuan, J.; Yang, Z. Cadmiumaccumulation in different rice cultivars and screening for pollution safe cultivars of rice. Sci. Total Environ. 2006, 370, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, H.; Zhang, Z.; Wan, X. Influences of different ion channel inhibitors on the absorption of fluoride in tea plants (Camellia sinesis L.). Plant Growth Regul. 2013, 69, 99–106. [Google Scholar]
- Arao, T.; Ae, N. Genotypic variations in cadmium levels of rice grain. Soil Sci. Plant Nutr. 2003, 49, 473–479. [Google Scholar] [CrossRef]
- He, J.; Zhu, C.; Ren, Y.; Yan, Y.; Jiang, D. Genotypic variation in grain cadmium concentration of lowland rice. J. Plant Nutr. Soil Sci. 2006, 169, 711–716. [Google Scholar] [CrossRef]
- Liu, H.J.; Zhang, J.L.; Christie, P.; Zhang, F.S. Influence of external zinc and phosphorus supply on Cd uptake by rice (Oryza sativa L.) seedlings with root surface iron plaque. Plant Soil. 2007, 300, 105–115. [Google Scholar] [CrossRef]
- Rodda, M.S.; Li, G.; Reid, R.J. The timing of grain Cd accumulation in rice plants: The relative importance of remobilisation within the plant and root Cd uptake post flowering. Plant Soil. 2011, 347, 105–114. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, X.; Zeng, M.; Liao, B.H.; Liu, L.; Yang, W.T.; We, Y.M.; Qiu, Q.Y.; Wang, Y.J. Effects of combined amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on contaminated paddy soil. Ecotoxicol. Environ. Saf. 2014, 101, 226–232. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, A.; Ansary, M.M.U.; Watanabe, A.; Fujita, M.; Tran, L.S.P. Hydrogen sulfide modulates cadmium-induced physiological and biochemical responses to alleviate cadmium toxicity in rice. Sci. Rep. 2015, 5, 14078. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Rizwan, M.; Ghafoor, A.; Naeem, A.; Ali, S.; Sabir, M.; Qayyum, M.F. Effect of inorganic amendments for in situ stabilization of cadmium in contaminated soils and its phyto-availability to wheat and rice under rotation. Environ. Sci. Pollut. Res. 2015, 22, 16897–16906. [Google Scholar] [CrossRef]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, X.; Gao, K.; Ge, Y.; Cheng, W. Non-protein thiols and glutathione S-transferase alleviate Cd stress and reduce root-to-shoot translocation of Cd in rice. J. Plant Nutr Soil Sci. 2013, 176, 626–633. [Google Scholar] [CrossRef]
- Choppala, G.; Saifullah, M.F.; Bolan, N.; Bibi, S.; Iqbal, M.; Rengel, Z.; Kunhikrishnan, A.; Ashwath, N.; Ok, Y.S. Cellular mechanisms in higher plants governing tolerance to cadmium toxicity. Crit. Rev. Plant Sci. 2014, 33, 374–391. [Google Scholar] [CrossRef]
- Yu, C.; Sun, C.; Shen, C.; Wang, S.; Liu, F.; Liu, Y.; Chen, Y.; Li, C.; Qian, Q.; Aryal, B.; et al. The auxin transporter, OsAUX1, is involved in primary root and root hair elongation and in Cd stress responses in rice (Oryza sativa L.). Plant J. 2015, 83, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, H.; Chen, L.; Liu, L.; Wan, X. Maintenance of mesophyll potassium and regulation of plasma membrane H+-ATPase are associated with physiological responses of tea plants to drought and subsequent rehydration. Crop. J. 2018, 6, 611–620. [Google Scholar] [CrossRef]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cai, G.; Hausman, J.F. Target or barrier? The cell wall of early- and later-diverging plants vs cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef] [PubMed]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal Interactions between Cadmium-Induced Cell Wall Responses and Oxidative Stress in Plants. Front. Plant Sci. 2017, 8, 1867. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Wang, Y.T.; Yang, Z.Y.; Yuan, J.G. Effects of phosphorus supplied in soil on subcellular distribution and chemical forms of cadmium in two Chinese flowering cabbage (Brassica parachinensis L.) cultivars differing in cadmium accumulation. Food Chem. Toxicol. 2011, 49, 2260–2267. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.O.; Zeng, G.M.; Chai, L.Y.; Song, X.C.; Min, Z.Y.; Xiao, X. Subcellular distribution and chemical forms of cadmium in Bechmeria nivea (L.). Gaud. Environ. Exp. Bot. 2008, 62, 389–395. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, W.; Li, Z.; Liang, Y.; Song, A. Cadmium fate and tolerance in rice cultivars. Agron. Sustain. Dev. 2009, 29, 483–490. [Google Scholar] [CrossRef]
- Fu, X.P.; Dou, C.M.; Chen, Y.X.; Chen, X.C.; Shi, J.Y.; Yu, M.G.; Xu, J. Subcellular distribution and chemical forms of cadmium in Phytolacca americana L. J. Hazard. Mater. 2011, 186, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y.S. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y.S. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Otani, M.; Uraguchi, S.; Akihiro, T.; Fujiwara, T. Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast. Biosci. Biotech. Biochem. 2011, 75, 1211–1213. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Koyama, E.; Yamaji, N.; Ma, J.F. Physiological, genetic, molecular characterization of a high-Cd-accumulating rice cultivar, Jarjan. J. Exp. Bot. 2011, 22, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Mao, W.H.; Zhang, G.P.; Wu, F.B.; Cai, Y. Root excretion and plant tolerance to cadmium toxicity—a review. Plant Soil Environ. 2007, 53, 193–200. [Google Scholar] [CrossRef]
- Haydon, M.J.; Cobbett, C.S. Transporters of ligands for essential metal ions in plants. New Phytol. 2007, 174, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, R.; Ahmad, A.; Iqbal, M. Phytoremediation of heavy metals: Physiological and molecular mechanisms. Bot. Rev. 2009, 75, 339–364. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Nocito, F.F.; Lancilli, C.; Dendena, B.; Lucchini, G.; Sacchi, G.A. Cadmium retention in rice roots is influenced by cadmium availability, chelation and translocation. Plant Cell Environ. 2011, 34, 994–1008. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.J.; Shao, G.; Zhang, G. Influence of cadmium toxicity on growth and antioxidant enzyme activity in rice cultivars with different grain cadmium accumulation. J. Plant Nutr. 2005, 28, 1259–1270. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Kao, C.H. Cadmium-induced oxidative damage in rice leaves is reduced by polyamines. Plant Soil. 2007, 291, 27–37. [Google Scholar] [CrossRef]
- Lin, R.; Wang, X.; Luo, Y.; Du, W.; Guo, H.; Yin, D. Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings Triticum aestivum L. Chemosphere 2007, 69, 89–98. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Antioxidants and stressrelated metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiol. Plant 2012, 34, 835–847. [Google Scholar] [CrossRef]
- Shen, G.M.; Zhu, C.; Du, Q.Z.; Shangguan, L.N. Ascorbate-glutathione cycle alteration in cadmium sensitive rice mutant cadB1. Rice Sci. 2012, 19, 185–192. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Dubey, R.S. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 2014, 251, 1047–1065. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants--role of plant growth regulators. Protoplasma 2015, 252, 399–413. [Google Scholar] [CrossRef]
- Aina, R.; Labra, M.; Fumagalli, P.; Vannini, C.; Marsoni, M.; Cucchi, U.; Bracale, M.; Sgorbati, S.; Citterio, S. Thiol-peptide level and proteomic changes in response to cadmium toxicity in Oryza sativa L. roots. Environ. Exp. Bot. 2007, 59, 381–392. [Google Scholar] [CrossRef]
- Chao, Y.Y.; Chen, C.Y.; Huang, W.D.; Kao, C.H. Salicylic acidmediated hydrogen peroxide accumulation and protection against Cd toxicity in rice leaves. Plant Soil. 2010, 329, 327–337. [Google Scholar] [CrossRef]
- Zhang, C.H.; Ying, G.E. Response of glutathione and glutathione Stransferase in rice seedlings exposed to cadmium stress. Rice Sci. 2008, 15, 73–76. [Google Scholar] [CrossRef]
- Lee, K.; Bae, D.W.; Kim, S.H.; Han, H.J.; Liu, X.; Park, H.C.; Lim, C.O.; Lee, X.Y.; Chung, W.S. Comparative proteomic analysis of the shortterm responses of rice roots and leaves to cadmium. J. Plant Physiol. 2010, 167, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Khavari-Nejad, R.A.; Najafi, F.; Rezaei, M. The influence of cadmium toxicity on some physiological parameters as affected by iron in rice (Oryza Sativa L.) plant. J. Plant Nutr. 2014, 37, 1202–1213. [Google Scholar] [CrossRef]
- Wang, M.Y.; Chen, A.K.; Wong, M.H.; Qiu, R.L.; Cheng, H.; Ye, Z.H. Cadmium accumulation in and tolerance of rice Oryza sativa L. varieties with different rates of radial oxygen loss. Environ. Pollut. 2011, 159, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Rakwal, R.; Yonekura, M.; Kubo, A.; Saji, H. Rapid induction of defense/stress-related proteins in leaves of rice (Oryza sativa) seedlings exposed to ozone is preceded by newly phosphorylated proteins and changes in a 66-kDA ERK-typeMAPK. J. Plant Physiol. 2002, 159, 361–369. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Iwahashi, H. Isolation of novel rice (Oryza sativa L.) multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues. Biochem. Biophys. Res. Commun. 2002, 294, 1009–1016. [Google Scholar] [CrossRef]
- Yeh, C.M.; Hsiao, L.J.H.; Hsiao, H.J. Cadmium activates a mitogenactivated protein kinase gene and MBP kinases in rice. Plant Cell Physiol. 2004, 45, 1306–1312. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Hu, F.; Zhang, S.Y.; Wang, K.; Zhang, C.R.; Liu, T. MAPKs regulate root growth by influencing auxin signaling and cell cyclerelated gene expression in cadmium-stressed rice. Environ. Sci. Pollut. Res. 2013, 20, 5449–5460. [Google Scholar] [CrossRef]
- Wang, X.; Yao, H.; Wong, M.H.; Ye, Z. Dynamic changes in radial oxygen loss and iron plaque formation and their effects on Cd and As accumulation in rice (Oryza sativa L.). Environ. Geochem. Health 2013, 35, 779–788. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, M.; Wong, M.H.; Ye, Z. Does radial oxygen loss and iron plaque formation on roots alter Cd and Pb uptake and distribution in rice plant tissues? Plant Soil. 2014, 375, 137–148. [Google Scholar] [CrossRef]
- Shao, J.F.; Che, J.; Yamaji, N.; Shen, R.F.; Ma, J.F. Silicon reduces cadmium accumulation by suppressing expression of transporter genes involved in cadmium uptake and translocation in rice. J. Exp. Bot. 2017, 68, 5641–5651. [Google Scholar] [CrossRef]
- Chen, Z.; Tang, Y.T.; Yao, A.J.; Cao, J.; Wu, Z.H.; Peng, Z.R.; Wang, S.Z.; Xiao, S.; Baker, A.J.M.; Qiu, R.L. Mitigation of Cd accumulation in paddy rice (Oryza sativa L.) by Fe fertilization. Environ. Pollut. 2017, 231, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Ding, C.; Zhou, Z.; Zhang, T.; Wang, X. A tillering application of zinc fertilizer based on basal stabilization reduces Cd accumulation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2019, 167, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, D.; Xue, R.; Long, J.; Lin, X.; Lin, Y.; Jia, L.; Zeng, R.; Song, Y. Effects of boron, silicon and their interactions on cadmium accumulation and toxicity in rice plants. J. Hazard. Mater. 2019, 367, 447–455. [Google Scholar] [CrossRef]
- Hermans, C.; Conn, S.J.; Chen, J.; Xiao, Q.; Verbruggen, N. An update on magnesium homeostasis mechanisms in plants. Metallomics 2013, 5, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, Y.Y.; Lee, Y.; An, G. Rice P1B-type heavy-metal ATPase, OsHMA9, is a metal efflux protein. Plant Physiol. 2007, 145, 831–842. [Google Scholar] [CrossRef]
- Shimo, H.; Ishimaru, Y.; An, G.; Yamakawa, T.; Nakanishi, H.; Nishizawa, N.K. Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice. J. Exp Bot. 2011, 62, 5727–5734. [Google Scholar] [CrossRef]
- Kuramata, M.; Masuya, S.; Takahashi, Y.; Kitagawa, E.; Inoue, C.; Ishikawa, S.; Youssefian, S.; Kusano, T. Novel cysteine-rich peptides from Digitaria ciliaris and Oryza sativa enhance tolerance to cadmium by limiting its cellular accumulation. Plant Cell Physiol. 2009, 50, 106–117. [Google Scholar] [CrossRef]
- Yang, J.; Gao, M.X.; Hu, H.; Ding, X.M.; Lin, H.W.; Wang, L.; Xu, J.M.; Mao, C.Z.; Zhao, F.J.; Wu, Z.C. OsCLT1, a CRT-like transporter 1, is required for glutathione homeostasis and arsenic tolerance in rice. New Phytol. 2016, 211, 658–670. [Google Scholar] [CrossRef]
- Moons, A. Ospdr9, which encodes a PDR-type ABC transporter, is induced by heavy metals, hypoxic stress and redox perturbations in rice roots. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Agrawal, S.K.; Shibato, J.; Iwahashi, H.; Rakwal, R. Novel rice MAP kinases OsMSRMK3 and OsWJUMK1 involved in encountering diverse environmental stresses and developmental regulation. Biochem. Biophys. Res. Commun. 2003, 300, 775–783. [Google Scholar] [CrossRef]
- Shim, D.; Jae-Ung, H.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell. 2009, 21, 4031–4043. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.A.; Shin, R.; Eide, D.J.; Schachtman, D.P. Differential metal selectivity and gene expression of two zinc transporters from rice. Plant Physiol. 2003, 133, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.S.; Chao, Y.Y.; Huang, W.D.; Hong, C.Y.; Kao, C.H. Effect of magnesium deficiency on antioxidant status and cadmium toxicity in rice seedlings. J. Plant Physiol. 2011, 168, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.H.; Umeda, M.; Liu, J.Y.; Zhao, N.M.; Uchimiya, H. A novel MT gene of rice plants is strongly expressed in the node portion of the stem. Gene 1998, 206, 29–35. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, F.; Shou, H.; Huang, F.; Zheng, L.; He, F.; Li, J.; Zhao, F.J.; Ueno, D.; Ma, J.F.; et al. Mutation in nicotianamine aminotransferase stimulated the Fe(II) acquisition system and led to iron accumulation in rice. Plant Physiol. 2007, 145, 1647–1657. [Google Scholar] [CrossRef]
- Luo, J.S.; Huang, J.; Zeng, D.L.; Peng, J.S.; Zhang, G.B.; Ma, H.L.; Guan, Y.; Yi, H.Y.; Fu, Y.L.; Han, B.; et al. A defensin-like protein drives cadmium efflux and allocation in rice. Nat. Commun. 2018, 9, 645. [Google Scholar] [CrossRef]
- Masuda, H.; Ishimaru, Y.; Aung, M.S.; Kobayashi, T.; Kakei, Y.; Takahashi, M.; Higuchi, K.; Nakanishi, H.; Nishizawa, N.K. Iron biofortification in rice by the introduction of multiple genes involved in iron nutrition. Sci. Rep. 2012, 2, 543. [Google Scholar] [CrossRef]
- Yan, H.; Xu, W.; Xie, J.; Gao, Y.; Wu, L.; Sun, L.; Feng, L.; Chen, X.; Zhang, T.; Dai, C.; et al. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Ansarypour, Z.; Shahpiri, A. Heterologous expression of a rice metallothionein isoform (OsMTI-1b) in Saccharomyces cerevisiae enhances cadmium, hydrogen peroxide and ethanol tolerance. Braz. J. Microbiol. 2017, 48, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Kakei, Y.; Shimo, H.; Bashir, K.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. A rice phenolic efflux transporter is essential for solubilizing precipitated apoplasmic iron in the plant stele. J. Biol. Chem. 2011, 286, 24649–24655. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Zeng, M.; Wang, J.; Zeng, Z.; Dai, J.; Xie, Z.; Yang, Y.; Tian, L.; Chen, L.; Li, D. A Node-Expressed Transporter OsCCX2 Is Involved in Grain Cadmium Accumulation of Rice. Front. Plant Sci. 2018, 9, 476. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, S.; Liu, B.; Zhang, M.; Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 2012, 31, 67–79. [Google Scholar] [CrossRef]
- Kavitha, P.G.; Kuruvilla, S.; Mathew, M.K. Functional characterization of a transition metal ion transporter, OsZIP6 from rice (Oryza sativa L.). Plant Physiol. Biochem. 2015, 97, 165–174. [Google Scholar]
- Tan, L.; Zhu, Y.; Fan, T.; Peng, C.; Wang, J.; Sun, L.; Chen, C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem. Biophys. Res. Commun. 2019, 512, 112–118. [Google Scholar] [CrossRef]
- Das, N.; Bhattacharya, S.; Bhattacharyya, S.; Maiti, M. Identification of alternatively spliced transcripts of rice phytochelatin synthase 2 gene OsPCS2 involved in mitigation of cadmium and arsenic stresses. Plant Mol. Biol. 2017, 94, 167–183. [Google Scholar] [CrossRef]
- Uraguchi, S.; Kamiya, T.; Sakamoto, T.; Kasai, K.; Sato, Y.; Nagamura, Y.; Yoshida, A.; Kyozuka, J.; Ishikawa, S.; Fujiwara, T. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc. Natl. Acad Sci. 2011, 108, 20959–20964. [Google Scholar] [CrossRef] [PubMed]
- Uraguchi, S.; Kamiya, T.; Clemens, S.; Fujiwara, T. Characterization of OsLCT1, a cadmium transporter from indica rice Oryza sativa. Physiol. Plant 2014, 151, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell. 2012, 24, 2155–2167. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.Y.; Zhang, L.; Hu, J.; Zhang, X.; Lu, K.; Dong, H.; Wang, D.; Zhao, F.J.; Huang, C.F.; et al. OsNRAMP5 contributes to manganese translocation and distribution in rice shoots. J. Exp. Bot. 2014, 65, 4849–4861. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S.; et al. From laboratory to field: OsNRAMP5-knockdown rice is a promising candidate for Cd phytoremediation in paddy fields. PLoS ONE 2014, 9, e98816. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal. Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.D.; Hwang, J.G.; Han, A.R.; Park, Y.C.; Lee, C.; Ok, Y.S.; Jang, C.S. Positive regulation of rice RING E3 ligase OsHIR1 in arsenic and cadmium uptakes. Plant Mol. Biol. 2014, 85, 365–379. [Google Scholar] [CrossRef] [PubMed]
- 125Mukhopadhyay, A.; Vij, S.; Tyagi, A.K. Overexpression of a zinc-finger protein gene from rice confers tolerance to cold, dehydration, and salt stress in transgenic tobacco. Proc. Natl. Acad. Sci. USA 2004, 101, 6309–6314. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.J.; Wang, M.; Liu, Z.P.; Shi, Y.; Han, T.Q.; Ye, Y.Y.; Gong, N.; Sun, J.W.; Zhu, C. Different responses of low grain-Cd-accumulating and high grain-Cd-accumulating rice cultivars to Cd stress. Plant Mol. Biol. 2015, 96, 261–269. [Google Scholar]
- Jin, S.; Cheng, Y.; Guan, Q.; Liu, D.; Takano, T.; Liu, S. A metallothionein-like protein of rice (rgMT) functions in E. coli and its gene expression is induced by abiotic stresses. Biotechnol. Lett. 2006, 28, 1749–1753. [Google Scholar]
- Harada, E.; Choi, Y.E.; Tsuchisaka, A.; Obata, H.; Sano, H. Transgenic tobacco plants expressing a rice cysteine synthase gene are tolerant to toxic levels of cadmium. J. Plant Physiol. 2001, 158, 655–661. [Google Scholar] [CrossRef]
- Lu, L.L.; Tian, S.K.; Yang, X.E.; Li, T.Q.; He, Z.L. Cadmium uptake and xylem loading are active processes in the hyperaccumulator Sedum alfredii. J. Plant Physiol. 2009, 166, 579–587. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Suzuki, M.; Tsukamoto, T.; Suzuki, K.; Nakazono, M.; Kobayashi, T.; Wada, Y.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J. 2006, 45, 335–346. [Google Scholar] [CrossRef]
- Liu, C.; Chen, G.; Li, Y.; Peng, Y.; Zhang, A.; Hong, K.; Jiang, H.; Ruan, B.; Zhang, B.; Yang, S.; et al. Characterization of a major QTL for manganese accumulation in rice grain. Sci. Rep. 2017, 7, 17704. [Google Scholar] [CrossRef] [PubMed]
- Uraguchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: Perspectives for reducing grain cadmium accumulation. Rice 2012, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Nagasawa, N.; Mori, M.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Functional relationship heavy metal P-type ATPases (OsHMA2 and OsHMA3) of rice (Oryza sativa) using RNAi. Plant Biotechnol. 2013, 30, 511–515. [Google Scholar] [CrossRef]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Fujimaki, S.; Fujiwara, T.; Yoneyama, T.; Hayashi, H. Quantitative estimation of the contribution of the phloem in cadmium transport to grains in rice plants (Oryza sativa L.). Soil Sci. Plant Nutr. 2007, 53, 72–77. [Google Scholar] [CrossRef]
- Wu, Z.C.; Zhao, X.H.; Sun, X.C.; Tan, Q.L.; Tang, Y.F.; Nie, Z.J.; Hu, C.X. Xylem transport and gene expression play decisive roles in cadmium accumulation in shoots of two oilseed rape cultivars (Brassica napus). Chemosphere 2015, 119, 1217–1223. [Google Scholar] [CrossRef]
- Fujimaki, S.; Suzui, N.; Ishioka, N.S.; Kawachi, N.; Ito, S.; Chino, M.; Nakamura, S. Tracing cadmium from culture to spikelet: Noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant. Plant Physiol. 2010, 152, 1796–1806. [Google Scholar] [CrossRef]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef]
- Ishikawa, S.; Abe, T.; Kuramata, M.; Yamaguchi, M.; Ando, T.; Yamamoto, T.; Yano, M. A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7. J. Exp. Bot. 2010, 61, 923–934. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Shindoh, K.; Hirotsu, N.; Ishimaru, K. Evidence for separate translocation pathways in determining cadmium accumulation in grain and aerial plant parts in rice. BMC Plant Biol. 2009, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Shirasawa, S.; Maeda, H.; Nakagomi, K.; Kaji, R.; Ohta, H.; Yamaguchi, M.; Takeshi, N. Analysis of QTL for lowering cadmium concentration in rice grains from ‘LAC- 23′. Breed. Sci. 2011, 61, 196–200. [Google Scholar]
- Yan, Y.F.; Lestari, P.; Lee, K.J.; Kim, M.Y.; Lee, S.H.; Lee, B.W. Identification of quantitative trait loci for cadmi um accumulation and distribution in rice (Oryza sativa). Genome 2013, 56, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Nonoue, Y.; Ono, N.; Omoteno, M.; Kuramata, M.; Fukuoka, S.; Yamamoto, T.; Yano, M.; Ishikawa, S. Detection of QTLs to reduce cadmium content in rice grains using LAC23/Koshihikari chromosome segment substitution lines. Breed. Sci. 2013, 63, 284–291. [Google Scholar]
- Liu, W.; Pan, X.; Li, Y.; Duan, Y.; Min, J.; Liu, S.; Liu, L.; Sheng, X.; Li, X. Identification of QTLs and Validation of qCd-2 Associated with Grain Cadmium Concentrations in Rice. Rice Sci. 2019, 26, 42–49. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Chen, M.; Zheng, G.; Peng, Y.; Shi, X.; Qin, P.; Xu, X.; Teng, S. Association Study Reveals Genetic Loci Responsible for Arsenic, Cadmium and Lead Accumulation in Rice Grain in Contaminated Farmlands. Front. Plant Sci. 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Koyama, E.; Kono, I.; Ando, T.; Yano, M.; Ma, J.F. Identification of a novel major quantitative trait locus controlling distribution of Cd between roots and shoots in rice. Plant Cell Physiol. 2009, 50, 2223–2233. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Kono, I.; Yokosho, K.; Ando, T.; Yano, M.; Ma, J.F. A major quantitative trait locus controlling cadmium translocation in rice (Oryza sativa). New Phytol. 2009, 182, 644–653. [Google Scholar] [CrossRef]
- Xue, D.; Chen, M.; Zhang, G. Mapping of QTLs associated with cadmium tolerance and accumulation during seedling stage in rice (Oryza sativa L.). Euphytica 2009, 165, 587. [Google Scholar] [CrossRef]
- Norton, G.J.; Deacon, C.M.; Xiong, L.Z.; Huang, S.Y.; Meharg, A.A.; Price, A.H. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 2010, 329, 139–153. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ae, N.; Yano, M. Chromosomal regions with quantitative trait loci controlling cadmium concentration in brown rice (Oryza sativa). New Phytol. 2005, 168, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Taguchi-Shiobara, F.; Kojima, Y.; Ebitani, T.; Kuramata, M.; Yamamoto, T.; Yano, M.; Ishikawa, S. Detection of a QTL for accumulating Cd in rice that enables efficient Cd phytoextraction from soil. Breed. Sci. 2011, 61, 43–51. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Zhang, G.P.; Guo, L.B.; Wang, H.Z.; Zeng, D.L.; Dong, G.J.; Qian, Q.; Xue, D.W. Identification of quantitative trait loci for Cd and Zn concentrations of brown rice grown in Cd-polluted soils. Euphytica 2011, 180, 173–179. [Google Scholar] [CrossRef]
- Huang, Y.; Sun, C.; Min, J.; Chen, Y.; Tong, C.; Bao, J. Association Mapping of Quantitative Trait Loci for Mineral Element Contents in Whole Grain Rice (Oryza sativa L.). J. Agric. Food Chem. 2015, 63, 10885–10892. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Zhou, Q.; Ma, L.Q. Availability and assessment of fixing additives for the in-situ remediation of heavy metal contaminated soils: A review. Environ. Monit. Assess. 2006, 116, 513–528. [Google Scholar] [CrossRef] [PubMed]
- Madejón, E.; de Mora, A.P.; Felipe, E.; Burgos, P.; Cabrera, F. Soil amendments reduce trace element solubility in a contaminated soil and allow regrowth of natural vegetation. Environ. Pollut. 2006, 139, 40–52. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, Y.Q.; Liang, C.H. Evaluation of phosphate fertilizers for the immobilization of Cd in contaminated soils. PLoS ONE 2015, 10, e0124022. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hu, P.J.; Ouyang, Y.N.; Wu, L.H.; Shen, L.B.; Luo, Y.M.; Christie, P. Effects of water management on arsenic and cadmium speciation and accumulation in an upland rice cultivar. J. Environ. Sci. 2015, 27, 225–231. [Google Scholar] [CrossRef]
- Honma, T.; Ohba, H.; Kaneko-Kadokura, A.; Makino, T.; Nakamura, K.; Katou, H. Optimal soil Eh, pH, and water management for simultaneously minimizing arsenic and cadmium concentrations in rice grains. Environ. Sci. Technol. 2016, 50, 4178–4185. [Google Scholar] [CrossRef]
- Yu, L.L.; Zhu, J.Y.; Huang, Q.Q.; Su, D.C.; Jiang, R.F.; Li, H.F. Application of a rotation system to oilseed rape and rice fields in Cd-contaminated agricultural land to ensure food safety. Ecotox. Environ. Safe. 2014, 108, 287–293. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ishimaru, Y.; Igura, M.; Kuramata, M.; Abe, T.; Senoura, T.; Hase, Y.; Arao, T.; Nishizawa, N.K.; Nakanishi, H. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. USA 2012, 109, 19166–19171. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, L.; Tang, Z.; Huang, X.Y.; Ma, J.F.; Zhao, F.J. Producing cadmium-free Indica rice by overexpressing OsHMA3. Environ. Int. 2019, 126, 619–626. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Chr. | Physical Location (bp) | Gene Name | Function | Reference |
|---|---|---|---|---|---|
| OsCDT3 | 1 | 4066623–4067218 | Encoding a Cys-rich peptide | Cd uptake inhibitor | [79] |
| Ospdr9 | 1 | 24075065–24082181 | Multidrug resistance ABC transporter | Redox protection in Cd stress | [81] |
| OsWJUMK1 | 1 | 29398191–29402466 | Mitogen-activated protein kinase | Cd signal | [82] |
| OsHsfA4a | 1 | 31370413–31372729 | Heat shock transcription factor gene | Cd tolerance | [83] |
| OsAUX1 | 1 | 36998334–37004685 | Auxin transport protein | Root development and Cd stress response | [34] |
| OsCLT1 | 1 | 42086484–42095424 | CRT-like transporter 1 | Cd tolerance | [80] |
| OsLCD | 1 | 42162592–42166462 | Low cadmium | Cd tolerance and accumulation | [78] |
| OsZIP1 | 1 | 42905566–42907474 | Zinc- and iron-regulated transporter | Cd and Zn transport | [84,85] |
| ricMT | 1 | 43047164–43047861 | Metallothionein gene | Cd tolerance | [86] |
| OsCDT4 | 2 | 6078179–6079111 | Encoding a Cys-rich peptide | Cd uptake inhibitor | [79] |
| OsNAAT1 | 2 | 11997094–12002633 | Nicotinamide aminotransferase gene | Cd accumulation | [87] |
| CAL1 | 2 | 25190487–25191188 | defensin-like protein | Cd accumulation in leaf | [88] |
| OsYSL2 | 2 | 26170387–26174970 | Metal-nicotinamide transporter | Cd translocation | [89] |
| OsCd1 | 3 | 842577–846408 | Major facilitator superfamily | Cd uptake | [90] |
| OsNramp2 | 3 | 5655157–5659147 | Natural resistance-associated macrophage protein | Cd transporter, Cd accumulation | [91] |
| OsMSRMK2 | 3 | 9847700–9850473 | Mitogen-activated protein kinase | Cd signal | [67] |
| OsMTI-1b | 3 | 9957335–9958362 | Metallothionein-like protein 1B | Cd tolerance | [92] |
| PEZ1 | 3 | 20793053–20799805 | Phenol efflux protein | Cd accumulation | [93] |
| OsCDT1/OsCCX2 | 3 | 25613825–25616179 | Cation/calcium (Ca) exchanger 2 | Cd tolerance and translocation | [79,94] |
| OsIRT2 | 3 | 26276301–26277206 | Iron-regulated transporter | Cd and Fe transporter | [95] |
| OsIRT1 | 3 | 26286156–26292023 | Iron-regulated transporter | Cd and Fe transporter | [95,96] |
| OsZIP3 | 4 | 31078200–31080734 | Zinc- and iron-regulated transporter | Cd accumulation | [84] |
| OsMTP1 | 5 | 1675488–1679056 | Metal tolerance protein gene | Cd translocation | [97] |
| OsZIP6 | 5 | 3807974–3810752 | Zinc- and iron-regulated transporter | Cd transport | [98] |
| OsCDT5 | 5 | 4665325–4667853 | Encoding a Cys-rich peptide | Cd uptake inhibitor | [79] |
| OsZIP7 | 5 | 6090801–6094068 | Zinc- and iron-regulated transporter | Cd and Zn accumulation | [99] |
| OsPCS2 | 6 | 167367–174319 | Plant chelatase synthase 2 | Cd tolerance | [100] |
| OsCDT2 | 6 | 2261681–2263972 | Encoding a Cys-rich peptide | Cd uptake inhibitor | [79] |
| OsLCT1 | 6 | 22566775–22571982 | Low affinity cation transporter | Cd transporter in phloem | [101,102] |
| OsHMA9 | 6 | 27517100–27523604 | P-Type Heavy Metal ATPase | Cd efflux | [77] |
| OsMSRMK3 | 6 | 29398191–29402466 | Mitogen-activated protein kinase | Cd signal | [82] |
| OsHMA2 | 6 | 29477949–29480905 | P-Type Heavy Metal ATPase | Cd and Zn translocation | [103,104] |
| OsHMA3 | 7 | 7405745–7409553 | P-Type Heavy Metal ATPase | Sequestration of Cd in root | [46,47,105] |
| OsNramp5 | 7 | 8871436–8878905 | Natural resistance-associated macrophage protein | Cd, Mn, and Fe transporters | [106,107,108,109,110] |
| OsNramp1 | 7 | 8966025–8970882 | Natural resistance-associated macrophage protein | Cd and Fe transporters | [111,112,113] |
| OsABCG43 | 7 | 20214025–20218702 | ATP-binding cassette transporter | Cd compartmentalization | [45] |
| OsMAPK2 | 8 | 3307520–3310590 | Mitogen-activated protein kinase | Cd signal | [68] |
| OsHIR1 | 8 | 19011814–19015998 | Heavy metal-induced RING E3 ligase 1 | Cd uptake | [114] |
| SISAP1 | 9 | 18760704–18761836 | Subspecies indica stress-associated protein gene | Cd tolerance | [115] |
| OsPCR1 | 10 | 826309–824623 | Plant cadmium resistance 1 | Cd tolerance | [116] |
| rgMT | 11 | 28827746–28828439 | Metallothionein-like protein | Cd tolerance | [117] |
| RCS1 | 12 | 26698650–26703087 | Cytosolic cysteine synthase gene | Cd complexation via sulfur | [118] |
| Stage | Parent Sources | Population | Marker | Trait | Chr. | QTL | Reference |
|---|---|---|---|---|---|---|---|
| Seedling stage | Tainan1/Chunjiang06 | 119 DH, 3651 BC3F3 | RFLP | Cd accumulation in leaves | 2 | CAL1 | [88] |
| Seedling stage | Nipponbare/Anjana Dhan | 965 F2 | SSR | Cd concentration in shoots | 7 | OsHMA3 | [105] |
| Seedling stage | SNU-SG1/Suwon490 | 91 RIL | 124 SSR | Cd concentration in shoots | 10 | scc10 | [133] |
| Seedling stage | Koshihikari/LAC23 | 46 CSSLs | 345 SNP | Cd concentration in shoots | 3 | glGCd3 | [134] |
| Seedling stage | Anjana Dhan/Nipponbare | 177 F2 | SSR | Root-to-shoot Cd translocation | 7 | qCdT7 | [137] |
| Seedling stage | Badari Dhan/Shwe War | 184 F2 | 141 SSR | Cd concentration in shoots | 2,5,11 | — | [138] |
| Seedling stage | JX17/ZYQ8 | 127 DH | 160 RFLP,83 SSR | Shoot/root rate of Cd concentration | 3 | qSRR3 | [139] |
| Seedling stage | JX17/ZYQ8 | 127 DH | 160 RFLP,83 SSR | Cd concentration in roots and shoots | 6,7 | qCDS7, qCDR6.1, qCDR6.2 | [139] |
| Seedling stage | Azucena/Bala | 79 RIL | 164 SSR | Cd concentration in leaves | 1,3,6 | qCd1, qCd3, qCd6, | [140] |
| Bfore heading | Kasalath/Nipponbare | 98 BILs | RFLP and SSR | Cd concentration in leaves and culms | 4,11 | qcd4–1, qcd4–2, qcd11 | [131] |
| Mature period | Sasanishiki/Habataki | 85 BIL | SSR | Cd accumulation in grains | 2,7 | qGCd7 | [130] |
| Mature period | Fukuhibiki/LAC23 | 126 RIL | 454 SNP | Cd accumulation in grains | 3,11 | gLCdG3, gLCdG11 | [132] |
| Mature period | SNU-SG1/Suwon490 | 91 RIL | 124 SSR | Cd accumulation in grains | 3,5,9,11 | gcc3, sgr5, gcc9, gcc11 | [133] |
| Mature period | Xiang 743/Katy | 115 RIL, | SSR | Cd accumulation in grains | 2,7 | qCd-2, qCd-7 | [135] |
| Mature period | Kasalath/Koshihikari | 39CSSL | 129 RFLP | Cd accumulation in grains | 3,6,8 | — | [141] |
| Mature period | Koshihikari/Jarjan | 103 BIL | 169 SSR | Cd accumulation in grains | 7 | — | [142] |
| Mature period | JX17/ZYQ8 | 127 DH | 160 RFLP,83 SSR | Cd accumulation in grains | 3,6 | gCdc3, gCdc6 | [143] |
| Mature period | 127 rice cultivars | GWAS | Cd accumulation in grains | 3 | OsCd1 | [90] | |
| Mature period | 378 rice cultivars | GWAS | Cd accumulation in grains | 3, 5 | qCd3, qCd5.1, qCd5.2 | [144] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zou, W.; Meng, L.; Fan, X.; Xu, G.; Ye, G. Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice. Int. J. Mol. Sci. 2019, 20, 3417. https://doi.org/10.3390/ijms20143417
Chen J, Zou W, Meng L, Fan X, Xu G, Ye G. Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice. International Journal of Molecular Sciences. 2019; 20(14):3417. https://doi.org/10.3390/ijms20143417
Chicago/Turabian StyleChen, Jingguang, Wenli Zou, Lijun Meng, Xiaorong Fan, Guohua Xu, and Guoyou Ye. 2019. "Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice" International Journal of Molecular Sciences 20, no. 14: 3417. https://doi.org/10.3390/ijms20143417
APA StyleChen, J., Zou, W., Meng, L., Fan, X., Xu, G., & Ye, G. (2019). Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice. International Journal of Molecular Sciences, 20(14), 3417. https://doi.org/10.3390/ijms20143417