In Vitro Entero-Capillary Barrier Exhibits Altered Inflammatory and Exosomal Communication Pattern after Exposure to Silica Nanoparticles

 and
and 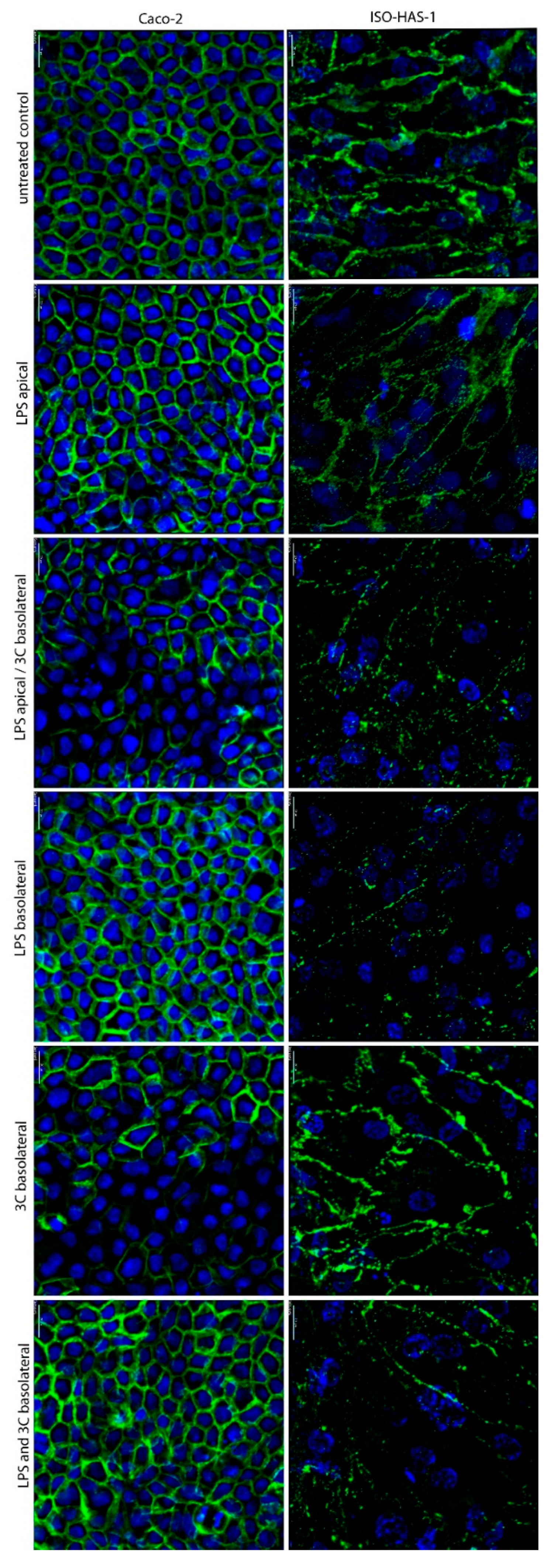{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
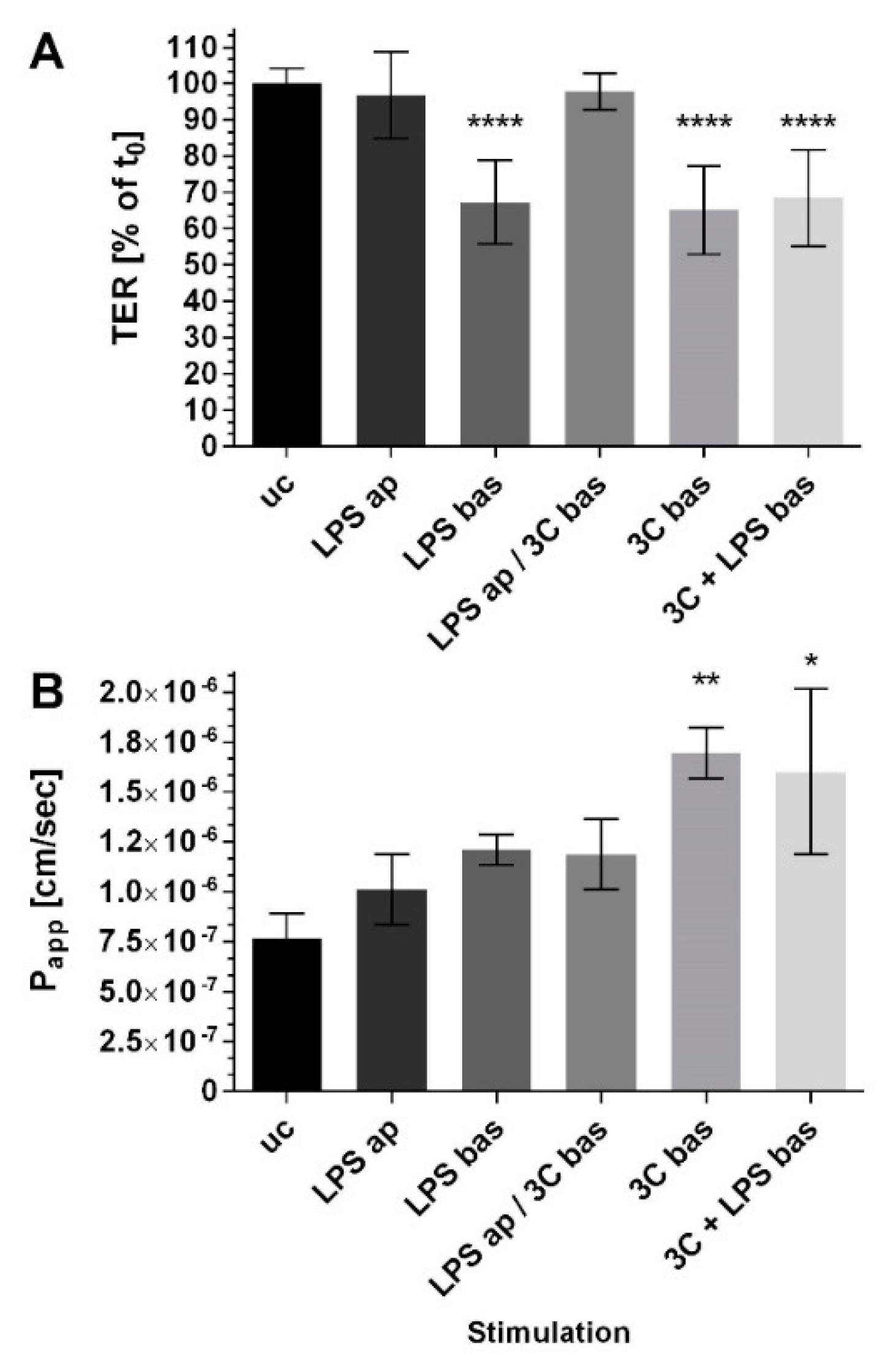
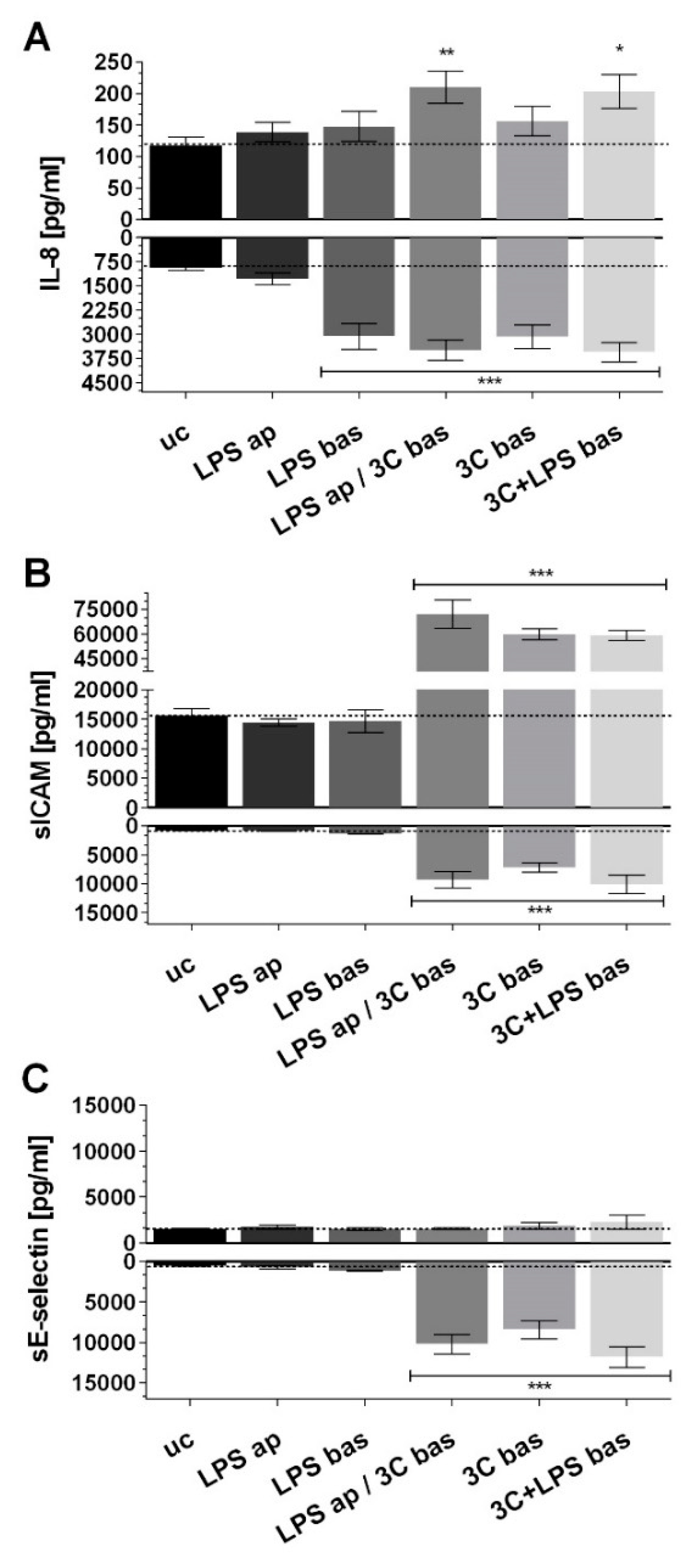
2. Results
3. Discussion
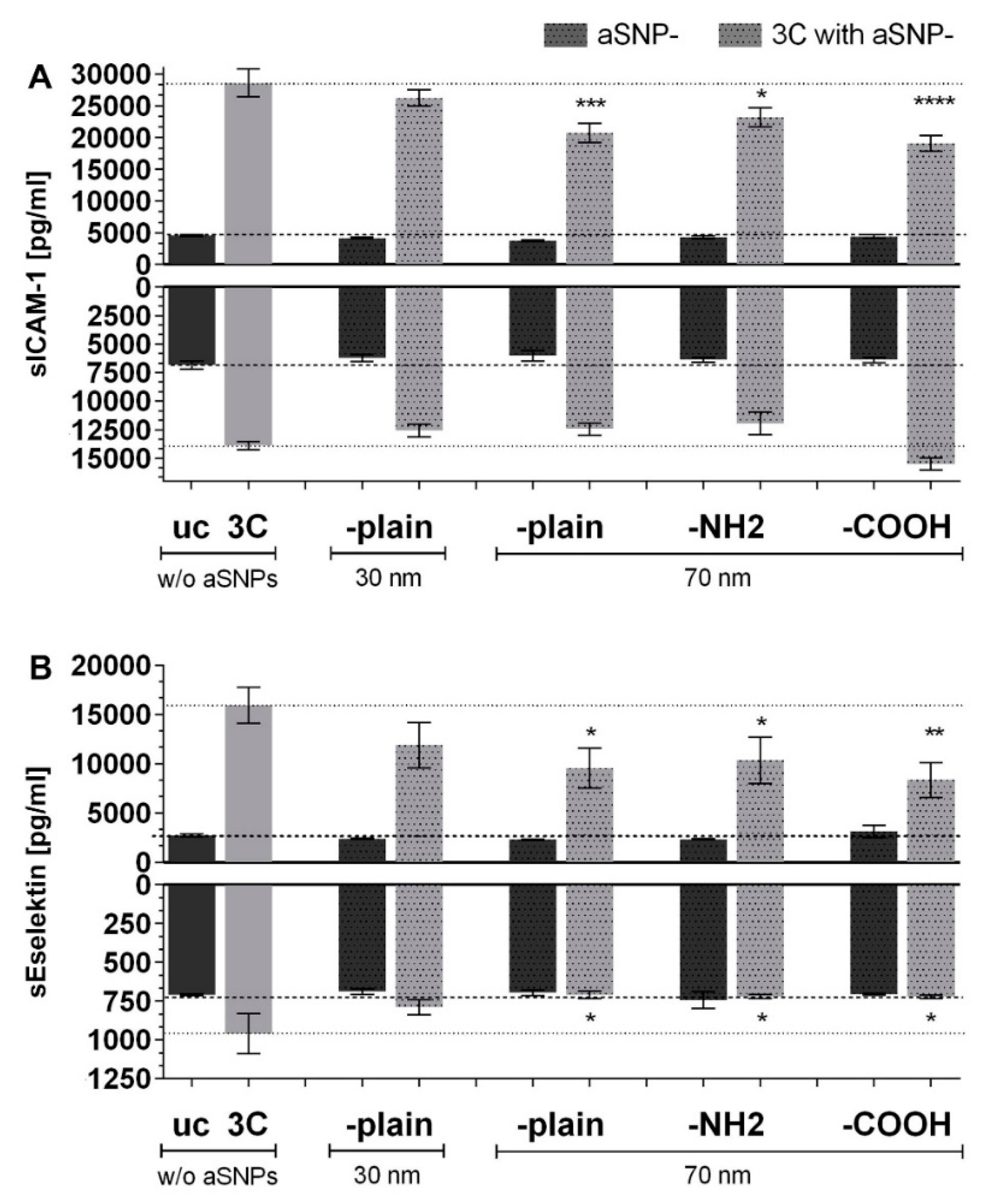
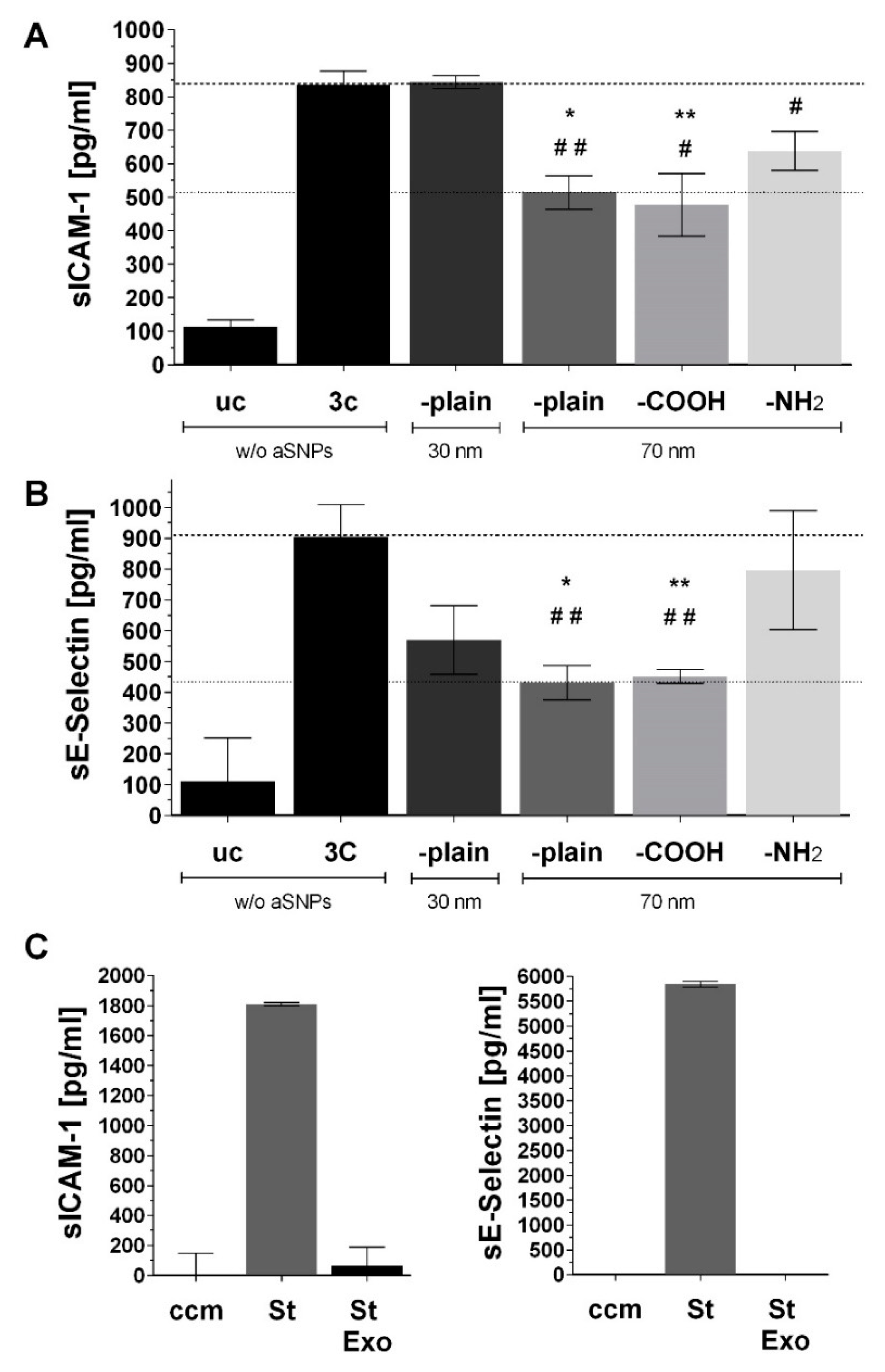
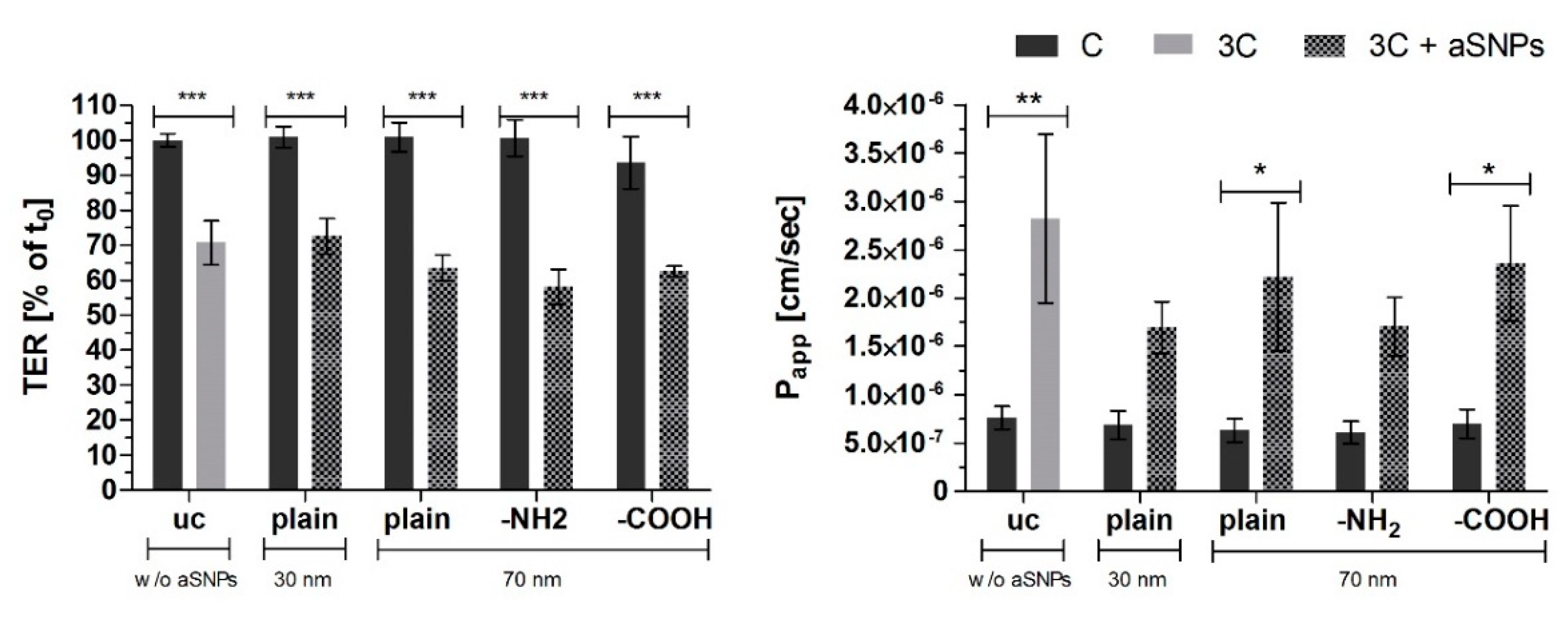
3.1. aSNP Exposure to the Endothelium Decreases sICAM and sE-Selectin Levels on both Sides of the Inflamed Coculture Model
3.2. The Endothelial Release of ICAM and E-Selectin-Bearing Exosomes/Microvesicles Is Increased during Endothelial Inflammation and Reduced during Endothelial aSNP Treatment
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Ethical Standards
Abbreviations
| iMV | intestinal microvasculature |
| IBD | inflammatory bowel disease |
| ICAM-1 | intercellular adhesion molecule 1 |
| TER | transepithelial-electrical resistance |
| Papp | permeability-coefficient |
| aSNP | amorphous silica nanoparticles |
| ELISA | enzyme-linked Immunosorbent Assay |
| EMP | endothelial microparticles |
| IMEC | intestinal microvascular endothelial cells |
| NaFlu | Sodium-Fluorescein |
| CC | coculture Caco-2/ISO-HAS-1 |
| CCinv | inverted coculture ISO-HAS-1/Caco-2 |
| 3C | cytokine mixture ((IL-1β, TNF-α, IFN-γ) |
| ap | apical stimulation |
| bas | Basolateral stimulation |
| ccm | cell culture media |
References
- Lissner, D.; Schumann, M.; Batra, A.; Kredel, L.I.; Kuhl, A.A.; Erben, U.; May, C.; Schulzke, J.D.; Siegmund, B. Monocyte and m1 macrophage-induced barrier defect contributes to chronic intestinal inflammation in ibd. Inflamm. Bowel Dis. 2015, 21, 1297–1305. [Google Scholar] [CrossRef]
- Detzel, C.J.; Horgan, A.; Henderson, A.L.; Petschow, B.W.; Warner, C.D.; Maas, K.J.; Weaver, E.M. Bovine immunoglobulin/protein isolate binds pro-inflammatory bacterial compounds and prevents immune activation in an intestinal co-culture model. PLoS ONE 2015, 10, e0120278. [Google Scholar] [CrossRef]
- Chalubinski, M.; Wojdan, K.; Gorzelak, P.; Borowiec, M.; Broncel, M. The effect of oxidized cholesterol on barrier functions and il-10 mrna expression in human intestinal epithelium co-cultured with dendritic cells in the transwell system. Food Chem. Toxicol. 2014, 69, 289–293. [Google Scholar] [CrossRef]
- Leonard, F.; Collnot, E.M.; Lehr, C.M. A three-dimensional coculture of enterocytes, monocytes and dendritic cells to model inflamed intestinal mucosa in vitro. Mol. Pharm. 2010, 7, 2103–2119. [Google Scholar] [CrossRef]
- Rogoz, A.; Reis, B.S.; Karssemeijer, R.A.; Mucida, D. A 3-D enteroid-based model to study t-cell and epithelial cell interaction. J. Immunol. Methods 2015, 421, 89–95. [Google Scholar] [CrossRef]
- Matsusaki, M.; Hikimoto, D.; Nishiguchi, A.; Kadowaki, K.; Ohura, K.; Imai, T.; Akashi, M. 3D-fibroblast tissues constructed by a cell-coat technology enhance tight-junction formation of human colon epithelial cells. Biochem. Biophys. Res. Commun. 2015, 457, 363–369. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, W.; Chen, W.; Sun, L.; Li, X.; Zhang, C.; Yang, H. Gdnf is involved in the barrier-inducing effect of enteric glial cells on intestinal epithelial cells under acute ischemia reperfusion stimulation. Mol. Neurobiol. 2014, 50, 274–289. [Google Scholar] [CrossRef]
- Cheadle, G.A.; Costantini, T.W.; Bansal, V.; Eliceiri, B.P.; Coimbra, R. Cholinergic signaling in the gut: A novel mechanism of barrier protection through activation of enteric glia cells. Surg. Infect. (Larchmt) 2014, 15, 387–393. [Google Scholar] [CrossRef]
- Heidemann, J.; Domschke, W.; Kucharzik, T.; Maaser, C. Intestinal microvascular endothelium and innate immunity in inflammatory bowel disease: A second line of defense? Infect. Immun. 2006, 74, 5425–5432. [Google Scholar] [CrossRef]
- Hatoum, O.A.; Heidemann, J.; Binion, D.G. The intestinal microvasculature as a therapeutic target in inflammatory bowel disease. Ann. N. Y. Acad. Sci. 2006, 1072, 78–97. [Google Scholar] [CrossRef]
- Patel, R.T.; Pall, A.A.; Adu, D.; Keighley, M.R. Circulating soluble adhesion molecules in inflammatory bowel disease. Eur. J. Gastroenterol. Hepatol. 1995, 7, 1037–1041. [Google Scholar] [CrossRef]
- Dignat-George, F.; Boulanger, C.M. The many faces of endothelial microparticles. Arterioscl. Throm. Vasc. 2011, 31, 27–33. [Google Scholar] [CrossRef]
- Lansink, M.O.; Patyk, V.; de Groot, H.; Effenberger-Neidnicht, K. Melatonin reduces changes to small intestinal microvasculature during systemic inflammation. J. Surg. Res. 2017, 211, 114–125. [Google Scholar] [CrossRef]
- Huda, S.A.; Pramanik, A.K.; Ganta, V.C.; Jennings, M.; Alexander, J.S. Metabolic-hypoxic modulation of cytokine induction of intestinal endothelial adhesion molecules: Relevance to ischemic injury mediated necrotizing enterocolitis? Pathophysiology 2019, 26, 169–174. [Google Scholar] [CrossRef]
- Kasper, J.Y.; Hermanns, M.I.; Cavelius, C.; Kraegeloh, A.; Jung, T.; Danzebrink, R.; Unger, R.E.; Kirkpatrick, C.J. The role of the intestinal microvasculature in inflammatory bowel disease: Studies with a modified caco-2 model including endothelial cells resembling the intestinal barrier in vitro. Int. J. Nanomed. 2016, 11, 6353–6364. [Google Scholar] [CrossRef]
- Tietjen, G.T.; Hosgood, S.A.; DiRito, J.; Cui, J.; Deep, D.; Song, E.; Kraehling, J.R.; Piotrowski-Daspit, A.S.; Kirkiles-Smith, N.C.; Al-Lamki, R.; et al. Nanoparticle targeting to the endothelium during normothermic machine perfusion of human kidneys. Sci. Transl. Med. 2017, 9, eaam6764. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Madsen, K.; Doyle, J.; Meddings, J. Reducing small intestinal permeability attenuates colitis in the il10 gene-deficient mouse. Gut 2009, 58, 41–48. [Google Scholar] [CrossRef]
- Berman, L.; Moss, R.L. Necrotizing enterocolitis: An update. Semin. Fetal Neonatal Med. 2011, 16, 145–150. [Google Scholar] [CrossRef]
- Clark, J.A.; Doelle, S.M.; Halpern, M.D.; Saunders, T.A.; Holubec, H.; Dvorak, K.; Boitano, S.A.; Dvorak, B. Intestinal barrier failure during experimental necrotizing enterocolitis: Protective effect of egf treatment. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G938–G949. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of tlr-4 and cd14. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef]
- Cush, J.J.; Rothlein, R.; Lindsley, H.B.; Mainolfi, E.A.; Lipsky, P.E. Increased levels of circulating intercellular adhesion molecule 1 in the sera of patients with rheumatoid arthritis. Arthritis Rheum. 1993, 36, 1098–1102. [Google Scholar] [CrossRef]
- Littler, A.J.; Buckley, C.D.; Wordsworth, P.; Collins, I.; Martinson, J.; Simmons, D.L. A distinct profile of six soluble adhesion molecules (icam-1, icam-3, vcam-1, e-selectin, l-selectin and p-selectin) in rheumatoid arthritis. Br. J. Rheumatol. 1997, 36, 164–169. [Google Scholar] [CrossRef]
- Staunton, D.E.; Dustin, M.L.; Erickson, H.P.; Springer, T.A. The arrangement of the immunoglobulin-like domains of icam-1 and the binding sites for lfa-1 and rhinovirus. Cell 1990, 61, 243–254. [Google Scholar] [CrossRef]
- Wenisch, C.; Myskiw, D.; Gessl, A.; Graninger, W. Circulating selectins, intercellular adhesion molecule-1, and vascular cell adhesion molecule-1 in hyperthyroidism. J. Clin. Endocrinol. Metab. 1995, 80, 2122–2126. [Google Scholar]
- Mrowka, C.; Sieberth, H.G. Circulating adhesion molecules icam-1, vcam-1 and e-selectin in systemic vasculitis: Marked differences between wegener’s granulomatosis and systemic lupus erythematosus. Clin. Investig. 1994, 72, 762–768. [Google Scholar] [CrossRef]
- Blann, A.D.; Steele, C.; McCollum, C.N. The influence of smoking and of oral and transdermal nicotine on blood pressure, and haematology and coagulation indices. Thromb. Haemost. 1997, 78, 1093–1096. [Google Scholar] [CrossRef]
- Rohde, L.E.; Hennekens, C.H.; Ridker, P.M. Cross-sectional study of soluble intercellular adhesion molecule-1 and cardiovascular risk factors in apparently healthy men. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1595–1599. [Google Scholar] [CrossRef]
- Bhatti, M.; Chapman, P.; Peters, M.; Haskard, D.; Hodgson, H.J. Visualising e-selectin in the detection and evaluation of inflammatory bowel disease. Gut 1998, 43, 40–47. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Brynskov, J.; Vainer, B. Increased mucosal concentrations of soluble intercellular adhesion molecule-1 (sicam-1), se-selectin, and interleukin-8 in active ulcerative colitis. Dig. Dis. Sci. 1996, 41, 1780–1785. [Google Scholar] [CrossRef]
- Witkowska, A.M.; Borawska, M.H. Soluble intercellular adhesion molecule-1 (sicam-1): An overview. Eur. Cytokine Netw. 2004, 15, 91–98. [Google Scholar]
- Dragovic, R.A.; Gardiner, C.; Brooks, A.S.; Tannetta, D.S.; Ferguson, D.J.; Hole, P.; Carr, B.; Redman, C.W.; Harris, A.L.; Dobson, P.J.; et al. Sizing and phenotyping of cellular vesicles using nanoparticle tracking analysis. Nanomedicine 2011, 7, 780–788. [Google Scholar] [CrossRef]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borras, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A compendium for extracellular vesicles with continuous community annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef]
- Urbanelli, L.; Magini, A.; Buratta, S.; Brozzi, A.; Sagini, K.; Polchi, A.; Tancini, B.; Emiliani, C. Signaling pathways in exosomes biogenesis, secretion and fate. Genes 2013, 4, 152–170. [Google Scholar] [CrossRef]
- de Jong, O.G.; van Balkom, B.W.; Gremmels, H.; Verhaar, M.C. Exosomes from hypoxic endothelial cells have increased collagen crosslinking activity through up-regulation of lysyl oxidase-like 2. J. Cell. Mol. Med. 2016, 20, 342–350. [Google Scholar] [CrossRef]
- Lang, H.L.; Hu, G.W.; Chen, Y.; Liu, Y.; Tu, W.; Lu, Y.M.; Wu, L.; Xu, G.H. Glioma cells promote angiogenesis through the release of exosomes containing long non-coding rna pou3f3. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 959–972. [Google Scholar]
- Liu, X.; Li, Q.; Niu, X.; Hu, B.; Chen, S.; Song, W.; Ding, J.; Zhang, C.; Wang, Y. Exosomes secreted from human-induced pluripotent stem cell-derived mesenchymal stem cells prevent osteonecrosis of the femoral head by promoting angiogenesis. Int. J. Biol. Sci. 2017, 13, 232–244. [Google Scholar] [CrossRef]
- Ma, J.; Zhao, Y.; Sun, L.; Sun, X.; Zhao, X.; Sun, X.; Qian, H.; Xu, W.; Zhu, W. Exosomes derived from akt-modified human umbilical cord mesenchymal stem cells improve cardiac regeneration and promote angiogenesis via activating platelet-derived growth factor d. Stem Cells Transl. Med. 2017, 6, 51–59. [Google Scholar] [CrossRef]
- Han, K.Y.; Tran, J.A.; Chang, J.H.; Azar, D.T.; Zieske, J.D. Potential role of corneal epithelial cell-derived exosomes in corneal wound healing and neovascularization. Sci. Rep. 2017, 7, 40548. [Google Scholar] [CrossRef]
- Fevrier, B.; Raposo, G. Exosomes: Endosomal-derived vesicles shipping extracellular messages. Curr. Opin. Cell Biol 2004, 16, 415–421. [Google Scholar] [CrossRef]
- Schorey, J.S.; Bhatnagar, S. Exosome function: From tumor immunology to pathogen biology. Traffic 2008, 9, 871–881. [Google Scholar] [CrossRef]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Lee, H.M.; Choi, E.J.; Kim, J.H.; Kim, T.D.; Kim, Y.K.; Kang, C.; Gho, Y.S. A membranous form of icam-1 on exosomes efficiently blocks leukocyte adhesion to activated endothelial cells. Biochem. Biophys. Res. Commun. 2010, 397, 251–256. [Google Scholar] [CrossRef]
- Meyer, D.M.; Dustin, M.L.; Carron, C.P. Characterization of intercellular adhesion molecule-1 ectodomain (sicam-1) as an inhibitor of lymphocyte function-associated molecule-1 interaction with icam-1. J. Immunol. 1995, 155, 3578–3584. [Google Scholar]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (misev2018): A position statement of the international society for extracellular vesicles and update of the misev2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Kasper, J.; Hermanns, M.I.; Bantz, C.; Koshkina, O.; Lang, T.; Maskos, M.; Pohl, C.; Unger, R.E.; Kirkpatrick, C.J. Interactions of silica nanoparticles with lung epithelial cells and the association to flotillins. Arch. Toxicol. 2013, 87, 1053–1065. [Google Scholar] [CrossRef]
- Kasper, J.; Hermanns, M.I.; Bantz, C.; Utech, S.; Koshkina, O.; Maskos, M.; Brochhausen, C.; Pohl, C.; Fuchs, S.; Unger, R.E.; et al. Flotillin-involved uptake of silica nanoparticles and responses of an alveolar-capillary barrier in vitro. Eur. J. Pharm. Biopharm. 2013, 84, 275–287. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, K.; Lim, D.; Kim, K.H.; Kim, H.S.; Lee, S.; Song, J.H.; Moon, B.G.; Choy, H.E.; Park, S.C. Microvasculature remodeling in the mouse lower gut during inflammaging. Sci. Rep. 2017, 7, 39848. [Google Scholar] [CrossRef]
- Ho, Y.T.; Adriani, G.; Beyer, S.; Nhan, P.T.; Kamm, R.D.; Kah, J.C.Y. A facile method to probe the vascular permeability of nanoparticles in nanomedicine applications. Sci. Rep. 2017, 7, 707. [Google Scholar] [CrossRef]
- Freese, C.; Schreiner, D.; Anspach, L.; Bantz, C.; Maskos, M.; Unger, R.E.; Kirkpatrick, C.J. In vitro investigation of silica nanoparticle uptake into human endothelial cells under physiological cyclic stretch. Part. Fibre Toxicol. 2014, 11, 68. [Google Scholar] [CrossRef]
- Masuzawa, M.; Fujimura, T.; Hamada, Y.; Fujita, Y.; Hara, H.; Nishiyama, S.; Katsuoka, K.; Tamauchi, H.; Sakurai, Y. Establishment of a human hemangiosarcoma cell line (iso-has). Int. J. Cancer 1999, 81, 305–308. [Google Scholar] [CrossRef]
- Unger, R.E.; Krump-Konvalinkova, V.; Peters, K.; Kirkpatrick, C.J. In vitro expression of the endothelial phenotype: Comparative study of primary isolated cells and cell lines, including the novel cell line hpmec-st1.6r. Microvasc. Res. 2002, 64, 384–397. [Google Scholar] [CrossRef]
- Kasper, J.; Hermanns, M.I.; Bantz, C.; Maskos, M.; Stauber, R.; Pohl, C.; Unger, R.E.; Kirkpatrick, J.C. Inflammatory and cytotoxic responses of an alveolar-capillary coculture model to silica nanoparticles: Comparison with conventional monocultures. Part. Fibre Toxicol. 2011, 8, 6. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasper, J.Y.; Hermanns, M.I.; Kraegeloh, A.; Roth, W.; Kirkpatrick, C.J.; Unger, R.E. In Vitro Entero-Capillary Barrier Exhibits Altered Inflammatory and Exosomal Communication Pattern after Exposure to Silica Nanoparticles. Int. J. Mol. Sci. 2019, 20, 3301. https://doi.org/10.3390/ijms20133301
Kasper JY, Hermanns MI, Kraegeloh A, Roth W, Kirkpatrick CJ, Unger RE. In Vitro Entero-Capillary Barrier Exhibits Altered Inflammatory and Exosomal Communication Pattern after Exposure to Silica Nanoparticles. International Journal of Molecular Sciences. 2019; 20(13):3301. https://doi.org/10.3390/ijms20133301
Chicago/Turabian StyleKasper, Jennifer Y., M. Iris Hermanns, Annette Kraegeloh, W. Roth, C. James Kirkpatrick, and Ronald E. Unger. 2019. "In Vitro Entero-Capillary Barrier Exhibits Altered Inflammatory and Exosomal Communication Pattern after Exposure to Silica Nanoparticles" International Journal of Molecular Sciences 20, no. 13: 3301. https://doi.org/10.3390/ijms20133301
APA StyleKasper, J. Y., Hermanns, M. I., Kraegeloh, A., Roth, W., Kirkpatrick, C. J., & Unger, R. E. (2019). In Vitro Entero-Capillary Barrier Exhibits Altered Inflammatory and Exosomal Communication Pattern after Exposure to Silica Nanoparticles. International Journal of Molecular Sciences, 20(13), 3301. https://doi.org/10.3390/ijms20133301