Protein Expression Analysis in Reversible Photobleached Cells of Scenedesmus vacuolatus after High Temperature Stress


Abstract
1. Introduction
2. Results
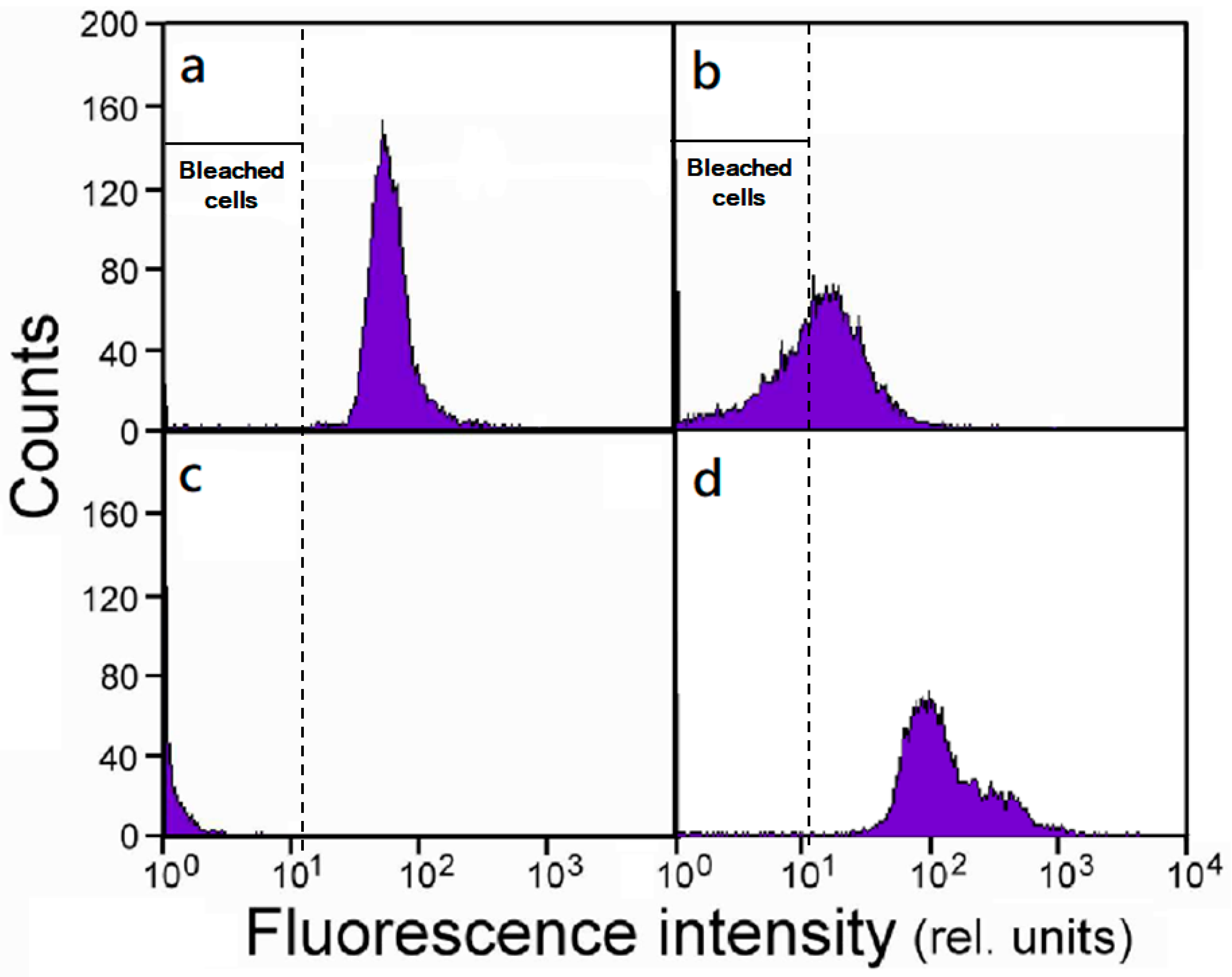
2.1. Population Responses.
2.2. Morphological Studies of Thylakoid Membranes.
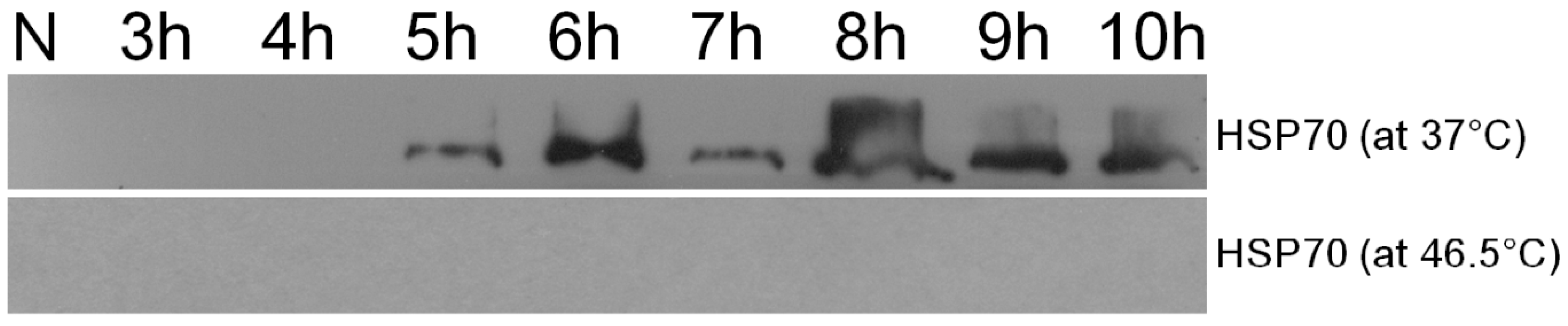
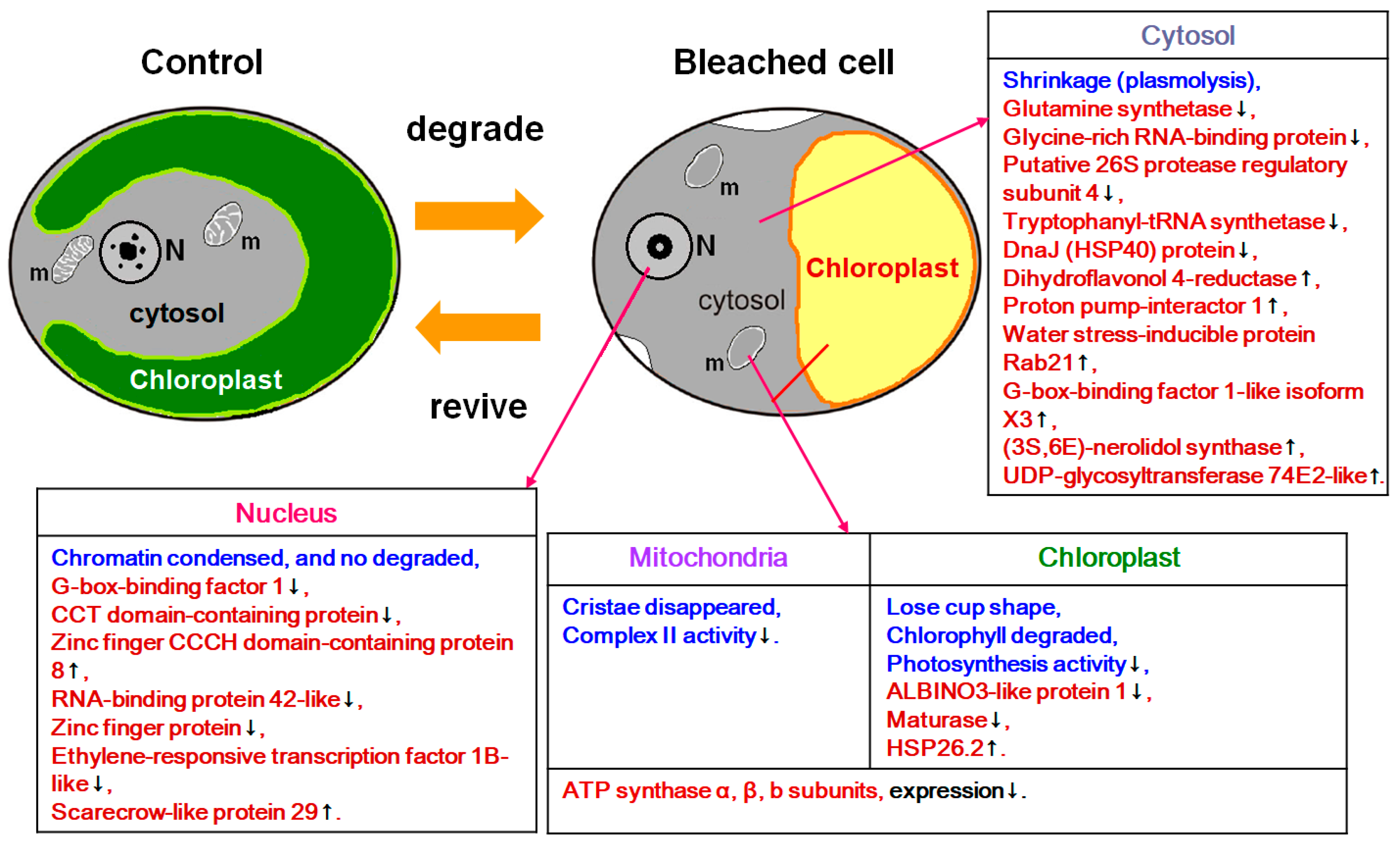
2.3. Proteins Expression Analysis.
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Heat Treatment.
4.2. Flow Cytometry Analysis.
4.3. Transmission Electron Microscopy (TEM).
4.4. Two Dimensional Gel Electrophoresis (2DE).
4.5. Protein Expression Level Analysis.
4.6. In-gel Digestion.
4.7. Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS).
4.8. Matrix Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry (MALDI-TOF MS).
4.9. Protein Identification.
4.10. Western Blotting Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| 2DE | Two-dimensional gel electrophoresis |
| ABA | Abscisic aci |
| HSP | Heat shock protein |
| LC-MS/MS | Liquid chromatography–tandem mass spectrometry |
| LHC | Light harvesting complex |
| MALDI-TOF MS | Matrix assisted laser desorption ionization–time of flight mass spectrometry |
| ROS | Reactive oxygen species |
| PSII | Photosystem II |
| TEM | Transmission electron microscopy |
References
- Gray, D.W.; Lewis, L.A.; Cardon, Z.G. Photosynthetic recovery following desiccation of desert green algae (Chlorophyta) and their aquatic relatives. Plant. Cell Environ. 2007, 30, 1240–1255. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, A.; Herburger, K.; Blaas, K.; Lewis, L.A.; Karsten, U. The terrestrial green macroalga Prasiola calophylla (Trebouxiophyceae, Chlorophyta): ecophysiological performance under water-limiting conditions. Protoplasma 2017, 254, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Agrawal, S.C. Survival of blue-green and green algae under stress conditions. Folia Microbiol. 2006, 51, 121–128. [Google Scholar] [CrossRef]
- Aoki, S.; Hase, E. De- and re-generation of chloroplasts in the cells of Chlorella protothecoides. I. Syntheses of nucleic acids and protein in relation to the process of regeneration of chloroplast. Plant. Cell Physiol. 1964, 5, 473–484. [Google Scholar]
- Torzillo, G.; Pushparaj, B.; Bocci, F.; Balloni, W.; Materassi, R.; Florenzano, G. Production of Spirulina biomass in closed photobioreactors. Biomass 1986, 11, 61–74. [Google Scholar] [CrossRef]
- Tredici, M.R.; Materassi, R. From open ponds to vertical alveolar panels: the Italian experience in the development of reactors for the mass cultivation of phototrophic microorganisms. J. Appl. Phycol. 1992, 4, 221–231. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: an overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Feller, U.; Crafts-Brandner, S.J.; Salvucci, M.E. Moderately high temperatures inhibit ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activase-mediated activation of Rubisco. Plant. Physiol. 1998, 116, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Temple, M.D.; Perrone, G.G.; Dawes, I.W. Complex cellular responses to reactive oxygen species. Trends Plant. Sci. 2005, 15, 319–326. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Møller, I.M.; Sweetlove, L.J. ROS signaling-specificity is required. Trends Plant. Sci. 2010, 15, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant. Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant. Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.C.; Hsu, B.D. The reversible disintegration of heat-treated Scenedesmus vacuolatus under continuous light cultivation conditions. Protoplasma 2014, 251, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Hemme, D.; Veyel, D.; Mühlhaus, T.; Sommer, F.; Jüppner, J.; Unger, A.-K.; Sandmann, M.; Fehrle, I.; Schönfelder, S.; Steup, M.; et al. Systems-wide analysis of acclimation responses to long-term heat stress and recovery in the photosynthetic model organism Chlamydomonas reinhardtii. Plant. Cell 2014, 26, 4270–4297. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.C.; Hsu, B.D. Characterization of the decline and recovery of heat-treated Scenedesmus vacuolatus. Bot. Stud. 2013, 54, 3. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matile, P.; Hörtensteiner, S.; Thomas, H. Chlorophyll degradation. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1999, 50, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature stress. Front. Plant. Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Takagi, D.; Takumi, S.; Hashiguchi, M.; Sejima, T.; Miyake, C. Superoxide and singlet oxygen produced within the thylakoid membranes both cause photosystem I photoinhibition. Plant. Physiol. 2016, 171, 1626–1634. [Google Scholar] [CrossRef]
- Vincent, D.; Zivy, M. Plant proteome responses to abiotic stress. In Plant Proteomics; Samaj, J., Thelen, J.J., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2007; pp. 346–364. [Google Scholar]
- Taylor, A.O.; Craig, A.S. Plants under climatic stress. II. Low temperature, high light effects on chloroplast ultrastructure. Plant. Physiol. 1971, 47, 719–725. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Jubany-Marí, T.; Alegre, L. Drought-induced senescence is characterized by loss of antioxidant defences in chloroplasts. Plant. Cell Environ. 2001, 24, 1319–1327. [Google Scholar] [CrossRef]
- Yamane, K.; Rahman, M.S.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Pretreatment with antioxidants decreases the effects of salt stress on chloroplast ultrastructure in rice leaf segments (Oryza sativa L.). Plant. Prod. Sci. 2004, 7, 292–300. [Google Scholar] [CrossRef]
- Park, J.H.; Tran, L.H.; Jung, S. Perturbations in the photosynthetic pigment status result in photooxidation-induced crosstalk between carotenoid and porphyrin biosynthetic pathways. Front. Plant. Sci. 2017, 8, 1992. [Google Scholar] [CrossRef]
- Ibba, M.; Soll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef]
- Bellafiore, S.; Ferris, P.; Naver, H.; Goehre, V.; Rochaix, J.D. Loss of Albino3 leads to the specific depletion of the light-harvesting system. Plant. Cell 2002, 14, 2303–2314. [Google Scholar] [CrossRef]
- Mahong, B.; Roytrakul, S.; Phaonaklop, N.; Wongratana, J.; Yokthongwattana, K. Proteomic analysis of a model unicellular green alga, Chlamydomonas reinhardtii, during short-term exposure to irradiance stress reveals significant down regulation of several heat-shock proteins. Planta 2012, 235, 499–511. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, N.; Nijhavan, A.; Khurana, J.P.; Khurana, P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant. Cell Environ. 2012, 35, 1912–1931. [Google Scholar] [CrossRef]
- Hu, X.; Yang, Y.; Gong, F.; Zhang, D.; Zhang, L.; Wu, L.; Li, C.; Wang, W. Protein sHSP26 improves chloroplast performance under heat stress by interacting with specific chloroplast proteins in maize (Zea mays). J. Proteomics 2015, 115, 81–92. [Google Scholar] [CrossRef]
- Guo, S.J.; Zhou, H.Y.; Zhang, X.S.; Li, X.G.; Meng, Q.W. Overexpression of CaHSP26 in transgenic tobacco alleviates photoinhibition of PSII and PSI during chilling stress under low irradiance. J. Plant. Physiol. 2007, 164, 126–136. [Google Scholar] [CrossRef]
- Kim, K.H.; Alam, I.; Kim, Y.G.; Sharmin, S.A.; Lee, K.W.; Lee, S.H.; Lee, B.H. Overexpression of a chloroplast-localized small heat shock protein OsHSP26 confers enhanced tolerance against oxidative and heat stresses in tall fescue. Biotechnol. Lett. 2012, 34, 371–377. [Google Scholar] [CrossRef]
- Mundy, J.; Chua, N.H. Abscisic acid and water-stress induce the expression of a novel rice gene. EMBO J. 1988, 7, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Hirai, N.; Kurimura, Y.; Ohigashi, H.; Tsuji, Y. Abscisic acid-dependent algal morphogenesis in the unicellular green alga Haematococcus pluvialis. Plant. Growth Regul. 1997, 22, 79–85. [Google Scholar] [CrossRef]
- Yoshida, K.; Igarashi, E.; Mukai, M.; Hirata, K.; Miyamoto, K. Induction of tolerance to oxidative stress in the green alga, Chlamydomonas reinhardtii, by abscisic acid. Plant. Cell Environ. 2003, 26, 451–457. [Google Scholar] [CrossRef]
- Yoshida, K.; Igarashi, E.; Wakatsuki, E.; Miyamoto, K.; Hirata, K. Mitigation of osmotic and salt stresses by abscisic acid through reduction of stress-derived oxidative damage in Chlamydomonas reinhardtii. Plant. Sci. 2004, 167, 1335–1341. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, J. Phytohormones in microalgae: a new opportunity for microalgal biotechnology? Trends Plant. Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef]
- Holzinger, A.; Pichrtová, M. Abiotic stress tolerance of charophyte green algae: new challenges for omics techniques. Front. Plant. Sci. 2016, 7, 678. [Google Scholar] [CrossRef]
- Park, W.K.; Yoo, G.; Moon, M.; Kim, C.W.; Choi, Y.E.; Yang, J.W. Phytohormone supplementation significantly increases growth of Chlamydomonas reinhardtii cultivated for biodiesel production. Appl. Biochem. Biotechnol. 2013, 171, 1128–1142. [Google Scholar] [CrossRef]
- Tatiana, A.K.; Bruce, P.H.; Priti, K.; Levin, B.D. Effect of phytohormones on growth and accumulation of pigments and fatty acids in the microalgae Scenedesmus quadricauda. Algal Res. 2017, 27, 325–334. [Google Scholar]
- García, M.N.M.; País, S.M.; Téllez-Iñón, M.T.; Capiati, D.A. Characterization of StPPI1, a proton pump interactor from Solanum tuberosum L. that is up-regulated during tuber development and by abiotic stress. Planta 2011, 233, 661–674. [Google Scholar] [CrossRef]
- Morandini, P.; Valera, M.; Albumi, C.; Bonza, M.; Giacometti, S.; Ravera, G.; Murgia, I.; Soave, C.; DeMichelis, M. A novel interaction partner for C-terminus of Arabidopsis thaliana plasma membrane H+-ATPase (AHA1 isoform): site and mechanism of action on H+-ATPase activity differ from those of 14-3-3 proteins. Plant. J. 2002, 31, 487–497. [Google Scholar] [CrossRef]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T. Plasma membrane H+-ATPase regulation in the center of plant physiology. Mol. Plant. 2016, 9, 323–337. [Google Scholar] [CrossRef]
- Hayashi, M.; Inoue, S.; Takahashi, K.; Kinoshita, T. Immunohistochemical detection of blue light-induced phosphorylation of the plasma membrane H+-ATPase in stomatal guard cells. Plant. Cell Physiol. 2011, 52, 1238–1248. [Google Scholar] [CrossRef]
- Janicka-Russak, M.; Kłobus, G. Modification of plasma membrane and vacuolar H+-ATPase in response to NaCl and ABA. J. Plant. Physiol. 2007, 164, 295–302. [Google Scholar] [CrossRef]
- Zhang, D.; Koay, E.S.C. Analysis of laser capture microdissected cells by 2-dimensional gel electrophoresis. In Clinical Proteomics, Methods and Protocols; Vlahou, A., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 77–91. [Google Scholar]
- Shevchenko, A.; Tomas, H.; Havliš, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Chan, H.L.; Chou, H.C.; Duran, M.C.; Gruenewald, J.; Waterfield, M.D.; Ridley, A.; Timms, J.F. Major role of EGFR and SRC kinases in promoting oxidative stress-dependent loss of adhesion and apoptosis in epithelial cells. J. Biol. Chem. 2010, 285, 4307–4318. [Google Scholar] [CrossRef]
- Lai, T.C.; Chou, H.C.; Chen, Y.W.; Lee, T.R.; Chan, H.T.; Shen, H.H.; Lee, W.T.; Lin, S.T.; Lu, Y.C.; Wu, C.L.; et al. Secretomic and proteomic analysis of potential breast cancer markers by two-dimensional differential gel electrophoresis. J. Proteome. Res. 2010, 9, 1302–1322. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Homologous Protein (LC/MS/MS) | Obs/Theor M. W. (kD) | Obs/Theor PI | Accession No. | Mascot Score | Identified Peptide Fragment | Location | Functional Description | Expression Fold Change (Bleached/Control) |
|---|---|---|---|---|---|---|---|---|---|
| 118 | ATP synthase β subunit [Spathelia wrightii] | 55/50.118 | 5.52/5.14 | CBY80088.1 | 67 | K.IGLFGGAGTGK.T | Chloroplast and mitochondria | Produces ATP from ADP. The catalytic sites are primarily by the subunits. | 0.02 |
| 130 | Glutamine synthetase [Chlorella sorokiniana] | 42/41.671 | 6.06/6.17 | PRW34037.1 | 79 | K.HETCDINTFR.F | Cytosol | Catalyzes the ATP-dependent biosynthesis of glutamine from glutamate and ammonia. | 0.10 |
| 178 | Dihydroflavonol 4-reductase [Vitis davidii] | 28/38.139 | 5.86/5.81 | AHK10250.1 | 66 | R.ATVRDPTNVK.K | Extrinsic component of endoplasmic reticulum | This protein is involved in flavonoid metabolism. | 1.13 |
| 199 | Proton pump-interactor 1 [Zea mays] | 170/170.178 | 4.48/4.59 | ONM22629.1 | 60 | K.AENMVEVKSAAR.E | Endoplasmic reticulum and plasma membrane | Regulation of proton transport. Enhances H+ATPase activity. | 4.82 |
| 205 | Maturase K (chloroplast) [Trochodendron aralioides] | 28/28.035 | 4.57/9.98 | AAB58649.1 | 77 | R.SQMIENAFLIDSTSKKFDTIVPISPLIGSLAK.A | Chloroplast | Probably assists in splicing its own and other chloroplast group II introns. mRNA and tRNA processing. | 0.04 |
| 224 | Glycine-rich RNA-binding protein [Dorcoceras hygrometricum] | 16/16.923 | 5.46/5.54 | KZV53374.1 | 66 | R.GGGGGGYGGDR.G | Shuttling between nucleus and cytoplasm | Plays a role in RNA transcription. | 0.05 |
| 240 | ATP synthase α subunit [Eriachne mucronata] | 56/55.704 | 5.42/5.94 | YP_009410967.1 | 69 | K.TAIATNTILNQK.S | Chloroplast and mitochondria | Produces ATP from ADP. The α subunit is a regulator of ATP synthase. | 0.01 |
| No. | Homologous Protein (MALDI-TOF) | Obs/Theor M. W. (kD) | Obs/Theor PI | Accession No. | Mascot Score | No. Match Peptides (Seq. Coverage) | Location | Functional Description | Expression Fold Change (Bleached/Control) |
|---|---|---|---|---|---|---|---|---|---|
| 113 | RNA-binding protein 42-like [Erythranthe guttata] | 32/27.233 | 5.0/9.73 | XP_012840025.1 | 83 | 29(49%) | Nucleus | Essential polysome-associated RNA-binding protein. | 0.03 |
| 185 | Zinc finger protein [Macleaya cordata] | 55/55.293 | 5.5/5.58 | OUZ99355.1 | 130 | 22(15%) | Nucleus | RNA methyltransferase activity. | 0.18 |
| 188 | ALBINO3-like protein 1, chloroplastic isoform X1 [Cucumis melo] | 58/56.039 | 5.28/7.71 | XP_008444397.1 | 110 | 23(20%) | Chloroplast | Membrane insertase activity | 0.65 |
| 207 | Putative 26S protease regulatory subunit 4 [Zostera marina] | 66/68.678 | 4.47/9.12 | KMZ68817.1 | 91 | 54(50%) | Cytoplasm | Polyubiquitin modification- dependent protein binding. | Only detected in control cell |
| 216 | ATP synthase FO subunit b [Chlorokybus atmophyticus] | 25/21.021 | 5.0/9.72 | YP_001315093.1 | 105 | 20(62%) | Chloroplast and mitochondria | Connect FO, F1 of ATP synthesis. | Only detected in control cell |
| 217 | Ethylene-responsive transcription factor 1B-like [Cajanus cajan] | 24/28.294 | 5.49/5.53 | XP_020237863.1 | 102 | 17(33%) | Nucleus | DNA-binding transcription factor activity. | Only detected in control cell |
| 225 | G-box-binding factor 1 [Arabidopsis lyrata subsp. lyrata] | 33/33.716 | 5.6/6.03 | XP_002866995.1 | 86 | 24(22%) | Nucleus | Binds to the G-box motif (5’-CCACGTGG-3’) of the rbcS-1A gene promoter, and 5’-CACGTG-3’ of LHCB2.4 (At3g27690) promoter. | Only detected in control cell |
| 229 | DnaJ protein P58IPK homolog [Cucurbita pepo subsp. pepo] | 40/53.895 | 5.65/6.85 | XP_023517230.1 | 85 | 27(34%) | Endoplasmic reticulum | Chaperone (Hsp40 family). May play a role in protein folding in the endoplasmic reticulum. | Only detected in control cell |
| 230 | Tryptophanyl-tRNA synthetase [Chlorella sorokiniana] | 39/34.287 | 5.69/6.97 | PRW57418.1 | 84 | 23(62%) | Cytoplasm | Aminoacyl-tRNA ligase activity | Only detected in control cell |
| 231 | CCT domain-containing protein [Cephalotus follicularis] | 48/49.843 | 5.77/6.17 | GAV71350.1 | 83 | 29(24%) | Nucleus | Regulation of gene expression. | 0.07 |
| 301 | Water stress-inducible protein Rab21 [Setaria italica] | 16/15.165 | 4.7/10.11 | XP_004980462.1 | 148 | 17(54%) | Cytoplasm | Responsive to abscisic acid. | Only detected in bleached cell |
| 307 | Scarecrow-like protein 29 [Camelina sativa] | 28/28.317 | 4.63/5.41 | XP_010489817.1 | 82 | 20(27%) | Nucleus | DNA-binding transcription factor activity. Red, far-red light phototransduction | Only detected in bleached cell |
| 310 | (3S,6E)-nerolidol synthase 1 [Herrania umbratica] | 59/59.063 | 5.3/6.44 | XP_021294536.1 | 198 | 28(35%) | Cytosol | Stress response protein. | Only detected in bleached cell |
| 314 | G-box-binding factor 1-like isoform X3 [Tarenaya hassleriana] | 29/33.637 | 5.53/6.99 | XP_010548353.1 | 85 | 24(43%) | Nucleus | Regulates blue light-mediated photomorphogenic growth. | Only detected in bleached cell |
| 319 | Unnamed protein product [Coffea canephora] | 23/27.230 | 6.95/5.78 | CDP02738.1 | 92 | 23(34%) | N/A | N/A | Only detected in bleached cell |
| 325 | UDP-glycosyltransferase 74E2-like [Hevea brasiliensis] | 26/25.751 | 6.33/4.87 | XP_021651079.1 | 82 | 18(33%) | Intracellular membrane-bounded organelle | Glycosyltransferase, transferase activity, transferring hexosyl groups | Only detected in bleached cell |
| 326 | Chloroplast low molecular weight heat shock protein HSP26.2 [Agrostis stolonifera var. palustris] | 26/26.196 | 6.5/7.85 | AAN74534.1 | 118 | 14(47%) | Chloroplast | Stress response protein. | Only detected in bleached cell |
| 327 | Uncharacterized protein LOC104232495 [Nicotiana sylvestris] | 27/28.821 | 6.7/6.15 | XP_009784020.1 | 95 | 14(24%) | N/A | N/A | Only detected in bleached cell |
| 331 | Glutathione S-transferase T2-like [Chenopodium quinoa] | 56/51.087 | 6.3/8.01 | XP_021721044.1 | 120 | 56(54%) | N/A | Transferase activity | 2.21 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, T.-H.; Leu, K.-L.; Hsu, B.-D.; Lee, T.-C. Protein Expression Analysis in Reversible Photobleached Cells of Scenedesmus vacuolatus after High Temperature Stress. Int. J. Mol. Sci. 2019, 20, 3082. https://doi.org/10.3390/ijms20123082
Ko T-H, Leu K-L, Hsu B-D, Lee T-C. Protein Expression Analysis in Reversible Photobleached Cells of Scenedesmus vacuolatus after High Temperature Stress. International Journal of Molecular Sciences. 2019; 20(12):3082. https://doi.org/10.3390/ijms20123082
Chicago/Turabian StyleKo, Tzu-Hsing, Kuen-Lin Leu, Ban-Dar Hsu, and Tzan-Chain Lee. 2019. "Protein Expression Analysis in Reversible Photobleached Cells of Scenedesmus vacuolatus after High Temperature Stress" International Journal of Molecular Sciences 20, no. 12: 3082. https://doi.org/10.3390/ijms20123082
APA StyleKo, T.-H., Leu, K.-L., Hsu, B.-D., & Lee, T.-C. (2019). Protein Expression Analysis in Reversible Photobleached Cells of Scenedesmus vacuolatus after High Temperature Stress. International Journal of Molecular Sciences, 20(12), 3082. https://doi.org/10.3390/ijms20123082