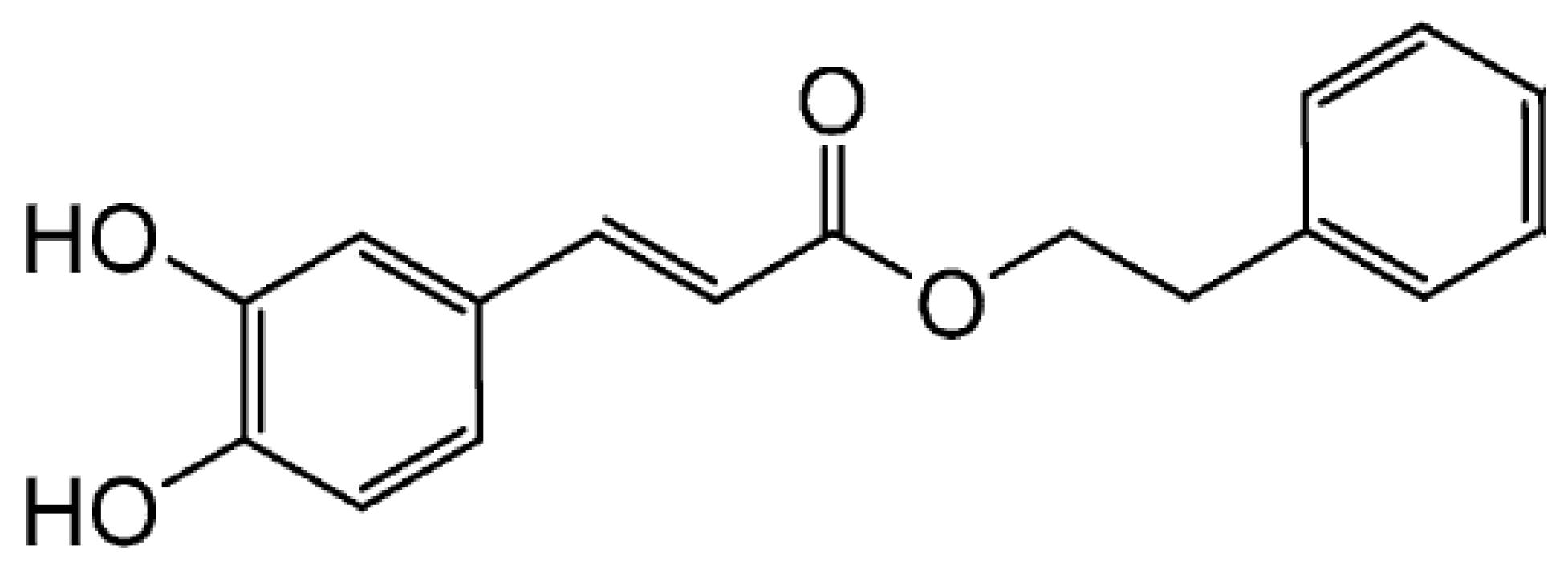
Caffeic Acid Phenethyl Ester Inhibits UV-Induced MMP-1 Expression by Targeting Histone Acetyltransferases in Human Skin

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
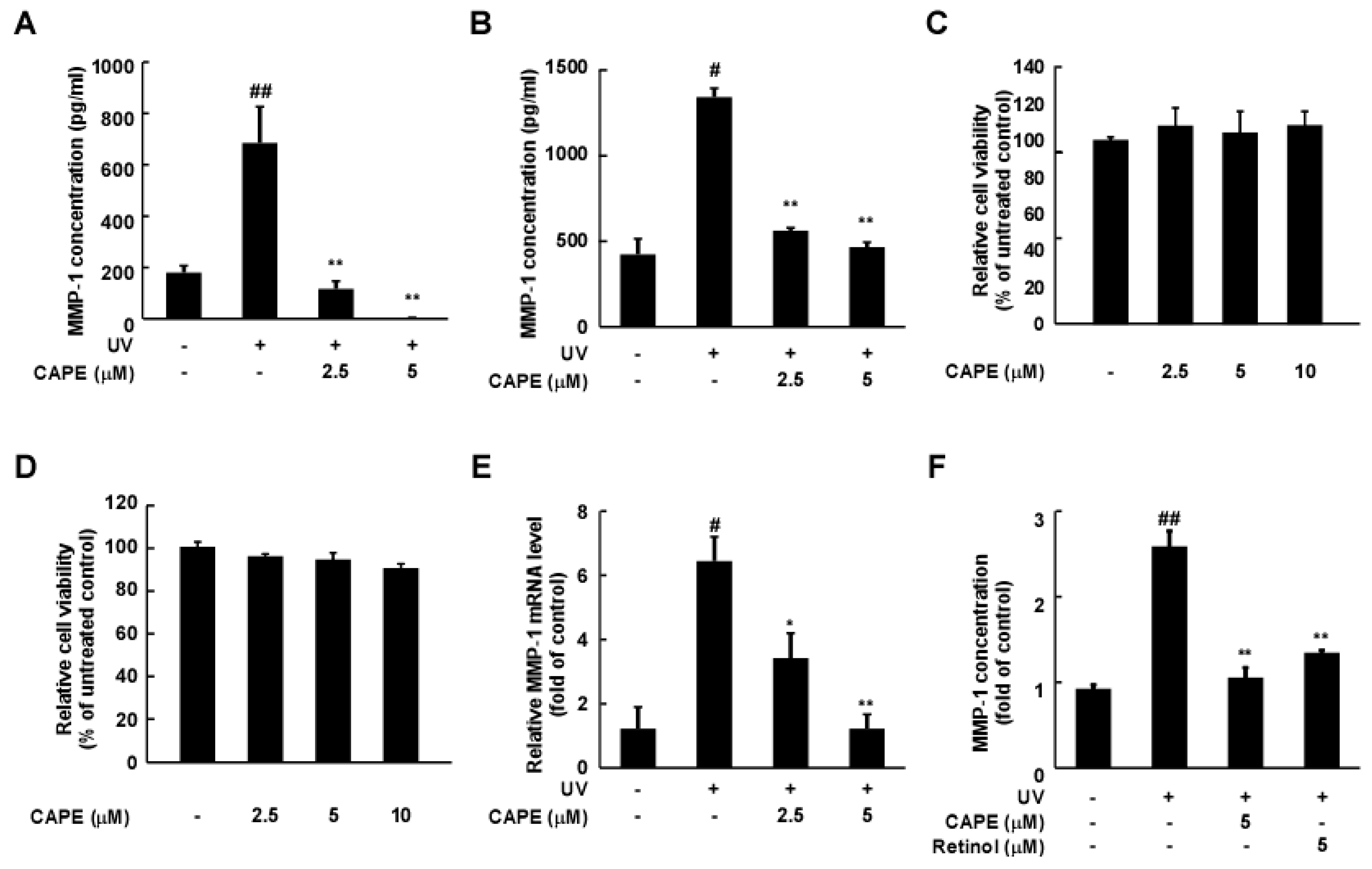
2.1. CAPE Reduces UV-Induced MMP-1 Expression in Human Dermal Fibroblasts
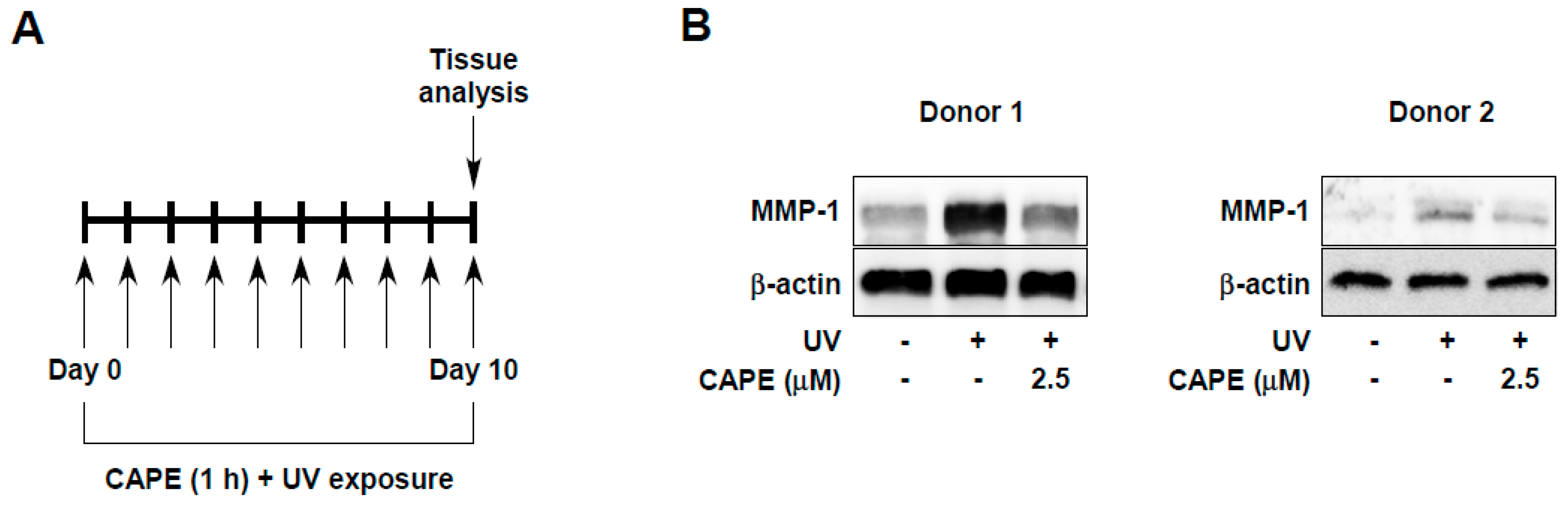
2.2. CAPE Suppresses UV-Induced MMP-1 Expression in Human Skin Tissues
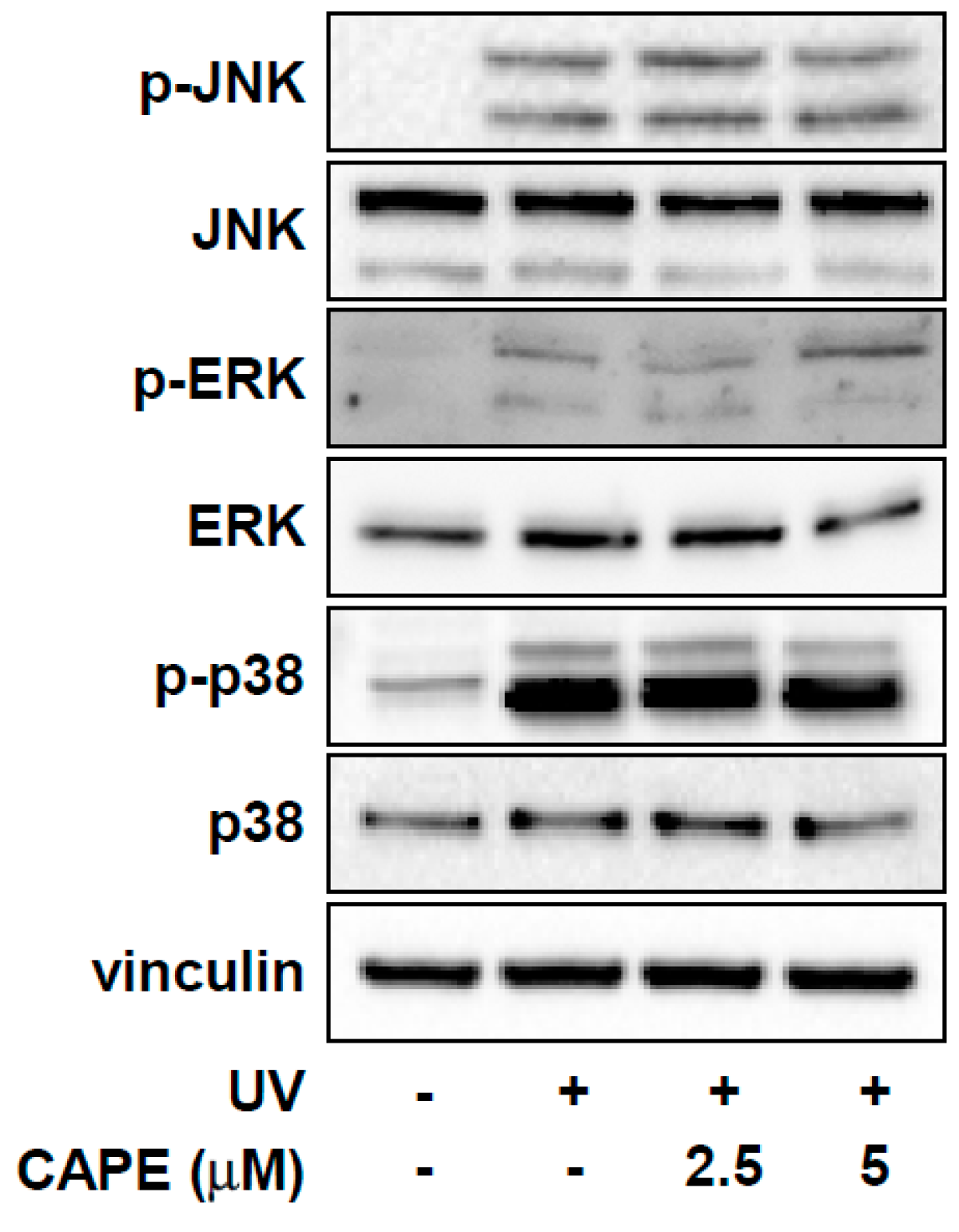
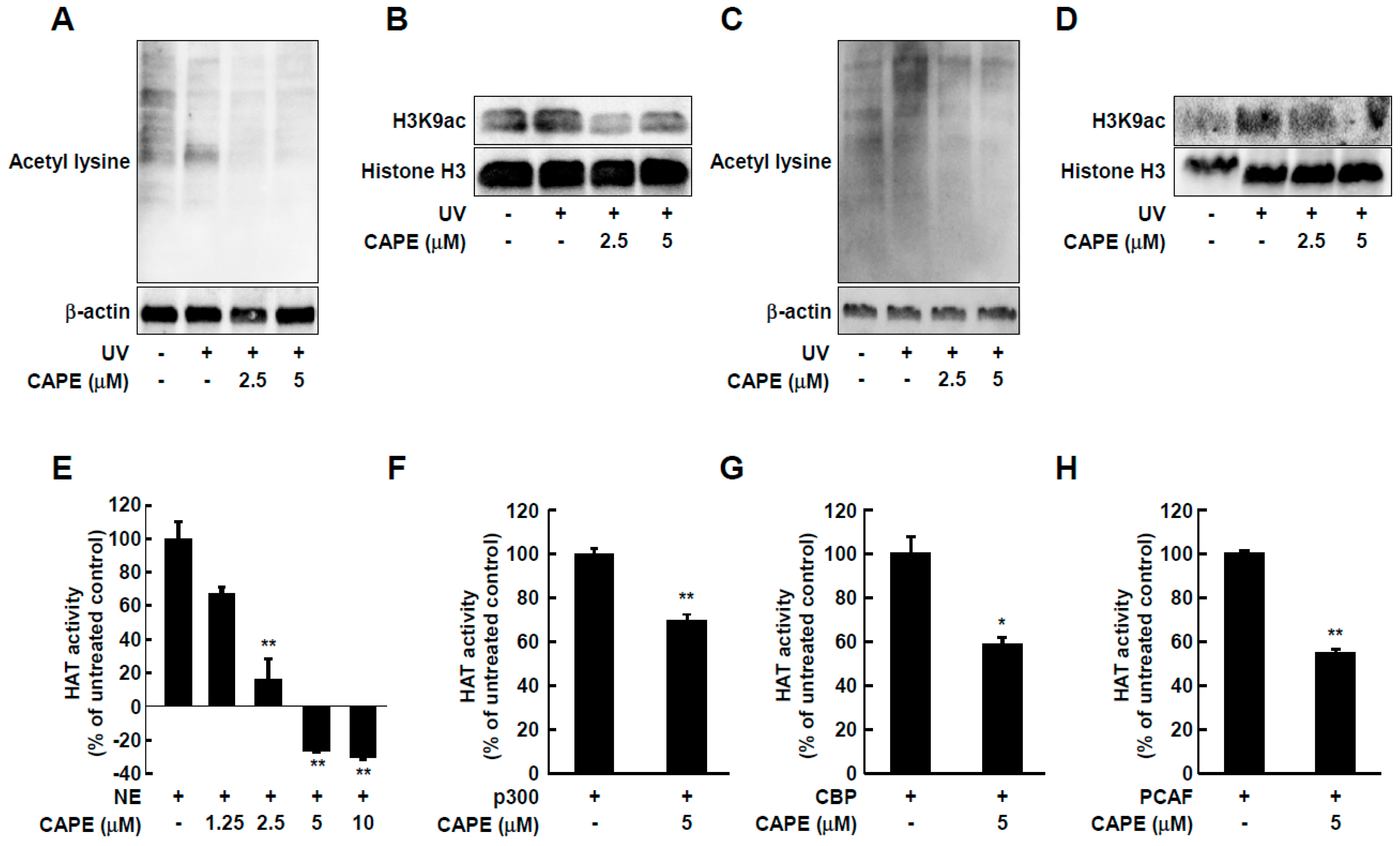
2.3. CAPE Inhibits UV-Stimulated Acetylation of Total Lysine and Histone H3 Lysine 9 in Both HDF Cells and Human Skin Tissues
2.4. CAPE Acts as an HAT Inhibitor
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and UV Irradiation
4.3. Excised Human Skin and UV Irradiation
4.4. Cell Viability Assay
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Real-Time PCR (qPCR)
4.7. Immunoblot Analysis
4.8. HAT Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CAPE | Caffeic acid phenethyl ester |
| UV | Ultraviolet |
| MMP-1 | Matrix metalloproteinase |
| HDF | Human dermal fibroblasts |
| HAT | Histone acetyltransferase |
| CBP | CREP-binding protein |
| PCAF | p300/CBP-associated factor |
| HDAC | Histone deacetylase |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular-signal-regulated kinase |
| JNK | c-Jun N-terminal kinase |
| AA | Anacardic acid |
| COX-2 | Cyclooxygenase-2 |
References
- Debacq-Chainiaux, F.; Leduc, C.; Verbeke, A.; Toussaint, O. UV, stress and aging. Dermato-Endocrinology 2012, 4, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Orioli, D.; Dellambra, E. Epigenetic Regulation of Skin Cells in Natural Aging and Premature Aging Diseases. Cells 2018, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.N.; Lee, Y.H.; Park, J.S.; Jun, W.J.; Kim, H.K.; Hong, B.S.; Shin, D.H.; Cho, H.Y. Protective effects of aucubin isolated from Eucommia ulmoides against UVB-induced oxidative stress in human skin fibroblasts. Biol. Pharm. Bull. 2005, 28, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Shankar, E.; Kanwal, R.; Candamo, M.; Gupta, S. Dietary phytochemicals as epigenetic modifiers in cancer: Promise and challenges. Semin. Cancer Biol. 2016, 40, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Peserico, A.; Simone, C. Physical and functional HAT/HDAC interplay regulates protein acetylation balance. BioMed Res. Int. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Shin, J.M.; Eun, H.C.; Chung, J.H. The role of p300 histone acetyltransferase in UV-induced histone modifications and MMP-1 gene transcription. PLoS ONE 2009, 4, e4864. [Google Scholar] [CrossRef]
- Ding, S.; Chen, J.; Zeng, Q.; Lu, J.; Tan, L.; Guo, A.; Kang, J.; Yang, S.; Xiang, Y.; Zuo, C.; et al. Chronic sun exposure is associated with distinct histone acetylation changes in human skin. Br. J. Dermatol. 2018, 179, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Lee, S.R.; Lee, E.; Kim, K.H.; Byun, S. Caffeic Acid Phenethyl Ester from the Twigs of Cinnamomum cassia Inhibits Malignant Cell Transformation by Inducing c-Fos Degradation. J. Nat. Prod. 2017, 80, 2124–2130. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Moon, S.K.; Chang, Y.C.; Ko, J.H.; Lee, Y.C.; Cho, G.; Kim, S.H.; Kim, J.G.; Kim, C.H. Novel and therapeutic effect of caffeic acid and caffeic acid phenyl ester on hepatocarcinoma cells: Complete regression of hepatoma growth and metastasis by dual mechanism. FASEB J. 2004, 18, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Lee, D.Y.; Kim, J.H.; Choi, I.; Park, S.G.; Seo, S.K.; Lee, S.W.; Lee, C.M.; Park, Y.M.; Jeon, Y.J.; et al. Anti-inflammatory activity of caffeic acid phenethyl ester (CAPE) extracted from Rhodiola sacra against lipopolysaccharide-induced inflammatory responses in mice. Process Biochem. 2008, 43, 783–787. [Google Scholar] [CrossRef]
- Kudugunti, S.K.; Vad, N.M.; Ekogbo, E.; Moridani, M.Y. Efficacy of Caffeic Acid Phenethyl Ester (CAPE) in skin B16-F0 melanoma tumor bearing C57BL/6 mice. Investig. New Drugs 2011, 29, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, G.; Karim, S.; Akram, M.R.; Khan, S.A.; Azhar, S.; Mumtaz, A.; Bin Asad, M.H. Caffeic acid phenethyl ester and therapeutic potentials. BioMed Res. Int. 2014, 2014, doi. [Google Scholar] [CrossRef] [PubMed]
- Seong, J.S.; Xuan, S.H.; Park, S.H.; Lee, K.S.; Park, Y.M.; Park, S.N. Antioxidative and Antiaging Activities and Component Analysis of Lespedeza cuneata G. Don Extracts Fermented with Lactobacillus pentosus. J. Microbiol. Biotechnol. 2017, 27, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed]
- Pollack, B.P.; Sapkota, B.; Boss, J.M. Ultraviolet Radiation-Induced Transcription is Associated with Gene-Specific Histone Acetylation. Photochem. Photobiol. 2009, 85, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Lee, D.H.; Lee, S.; Kim, E.J.; Chung, J.H. UV-induced DNA damage and histone modification may involve MMP-1 gene transcription in human skin In Vivo. J. Dermatol. Sci. 2014, 73, 169–171. [Google Scholar] [CrossRef]
- Ujfaludi, Z.; Tuzesi, A.; Majoros, H.; Rothler, B.; Pankotai, T.; Boros, I.M. Coordinated activation of a cluster of MMP genes in response to UVB radiation. Sci. Rep. 2018, 8, 2660. [Google Scholar] [CrossRef]
- Roth, S.Y.; Denu, J.M.; Allis, C.D. Histone acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef]
- Omene, C.; Kalac, M.; Wu, J.; Marchi, E.; Frenkel, K.; O’Connor, O.A. Propolis and Its Active Component, Caffeic Acid Phenethyl Ester (CAPE), Modulate Breast Cancer Therapeutic Targets via an Epigenetically Mediated Mechanism of Action. J. Cancer Sci. Ther. 2013, 5, 334–342. [Google Scholar] [PubMed]
- Shankar, S.; Kumar, D.; Srivastava, R.K. Epigenetic modifications by dietary phytochemicals: Implications for personalized nutrition. Pharmacol. Ther. 2013, 138, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Masumi, A. Histone Acetyltransferases as Regulators of Nonhistone Proteins: The Role of Interferon Regulatory Factor Acetylation on Gene Transcription. BioMed Res. Int. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Lee, S.; Kim, E.J.; Kong, K.H.; Lee, D.H.; Chung, J.H. Topical application of anacardic acid (6-nonadecyl salicylic acid) reduces UV-induced histone modification, MMP-13, MMP-9, COX-2 and TNF-alpha expressions in hairless mice skin. J. Dermatol. Sci. 2013, 70, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Han, A.; Lee, J.; Lee, M.H.; Lee, S.Y.; Shin, E.J.; Song, Y.R.; Lee, K.M.; Lee, K.W.; Lim, T.G. Sulfuretin, a natural Src family kinases inhibitor for suppressing solar UV-induced skin aging. J. Funct. Foods 2019, 52, 442–449. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, E.J.; Jo, S.; Choi, H.-k.; Choi, S.; Byun, S.; Lim, T.-G. Caffeic Acid Phenethyl Ester Inhibits UV-Induced MMP-1 Expression by Targeting Histone Acetyltransferases in Human Skin. Int. J. Mol. Sci. 2019, 20, 3055. https://doi.org/10.3390/ijms20123055
Shin EJ, Jo S, Choi H-k, Choi S, Byun S, Lim T-G. Caffeic Acid Phenethyl Ester Inhibits UV-Induced MMP-1 Expression by Targeting Histone Acetyltransferases in Human Skin. International Journal of Molecular Sciences. 2019; 20(12):3055. https://doi.org/10.3390/ijms20123055
Chicago/Turabian StyleShin, Eun Ju, Seongin Jo, Hyo-kyoung Choi, Sungbin Choi, Sanguine Byun, and Tae-Gyu Lim. 2019. "Caffeic Acid Phenethyl Ester Inhibits UV-Induced MMP-1 Expression by Targeting Histone Acetyltransferases in Human Skin" International Journal of Molecular Sciences 20, no. 12: 3055. https://doi.org/10.3390/ijms20123055
APA StyleShin, E. J., Jo, S., Choi, H.-k., Choi, S., Byun, S., & Lim, T.-G. (2019). Caffeic Acid Phenethyl Ester Inhibits UV-Induced MMP-1 Expression by Targeting Histone Acetyltransferases in Human Skin. International Journal of Molecular Sciences, 20(12), 3055. https://doi.org/10.3390/ijms20123055