Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae)

 ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
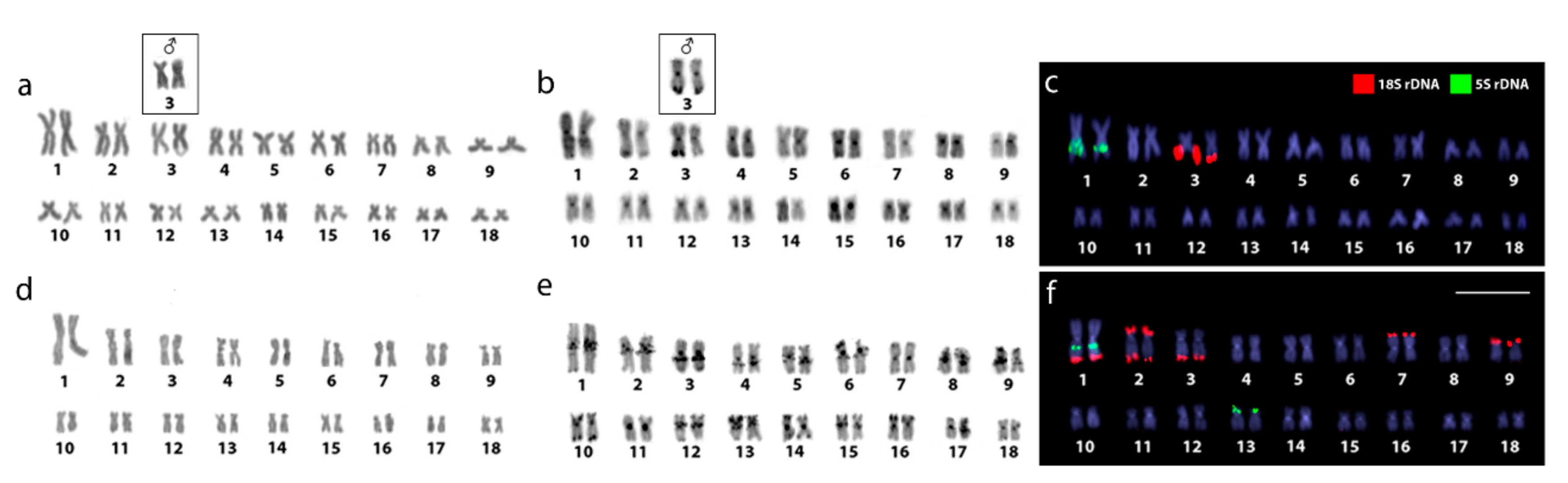
2.1. Karyotypes and C-Banding
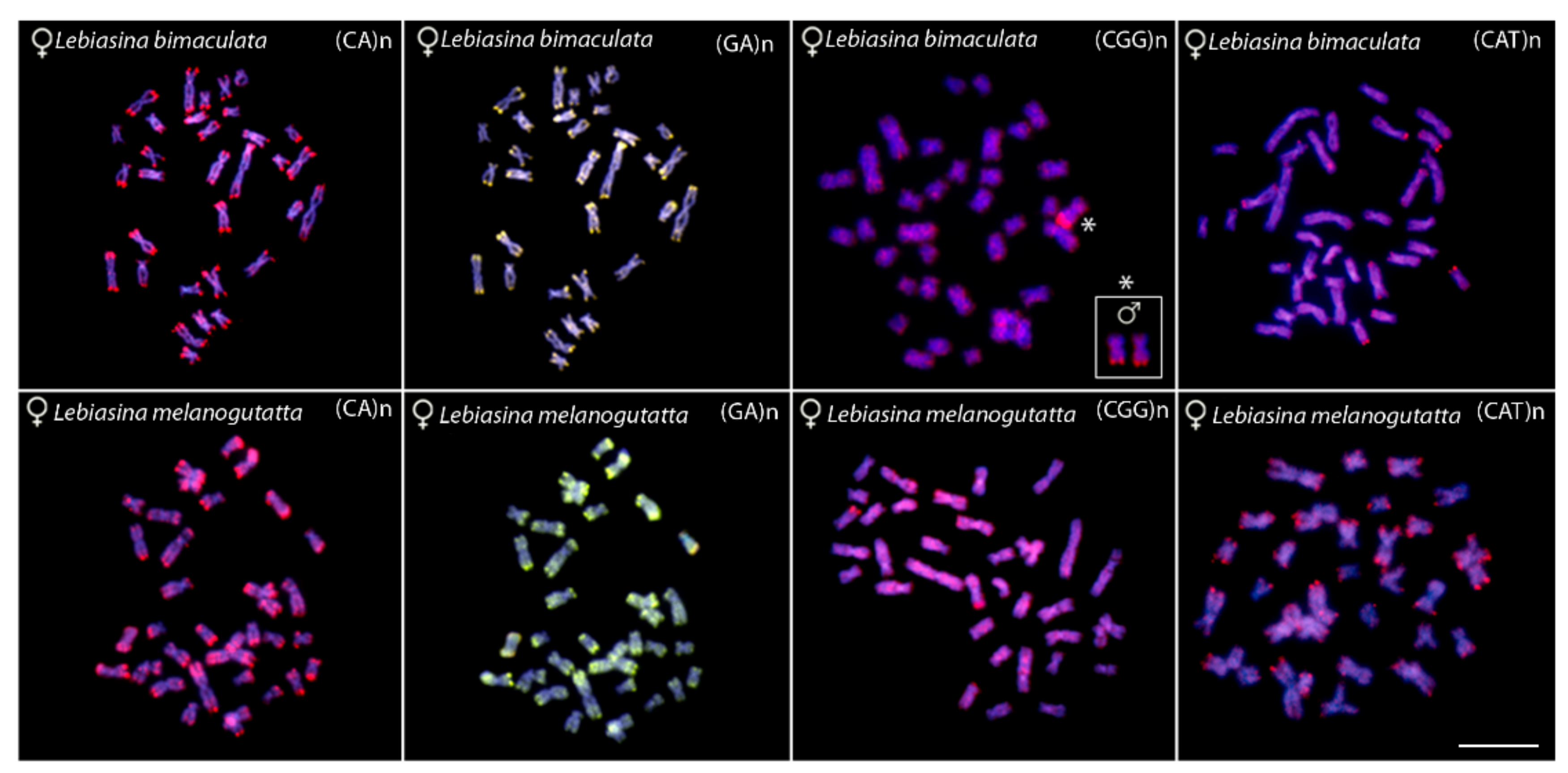
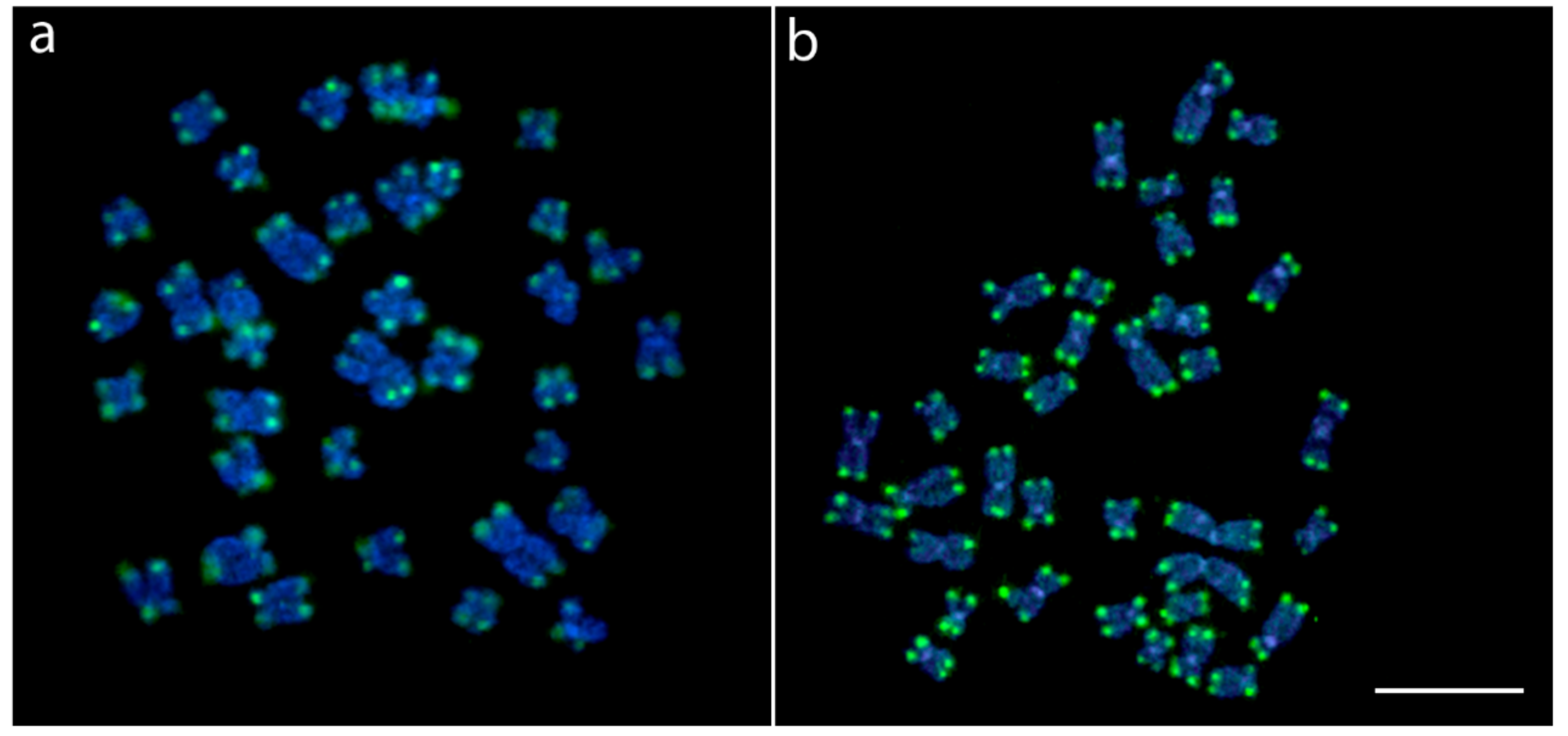
2.2. Chromosomal Mapping of Repetitive DNAs and CMA3 Banding
2.3. Comparative Genomic Hybridization (CGH)
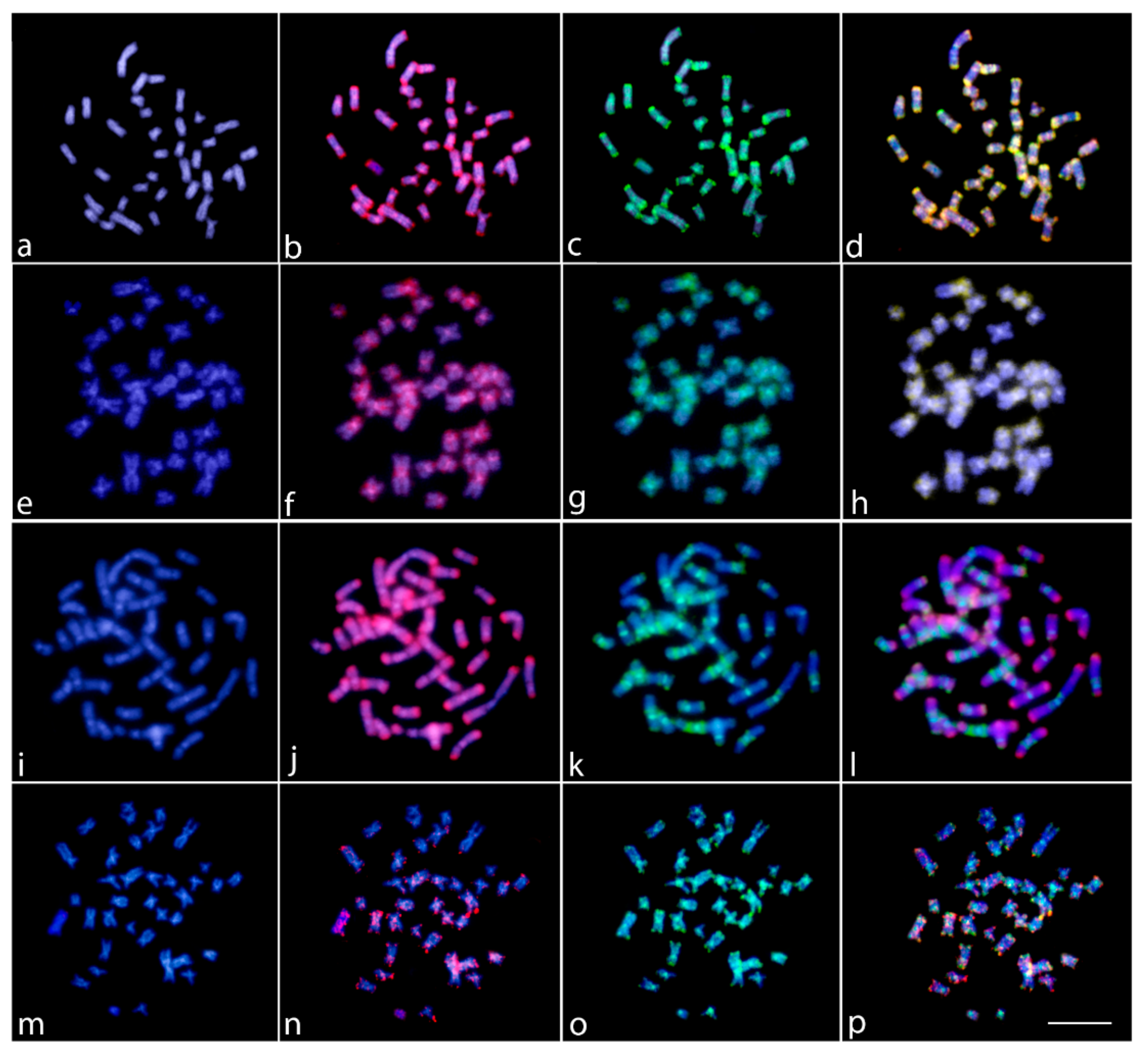
2.4. Whole Chromosome Painting (WCP)
3. Discussion
3.1. Chromosomal Features of Lebiasina Species
3.2. Heterochromatin Polymorphism and CGH: Putative Sex Chromosomes in L. bimaculata?
3.3. Relationships between Lebiasinidae and Ctenolucidae
4. Materials and Methods
4.1. Individuals
4.2. Chromosome Preparations, C- and CMA3 Bandings
4.3. Fluorescence In Situ Hybridization (FISH) for Repetitive DNA Mapping
4.4. Comparative Genome Hybridization (CGH)
4.5. Whole Chromosome Painting (WCP)
4.6. Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weitzman, M.; Weitzman, S.H. Family Lebiasinidae. In Check List of the Freshwater Fishes of South and Central America; Edipucrs: Porto Alegre, Brazil, 2003; pp. 241–250. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 9 April 2019).
- Weitzman, S.H.; Vari, R.P. Miniaturization South American Fishes; An Overview and Discussion. Proc. Biol. Soc. Washingt. 1988, 2, 444–465. [Google Scholar]
- Netto-Ferreira, A.L. Revisão taxonômica e relações interespecíficas de Lebiasinidae (Ostariophysi: Characiformes: Lebiasinidae). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2010. [Google Scholar]
- Oyakawa, O.T. Relações filogenéticas das famílias Pyrrhulinidae, Lebiasinidae e Erythrinidae (Osteichthyes: Characiformes). Ph.D. Thesis, Universidade de Sao Paulo, Sao Paulo, Brazil, 1998. [Google Scholar]
- Buckup, P.A. Relationships of the Characidiinae and phylogeny of characiform fishes (Teleostei: Ostariophysi). In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Ed.; EdiPUCRS: Porto Alegre, Brazil, 1998; pp. 123–144. [Google Scholar]
- De Pinna, M.; Zuanon, J.; Rapp Py-Daniel, L.; Petry, P. A new family of neotropical freshwater fishes from deep fossorial amazonian habitat, with a reappraisal of morphological characiform phylogeny (Teleostei: Ostariophysi). Zool. J. Linn. Soc. 2018, 182, 76–106. [Google Scholar] [CrossRef]
- Oliveira, C.; Avelino, G.S.; Abe, K.T.; Mariguela, T.C.; Benine, R.C.; Ortí, G.; Vari, R.P.; Corrêa E Castro, R.M. Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling. BMC Evol. Biol. 2011, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Arcila, D.; Ortí, G.; Vari, R.; Armbruster, J.W.; Stiassny, M.L.J.; Ko, K.D.; Sabaj, M.H.; Lundberg, J.; Revell, L.J.; Betancur, R.R. Genome-wide interrogation advances resolution of recalcitrant groups in the tree of life. Nat. Ecol. Evol. 2017, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Betancur, R.-R.; Arcila, D.; Vari, R.P.; Hughes, L.C.; Oliveira, C.; Sabaj, M.H.; Ortí, G. Phylogenomic incongruence, hypothesis testing, and taxonomic sampling: The monophyly of characiform fishes*. Evolution 2019, 73, 329–345. [Google Scholar] [CrossRef] [PubMed]
- De Bello Cioffi, M.; Moreira-Filho, O.; Ráb, P.; Sember, A.; Molina, W.F.; Bertollo, L.A.C. Conventional Cytogenetic Approaches—Useful and Indispensable Tools in Discovering Fish Biodiversity. Curr. Genet. Med. Rep. 2018, 6, 176–186. [Google Scholar] [CrossRef]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Business Media: Tokyo, Japan, 2011. [Google Scholar]
- Netto-Ferreira, A.L.; Marinho, M.M.F. New species of Pyrrhulina (Ostariophysi: Characiformes: Lebiasinidae) from the brazilian shield, with comments on a putative monophyletic group of species in the genus. Zootaxa 2013, 3664, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Marinho, M.M.F.; Menezes, N.A. Taxonomic review of Copella (Characiformes: Lebiasinidae) with an identification key for the species. PLoS ONE 2017, 12, e0183069. [Google Scholar] [CrossRef]
- Schell, J.J. Fish Chromosomes and Their Evolution; Internal Report of Danmarks Akvarium: Charlottenlund, Denmark, 1973. [Google Scholar]
- Arefjev, V.A. Problems of karyotypic variability in the family Characidae (Pisces, Characiformes) with the description of somatic karyotypes for six species of tetras. Caryologia 1990, 43, 305–319. [Google Scholar] [CrossRef]
- Oliveira, C.; Andreata, A.A.; Almeida-Toledo, L.F.; Toledo-Filho, S.A. Karyotype and nucleolus organizer regions of Pyrrhulina cf. australis (Pisces, Characiformes, Lebiasinidae). Rev. Bras. Genet. 1991, 14, 685–690. [Google Scholar]
- Oliveira, M.I.B.; Sanguino, E.C.B.; Falcão, J.N. Estudos citogenéticos em Pyrrhulina sp. (Teleostei, Characiformes, Lebiasinidae) IV. Simp. Citogenet. Evol. E Aplic. De Peixes Neotropicais: 13. Rio de Janeiro, RJ, Brasil, 1992 (abstract).
- Moraes, R.L.R.; Bertollo, L.A.C.; Marinho, M.M.F.; Yano, C.F.; Hatanaka, T.; Barby, F.F.; Troy, W.P.; de Bello Cioffi, M. Evolutionary Relationships and Cytotaxonomy Considerations in the Genus Pyrrhulina (Characiformes, Lebiasinidae). Zebrafish 2017, 14, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Moraes, R.L.R.; Sember, A.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Hatanaka, T.; Marinho, M.M.F.; Liehr, T.; Al-Rikabi, A.B.H.; Feldberg, E.; et al. Evolutionary trends and sex chromosome evolution in small-sized fish species of the genus Pyrrhulina (Characiformes, Lebiasinidae). Front. Genet. 2019. submmited. [Google Scholar]
- Barby, F.F.; Bertollo, L.A.C.; de Oliveira, E.A.; Yano, C.F.; Hatanaka, T.; Ráb, P.; Sember, A.; Ezaz, T.; Artoni, R.F.; Liehr, T.; et al. Emerging patterns of genome organization in Notopteridae species (Teleostei, Osteoglossiformes) as revealed by Zoo-FISH and Comparative Genomic Hybridization (CGH). Sci. Rep. 2019, 9, 1112. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.C.; de Oliveira, E.A.; Bertollo, L.A.C.; Yano, C.F.; Oliveira, C.; Decru, E.; Jegede, O.I.; Hatanaka, T.; Liehr, T.; Al-Rikabi, A.B.H.; et al. First chromosomal analysis in Hepsetidae (Actinopterygii, Characiformes): Insights into relationship between African and Neotropical fish groups. Front. Genet. 2017, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, N.L.; Al-Rikabi, A.B.H.; Bertollo, L.A.C.; Ezaz, T.; Yano, C.F.; de Oliveira, E.A.; Hatanaka, T.; de Bello Cioffi, M. Early Stages of XY Sex Chromosomes Differentiation in the Fish Hoplias malabaricus (Characiformes, Erythrinidae) Revealed by DNA Repeats Accumulation. Curr. Genomics 2017, 19, 216–226. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.A.; Bertollo, L.A.C.; Rab, P.; Ezaz, T.; Yano, C.F.; Hatanaka, T.; Jegede, O.I.; Tanomtong, A.; Liehr, T.; Sember, A.; et al. Cytogenetics, genomics and biodiversity of the South American and African Arapaimidae fish family (Teleostei, Osteoglossiformes). PLoS ONE 2019, 14, 1–19. [Google Scholar] [CrossRef]
- Sember, A.; Bertollo, L.A.C.; Ráb, P.; Yano, C.F.; Hatanaka, T.; de Oliveira, E.A.; de Bello Cioffi, M. Sex Chromosome Evolution and Genomic Divergence in the Fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Vari, R.P.; Malabarba, L.R. Neotropical Ichthyology: An Overview. In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Ed.; EdiPUCRS: Porto Alegre, Brazil, 1998; pp. 1–11. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilsoni, M.V.H. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- De Souza E Sousa, J.F.; Viana, P.F.; Bertollo, L.A.C.; Cioffi, M.B.; Feldberg, E. Evolutionary Relationships among Boulengerella Species (Ctenoluciidae, Characiformes): Genomic Organization of Repetitive DNAs and Highly Conserved Karyotypes. Cytogenet. Genome Res. 2017, 152, 194–203. [Google Scholar] [CrossRef]
- Nirchio, M.; Rossi, A.R.; Foresti, F.; Oliveira, C. Chromosome evolution in fishes: a new challenging proposal from Neotropical species. Neotrop. Ichthyol. 2014, 12, 761–770. [Google Scholar] [CrossRef]
- Jacobina, U.P.; Martinez, P.A.; de Bello Cioffi, M.; Garcia, J.; Bertollo, L.A.C.; Molina, W.F. Morphological and karyotypic differentiation in Caranx lugubris (Perciformes: Carangidae) in the St. Peter and St. Paul Archipelago, mid-Atlantic Ridge. Helgol. Mar. Res. 2014, 68, 17. [Google Scholar] [CrossRef]
- White, M.J.D. Modes of Speciation; WH Freeman: San Francisco, CA, USA, 1978. [Google Scholar]
- Lowry, D.B.; Willis, J.H. A widespread chromosomal inversion polymorphism contributes to a major life-history transition, local adaptation, and reproductive isolation. PLoS Biol. 2010, 8, e1000500. [Google Scholar] [CrossRef] [PubMed]
- Jay, P.; Whibley, A.; Frézal, L.; Rodríguez de Cara, M.Á.; Nowell, R.W.; Mallet, J.; Dasmahapatra, K.K.; Joron, M. Supergene Evolution Triggered by the Introgression of a Chromosomal Inversion. Curr. Biol. 2018, 28, 1839–1845. [Google Scholar] [CrossRef] [PubMed]
- Mérot, C.; Berdan, E.L.; Babin, C.; Normandeau, E.; Wellenreuther, M.; Bernatchez, L. Intercontinental karyotype-environment parallelism supports a role for a chromosomal inversion in local adaptation in a seaweed fly. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180519. [Google Scholar] [CrossRef] [PubMed]
- Supiwong, W.; Pinthong, K.; Seetapan, K.; Saenjundaeng, P.; Bertollo, L.A.C.; de Oliveira, E.A.; Yano, C.F.; Liehr, T.; Phimphan, S.; Tanomtong, A.; et al. Karyotype diversity and evolutionary trends in the Asian swamp eel Monopterus albus (Synbranchiformes, Synbranchidae): A case of chromosomal speciation? BMC Evol. Biol. 2019, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Arlt, M.F.; Durkin, S.G.; Ragland, R.L.; Glover, T.W. Common fragile sites as targets for chromosome rearrangements. DNA Repair 2006, 5, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Dedkova, O.S.; Gay, G.; Pukhalskyi, V.A.; Zelenin, A.V.; Bernard, S.; Bernard, M. Chromosomal rearrangements in wheat: Their types and distribution. Genome 2007, 50, 907–926. [Google Scholar] [CrossRef] [PubMed]
- López-Flores, I.; Garrido-Ramos, M.A. The repetitive DNA content of eukaryotic genomes. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; Karger Publishers: Basel, Switzerland, 2012; Volume 7, pp. 1–28. [Google Scholar]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [CrossRef]
- Raskina, O.; Barber, J.C.; Nevo, E.; Belyayev, A. Repetitive DNA and chromosomal rearrangements: Speciation related events in plant genomes. Cytogenet. Genome Res. 2008, 120, 351–357. [Google Scholar] [CrossRef]
- Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R. Fragile sites, dysfunctional telomere and chromosome fusions: What is 5S rDNA role? Gene 2017, 608, 20–27. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2017, 127, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Majtánová, Z.; Arias-Rodriguez, L.; Mořkovský, L.; Kořínková, T.; Cavin, L.; Pokorná, M.J.; Doležálková, M.; Flajšhans, M.; Normandeau, E.; et al. Genome Compositional Organization in Gars Shows More Similarities to Mammals than to Other Ray-Finned Fish. J. Exp. Zool. B Mol. Dev. Evol. 2016, 328, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; Karger: Basel, Switzerland, 2012; pp. 197–221. ISBN 9783318021509. [Google Scholar]
- Kobayashi, T. DNA replication, recombination, and repair: Molecular mechanisms and pathology. In DNA Replication, Recombination, and Repair: Molecular Mechanisms and Pathology; Hanaoka, F., Sugasawa, K., Eds.; Springer: Tokyo, Japan, 2016; pp. 235–247. ISBN 9784431558736. [Google Scholar]
- Zimmer, E.A.; Martins, S.L.; Beverly, S.M.; Kan, Y.W.; Wilson, A.C. Rapid duplication and loss of genes coding for the alpha chains of hemoglobin. Proc. Natl. Acad. Sci. USA 1980, 77, 2158–2162. [Google Scholar] [CrossRef]
- Dover, G.A. Molecular drive: A cohesive model of species evolution. Nature 1982, 199, 111–117. [Google Scholar] [CrossRef]
- Roy, V.; Monti-Dedieu, L.; Chaminade, N.; Siljak-Yakovlev, S.; Aulard, S.; Lemeunier, F.; Montchamp-Moreau, C. Evolution of the chromosomal location of rDNA genes in two Drosophila species subgroups: Ananassae and melanogaster. Heredity 2005, 94, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, B.; Huang, W.; Wang, Y.; Zhou, J. Bacterial community structure in simultaneous nitrification, denitrification and organic matter removal process treating saline mustard tuber wastewater as revealed by 16S rRNA sequencing. Bioresour. Technol. 2017, 228, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Khajavi, M.; Connolly, A.M.; Towne, C.F.; Batish, S.D.; Lupski, J.R. The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nat. Genet. 2009, 41, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef]
- Gold, J.R.; Karel, W.J.; Strand, M.R. Chromosome formulae of North American fishes. Progress. Fish Culturist 1980, 42, 10–23. [Google Scholar] [CrossRef]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Calcagnotto, D.; Schaefer, S.A.; DeSalle, R. Relationships among characiform fishes inferred from analysis of nuclear and mitochondrial gene sequences. Mol. Phylogenet. Evol. 2005, 36, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.S. (Instituto Nacional de Pesquisas da Amazônia—INPA, Manaus, Brazil). Personal communication, 2019.
- Bertollo, L.A.C.; Cioffi, M.B.; Moreira-Filho, O. Direct chromosome preparation from Freshwater Teleost Fishes. In Fish cytogenetic techniques (Chondrichthyans and Teleosts); Ozouf-Costaz, C., Pisano, E., Foresti, F., Almeida Toledo, L.F., Eds.; CRC Press: Enfield, CT, USA, 2015; pp. 21–26. [Google Scholar]
- Schmid, M. Chromosome banding in Amphibia. IV. Differentiation of GC-and AT-rich chromosome regions in Anura. Chromosoma 1980, 77, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Martins, C.; Ferreira, I.A.; Oliveira, C.; Foresti, F.; Galetti, P.M. A tandemly repetitive centromeric DNA sequence of the fish Hoplias malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA. Genetica 2006, 127, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation in the Y chromosome of Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning, A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zwick, M.S.; Hanson, R.E.; Mcknight, T.D.; Islam-Faridi, M.H.; Stelly, D.M.; Wing, R.A.; Price, H.J. A rapid procedure for the isolation of C 0 t-1 DNA from plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef]
- Symonová, R.; Flajšhans, M.; Sember, A.; Havelka, M.; Gela, D.; Kořínková, T.; Rodina, M.; Rábová, M.; Ráb, P.; Flajšhans, M.; et al. Molecular cytogenetics in artificial hybrid and highly polyploid sturgeons: An evolutionary story narrated by repetitive sequences. Cytogenet. Genome Res. 2013, 141, 153–162. [Google Scholar] [CrossRef]
- Yang, F.; Trifonov, V.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of paint probes by flow-sorted and microdissected chromosomes. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Liehr, T., Ed.; Springer-Verlag: Berlin, Germany, 2009. [Google Scholar]
- Yano, C.F.; Bertollo, L.A.C.; Ezaz, T.; Trifonov, V.; Sember, A.; Liehr, T.; Cioffi, M.B. Highly conserved Z and molecularly diverged W chromosomes in the fish genus Triportheus (Characiformes, Triportheidae). Heredity 2017, 118, 276. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
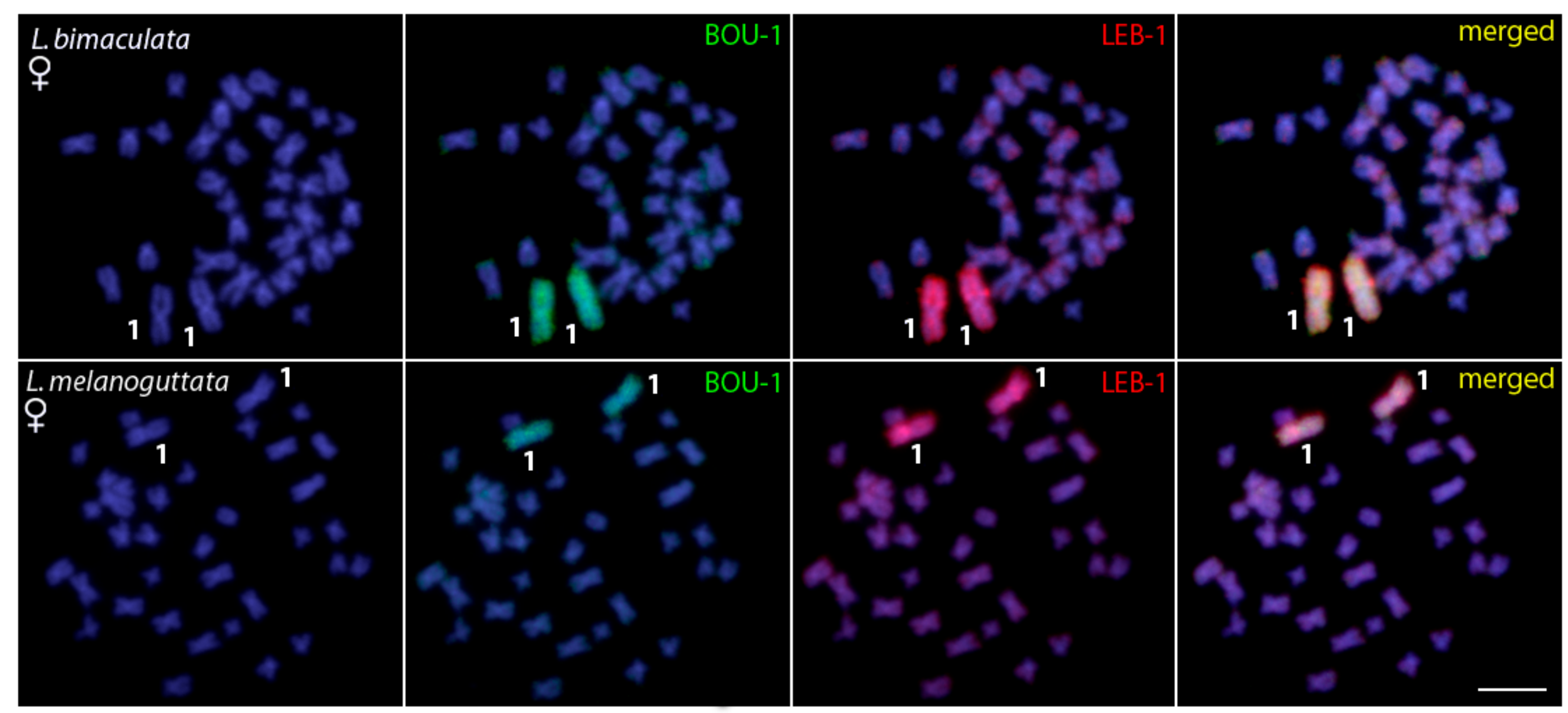{kind=link}
{kind=link}
| Species | 2n (Sex) | Karyotype | Reference |
|---|---|---|---|
| Copeina | |||
| C. guttata | 42 (?) | - | [15] |
| Copella | |||
| C. arnoldi | 44 (?) | - | [15] |
| C. nattereri | 36 (?) | - | [15] |
| Copella sp. | 26 (?) | - | [15] |
| Copella sp. | 24 (?) | - | [15] |
| Nannostomus | |||
| N. beckfordi (A) | 42 ♂ | 2m + 40a | [16] |
| N. beckfordi (B) | 44 (?) | - | [15] |
| N. beckfordi (C) | 36 (?) | - | [15] |
| N. eques (A) | 34 (?) | 34a | [16] |
| N. eques (B) | 36 (?) | - | [15] |
| N. arrisoni | 40 (?) | - | [15] |
| N. marginatus | 42 (?) | - | [15] |
| N. trifasciatus (A) | 46 (?) | - | [15] |
| N. trifasciatus (B) | 38 (?) | - | [15] |
| N. trifasciatus (C) | 30 (?) | - | [15] |
| N. trifasciatus (D) | 24 (?) | - | [15] |
| N. unifasciatus | 22 (?) | - | [15] |
| Pyrrhulina | |||
| Pyrrhulina cf. australis | 40♂♀ | 6st + 34a | [17] |
| Pyrrhulina sp. | 42 (?) | 2m + 2sm + 38st/a | [18] |
| P. australis | 40♂♀ | 4st + 36a | [19] |
| Pyrrhulina cf. australis | 40♂♀ | 4st + 36a | [19] |
| P. brevis | 42♂♀ | 2sm + 4st + 36a | [20] |
| P. semifasciata | 41♂42♀ | 1m + 4st + 36a ♂ 4st + 38a ♀ | [20] |
| Species | Locality | N |
|---|---|---|
| Lebiasina bimaculata | Arenillas river lakes—El Oro (Ecuador) (S03°30′57.204″, W80°3′44.2656″) | 04♂, 03♀ |
| Lebiasina melanoguttata | Altamira—PA (Brazil) (S08° 46′ 59,4″, W54°58′26,9″) | 10♂, 04♀ |
| Lebiasina melanoguttata | Cachoeira da Serra—PA (Brazil) (S08°58′18,7″, W54°58′18,7″) | 04♂, 18♀ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Menezes Cavalcante Sassi, F.; Aguiar de Oliveira, E.; Antonio Carlos Bertollo, L.; Nirchio, M.; Hatanaka, T.; Maria Ferreira Marinho, M.; Moreira-Filho, O.; Aroutiounian, R.; Liehr, T.; B. H. Al-Rikabi, A.; et al. Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae). Int. J. Mol. Sci. 2019, 20, 2944. https://doi.org/10.3390/ijms20122944
de Menezes Cavalcante Sassi F, Aguiar de Oliveira E, Antonio Carlos Bertollo L, Nirchio M, Hatanaka T, Maria Ferreira Marinho M, Moreira-Filho O, Aroutiounian R, Liehr T, B. H. Al-Rikabi A, et al. Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae). International Journal of Molecular Sciences. 2019; 20(12):2944. https://doi.org/10.3390/ijms20122944
Chicago/Turabian Stylede Menezes Cavalcante Sassi, Francisco, Ezequiel Aguiar de Oliveira, Luiz Antonio Carlos Bertollo, Mauro Nirchio, Terumi Hatanaka, Manoela Maria Ferreira Marinho, Orlando Moreira-Filho, Rouben Aroutiounian, Thomas Liehr, Ahmed B. H. Al-Rikabi, and et al. 2019. "Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae)" International Journal of Molecular Sciences 20, no. 12: 2944. https://doi.org/10.3390/ijms20122944
APA Stylede Menezes Cavalcante Sassi, F., Aguiar de Oliveira, E., Antonio Carlos Bertollo, L., Nirchio, M., Hatanaka, T., Maria Ferreira Marinho, M., Moreira-Filho, O., Aroutiounian, R., Liehr, T., B. H. Al-Rikabi, A., & de Bello Cioffi, M. (2019). Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae). International Journal of Molecular Sciences, 20(12), 2944. https://doi.org/10.3390/ijms20122944