Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function

 ,
, 
Abstract
1. Introduction
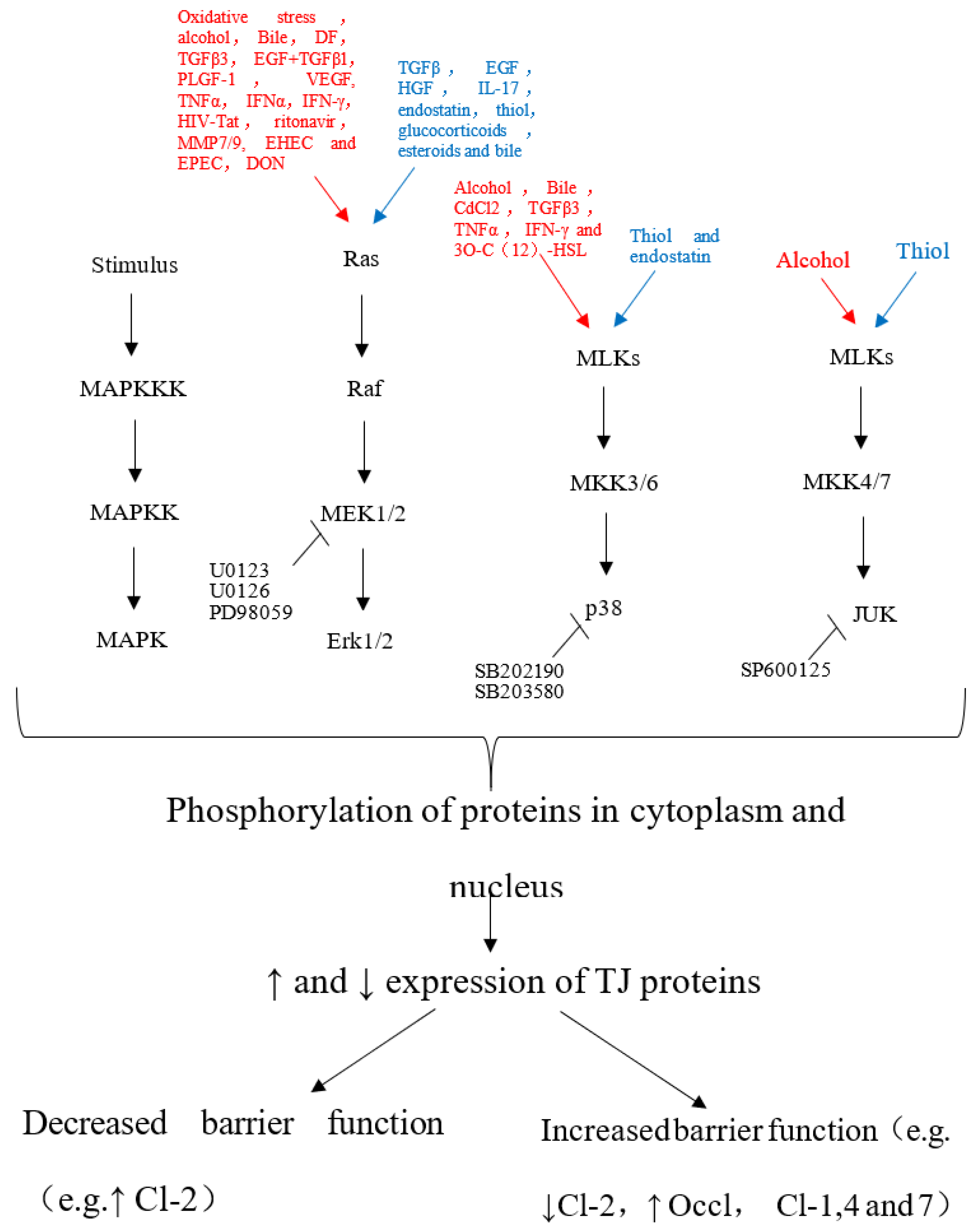
2. Effect of Mycotoxins on the Mechanical Barrier Function of the Intestinal Mucosa
3. Effect of Mycotoxins on Intestinal Chemical Barrier Function
4. Effect of Mycotoxins on Intestinal Mucosal Immune Barrier Function
5. Effect of Mycotoxins on Intestinal Mucosal Biological Barrier Function
6. Summary
Funding
Acknowledgments
Conflicts of Interest
References
- Maresca, M.; Yahi, N.; Younès-Sakr, L.; Boyron, M.; Caporiccio, B.; Fantini, J. Both direct and indirect effects account for the pro-inflammatory activity of enteropathogenic mycotoxins on the human intestinal epithelium: Stimulation of interleukin-8 secretion, potentiation of interleukin-1β effect and increase in the transepithelial passage of commensal bacteria. Toxicol. Appl. Pharmacol. 2008, 228, 84–92. [Google Scholar] [PubMed]
- Biomin. Report on the Detection of Mycotoxins in Raw Materials and Feeds in 2018; Biomin: Inzersdorf-Getzersdorf, Austria, 2019. [Google Scholar]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in finish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Asi, M.R.; Ariňo, A. Aflatoxin M1 contamination in cow and buffalo milk samples from the North West Frontier Province (NWFP) and Punjab provinces of Pakistan. Food Addit. Contam. Part B Surveill. 2011, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Ruiz, M.J.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Font, G.; Mañes, J. Isolation and purification of enniatins A, A1, B, B1, produced by Fusarium tricinctum in solid culture, and cytotoxicity effects on Caco-2 cell. Toxicon 2010, 56, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Malachova, A.; Dzuman, Z.; Veprikova, Z.; Vaclavikova, M.; Zachariasova, M.; Hajslova, J. Deoxynivalenol, deoxynivalenol-3-glucoside, and enniatins: The major mycotoxins found in cereal-based products on the Czech market. J. Agric. Food Chem. 2011, 59, 12990–12997. [Google Scholar] [CrossRef]
- Tatay, E.; Meca, G.; Font, G.; Ruiz, M.J. Interactive effects of zearalenone and its metabolites on cytotoxicity and metabolization in ovarian CHO-K1 cells. Toxicol. In Vitro 2014, 28, 95–103. [Google Scholar] [CrossRef]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.-M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Animal Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef]
- Wan, L.; Turner, P.; Allen, K.; El-Nezami, H. Modulation of mucin mRNA (MUC5AC and MUC5B) expression and protein production and secretion in Caco-2/HT29-MTX Co-cultures following exposure to individual and combined fusarium mycotoxins. Toxicol. Sci. 2014, 139, 83–98. [Google Scholar] [CrossRef]
- Constantinescu, C.S.; Chou, I.-J. Intestinal Bacterial Antigens, Toxin-Induced Pathogenesis and Immune Cross-Reactivity in Neuromyelitis Optica and Multiple Sclerosis. In Neuro-Immuno-Gastroenterology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 227–236. [Google Scholar]
- Capaldo, C.T.; Farkas, A.E.; Nusrat, A. Epithelial adhesive junctions. F1000prime Rep. 2014, 6, 1. [Google Scholar] [CrossRef]
- Manasa, J.S. Role of the normal gut microbiota. World J. Gastro. Oncol. 2015, 21, 8787–8803. [Google Scholar]
- Gill, N.; Wlodarska, M.; Finlay, B.B. Roadblocks in the gut: Barriers to enteric infection. Cell Microbiol. 2011, 13, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Potten, C.S. Gut instincts: Thoughts on intestinal epithelial stem cells. J. Clin. Investig. 2000, 105, 1493–1499. [Google Scholar]
- Awad, W.A.; JBöhm Razzazi-Fazeli, E.; Ghareeb, K.; Zentek, J. Effect of addition of a probiotic microorganism to broiler diets contaminated with deoxynivalenol on performance and histological alterations of intestinal villi of broiler chickens. Poult. Sci. 2006, 85, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, F.; Hara-Kudo, Y.; Saito, N.; Kumagai, S.; Sugita-Konishi, Y. In vitro effect of deoxynivalenol on the differentiation of human colonic cell lines Caco-2 and t84. Mycopathologia 1998, 142, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Jiang, M.; Peng, X.; Cui, H.; Zhou, Y.; He, M.; Zuo, Z.; Ouyang, P.; Fan, J. The molecular mechanism of G2M cell cycle arrest induced by AFB1 in the jejunum. Oncotarget 2016, 7, 35592–35606. [Google Scholar] [CrossRef]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.; Anadón, A.; Martínez, M. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology 2016, 353–354, 21–33. [Google Scholar] [CrossRef]
- Gao, Y.; Li, S.; Wang, J.; Luo, C.; Zhao, S.; Zheng, N. Modulation of intestinal epithelial permeability in differentiated Caco-2 cells exposed to aflatoxin M1 and ochratoxin A individually or collectively. Toxins 2018, 10, 13. [Google Scholar] [CrossRef]
- Bracarense, A.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Osselaere, A.; Santos, R.; Hautekiet, V.; De Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol impairs hepatic and intestinal gene expression of selected oxidative stress, tight junction and inflammation proteins in broiler chickens, but addition of an adsorbing agent shifts the effects to the distal parts of the small intestine. PLoS ONE 2013, 8, e69014. [Google Scholar] [CrossRef]
- Awad, W.; Böhm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of feeding deoxynivalenol contaminated wheat on growth performance, organ weights and histological parameters of the intestine of broiler chickens. J. Animal Physiol. Animal Nutr. Berl 2006, 90, 6. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Su, Y.; Shen, D.; Dai, P.; Li, C. Short-term ingestion of deoxynivalenol in naturally contaminated feed alters piglet performance and gut hormone secretion. Animal Sci. J. 2018, 89, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Braber, S.; Gremmels, H.; Koelink, P.; Verheijden, K.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hsiao, F.; Proskura, W.; Dybus, A.; Siao, Y.; Cheng, Y. An impact of Deoxynivalenol produced by Fusarium graminearum on broiler chickens. J. Animal Physiol. Animal Nutr. Berl. 2018, 102, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Duan, Y.; Wang, F.; Guo, F.; Yan, F.; Yang, X.; Yang, X. Intestinal toxicity of deoxynivalenol is limited by supplementation with Lactobacillus plantarum JM113 and consequentially altered gut microbiota in broiler chickens. J. Animal Sci. Biotechnol. 2018, 9. [Google Scholar] [CrossRef]
- Yunus, A.; Ghareeb, K.; Abd-El-Fattah, A.; Twaruzek, M.; Böhm, J. Gross intestinal adaptations in relation to broiler performance during chronic aflatoxin exposure. Poult. Sci. 2011, 90, 1683–1689. [Google Scholar] [CrossRef]
- Kana, J.; Teguia, A.; Tchoumboue, J. Effect of dietary plant charcoal from Canarium schweinfurthii Engl. and maize cob on aflatoxin B1 toxicosis in broiler chickens. Adv. Animal Biosci. 2010, 1, 462–463. [Google Scholar] [CrossRef]
- Wang, F.; Zuo, Z.; Chen, K.; Gao, C.; Yang, Z.; Zhao, S.; Li, J.; Song, H.; Peng, X.; Fang, J.; et al. Histopathological injuries, ultrastructural changes, and depressed TLR expression in the small intestine of broiler chickens with aflatoxin B1. Toxins 2018, 10, 131. [Google Scholar] [CrossRef]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Hanuszewska, M.; Petrusewicz-Kosińska, M.; Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The effects of deoxynivalenol and zearalenone on the pig large intestine. A light and electron microscopy study. Toxins 2018, 10, 148. [Google Scholar] [CrossRef]
- Lessard, M.; Savard, C.; Deschene, K.; Lauzon, K.; Pinilla, V.; Gagnon, C.; Lapointe, J.; Guay, F.; Chorfi, Y. Impact of deoxynivalenol (DON) contaminated feed on intestinal integrity and immune response in swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef]
- Diesing, A.; Nossol, C.; Panther, P.; Walk, N.; Post, A.; Kluess, J.; Kreutzmann, P.; Dänicke, S.; Rothkötter, H.; Kahlert, S. Mycotoxin deoxynivalenol (DON) mediates biphasic cellular response in intestinal porcine epithelial cell lines IPEC-1 and IPEC-J2. Toxicol. Lett. 2011, 200, 8–18. [Google Scholar] [CrossRef]
- Pinton, P.; Braicu, C.; Nougayrede, J.; Laffitte, J.; Taranu, I.; Oswald, I. Deoxynivalenol impairs porcine intestinal barrier function and decreases the protein expression of claudin-4 through a mitogen-activated protein kinase-dependent mechanism. J. Nutr. 2010, 140, 1956. [Google Scholar] [CrossRef]
- González-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of tight junction components with signaling pathways. BBA Biomembr. 2008, 1778, 729–756. [Google Scholar] [CrossRef]
- Bensassi, F.; El Golli-Bennour, E.; Abid-Essefi, S.; Bouaziz, C.; Hajlaoui, M.; Bacha, H. Pathway of deoxynivalenol-induced apoptosis in human colon carcinoma cells. Toxicology 2009, 264, 104–109. [Google Scholar] [CrossRef]
- Taranu, I.; Braicu, C.; Marin, D.; Pistol, G.; Motiu, M.; Balacescu, L.; Beridan Neagoe, I.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef]
- Abid-Essefi, S.; Baudrimont, I.; Hassen, W.; Ouanes, Z.; Mobio, T.; Anane, R.; Creppy, E.; Bacha, H. DNA fragmentation, apoptosis and cell cycle arrest induced by zearalenone in cultured DOK, Vero and Caco-2 cells: Prevention by Vitamin E. Toxicology 2003, 192, 237–248. [Google Scholar] [CrossRef]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.; De Baere, S.; Mountzouris, K.; Su, S.; Wong, E.; De Meulenaer, B.; et al. The mycotoxins deoxynivalenol and fumonisins alter the extrinsic component of intestinal barrier in broiler chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef]
- Wan, M.; Turner, P.; Allen, K.; Nezami, H. Lactobacillus rhamnosus GG modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J. Funct. Foods 2016, 22, 34–43. [Google Scholar] [CrossRef]
- Pinton, P.; Graziani, F.; Pujol, A.; Nicoletti, C.; Paris, O.; Ernouf, P.; Di Pasquale, E.; Perrier, J.; Oswald, I.; Maresca, M. Deoxynivalenol inhibits the expression by goblet cells of intestinal mucins through a PKR and MAP kinase dependent repression of the resistin-like molecule β. Mol. Nutr. Food Res. 2015, 59, 1076–1087. [Google Scholar] [CrossRef]
- Applegate, T.; Schatzmayr, G.; Prickel, K.; Troche, C.; Jiang, Z. Effect of aflatoxin culture on intestinal function and nutrient loss in laying hens. Poult. Sci. 2009, 88, 1235. [Google Scholar] [CrossRef]
- Wan, M.; Woo, C.; Allen, K.; Turner, P.; El-Nezami, H. Modulation of porcine?—Defensins 1 and 2 upon individual and combined fusarium toxin exposure in a swine jejunal epithelial cell line. Appl. Environ. Microbiol. 2013, 79, 2225–2232. [Google Scholar] [CrossRef]
- Han, F.; Zhang, H.; Xia, X.; Xiong, H.; Song, D.; Zong, X.; Wang, Y. Porcine beta-defensin 2 attenuates inflammation and mucosal lesions in dextran sodium sulfate-induced colitis. J. Immunol. 2015, 194, 1882–1893. [Google Scholar] [CrossRef]
- Xiao, H.; Tan, B.; Wu, M.; Yin, Y.; Li, T.; Yuan, D.; Li, L. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: II. Intestinal morphology and function. J. Animal Sci. 2013, 91, 4750–4756. [Google Scholar] [CrossRef]
- Yi, H. Effect of antimicrobial peptide CWA on intestinal inflammation and intestinal barrier function in weaned piglets and its mechanism. Sci. Rep. 2016, 6, 25679. [Google Scholar] [CrossRef]
- Jiang, M.; Fang, J.; Peng, X.; Cui, H.; Yu, Z. Effect of aflatoxin B1 on IgA(+) cell number and immunoglobulin mRNA expression in the intestine of broilers. Immunopharmacol. Immunotoxicol. 2015, 37, 450–457. [Google Scholar] [CrossRef]
- Vignal, C.; Djouina, M.; Pichavant, M.; Caboche, S.; Waxin, C.; Beury, D.; Hot, D.; Gower-Rousseau, C.; Body-Malapel, M. Chronic ingestion of deoxynivalenol at human dietary levels impairs intestinal homeostasis and gut microbiota in mice. Arch. Toxicol. 2018, 92, 2327–2338. [Google Scholar] [CrossRef]
- Pinton, P.; Accensi, F.; Beauchamp, E.; Cossalter, A.; Callu, P.; Grosjean, F.; Oswald, I. Ingestion of deoxynivalenol (DON) contaminated feed alters the pig vaccinal immune responses. Toxicol. Lett. 2008, 177, 215–222. [Google Scholar] [CrossRef]
- Becker, C.; Reiter, M.; Pfaffl, M.; Meyer, H.; Bauer, J.; Meyer, K. Expression of immune relevant genes in pigs under the influence of low doses of deoxynivalenol (DON). Mycotoxin Res. 2011, 27, 287–293. [Google Scholar] [CrossRef]
- Moon, Y.; Yang, H.; Lee, S. Modulation of early growth response gene 1 and interleukin-8 expression by ribotoxin deoxynivalenol (vomitoxin) via ERK1/2 in human epithelial intestine 407 cells. Biochem. Biophys. Res. Commun. 2007, 362, 256–262. [Google Scholar] [CrossRef]
- Walle, J.; Sergent, T.; Piront, N.; Toussaint, O.; Schneider, Y.; Larondelle, Y. Deoxynivalenol affects in vitro intestinal epithelial cell barrier integrity through inhibition of protein synthesis. Toxicol. Appl. Pharm. 2010, 245, 291–298. [Google Scholar] [CrossRef]
- Walle, J.; Romier, B.; Larondelle, Y.; Schneider, Y. Influence of deoxynivalenol on NF-kappa B activation and IL-8 secretion in human intestinal Caco-2 cells. Toxicol. Lett. 2008, 177, 205–214. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, Z.; Yang, W.; Jiang, S.; Zhang, G.; Ji, F.; Chen, J. Effects of zearalenone on performance and internal organs development of piglets. Food Feed Ind. 2008, 10, 37–38. [Google Scholar]
- He, Y.; Fang, J.; Peng, X.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W.; Shu, G.; et al. Effects of sodium selenite on aflatoxin B1-induced decrease of ileal IgA+ cell numbers and immunoglobulin contents in broilers. Biol. Trace Elem. Res. 2014, 160, 49–55. [Google Scholar] [CrossRef]
- Cano, P.; Seeboth, J.; François, M.; Juliette, C.; Roberta, A.; Isabelle, P.; Laurence, G. Deoxynivalenol as a new factor in the persistence of intestinal inflammatory diseases: An emerging hypothesis through possible modulation of Th17-mediated response. PLoS ONE 2013, 8, e53647. [Google Scholar] [CrossRef]
- Kadota, T.; Furusawa, H.; Hirano, S.; Tajima, O.; Kamata, Y.; Sugita-Konishi, Y. Comparative study of deoxynivalenol, 3-acetyldeoxynivalenol, and 15-acetyldeoxynivalenol on intestinal transport and IL-8 secretion in the human cell line Caco-2. Toxicol. In Vitro 2013, 27, 1888–1895. [Google Scholar] [CrossRef]
- Pesaka, J.J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam. Part A 2008, 25, 1128–1140. [Google Scholar]
- Ghareeb, K.; Awad, W.; Böhm, J.; Zebeli, Q. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–337. [Google Scholar] [CrossRef]
- Awad, W.; Zentek, J. The feed contaminant deoxynivalenol affects the intestinal barrier permeability through inhibition of protein synthesis. Arch. Toxicol. 2015, 89, 961–965. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Glenn, T.; Wang, J. Aflatoxin B1 induced compositional changes in gut microbial communities of male F344 rats. Toxicol. Sci. 2015, 150, 54–63. [Google Scholar] [CrossRef]
- Guo, M.; Huang, K.; Chen, S.; Qi, X.; He, X.; Cheng, W.; Luo, Y.; Xia, K.; Xu, W. Combination of metagenomics and culture-based methods to study the interaction between ochratoxin A and gut microbiota. Toxicol. Sci. 2014, 141, 314–323. [Google Scholar] [CrossRef]
- Piotrowska, M.; Śliżewska, K.; Nowak, K.; Zielonka, Ł.; Żakowska, Z.; Gajęcka, M.; Gajęcki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef]
- Lucke, A.; Böhm, J.; Zebeli, Q.; Metzler-Zebeli, B. Dietary deoxynivalenol contamination and oral lipopolysaccharide challenge alters the cecal microbiota of broiler chickens. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Saint-Cyr, M.; Perrin-Guyomard, A.; Houée, P.; Rolland, J.; Laurentie, M. Evaluation of an oral subchronic exposure of deoxynivalenol on the composition of human gut microbiota in a model of human microbiota-associated rats. PLoS ONE 2013, 8, e80578. [Google Scholar] [CrossRef]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.; et al. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef]
- Payros, D.; Dobrindt, U.; Martin, P.; Secher, T.; Bracarense, A.; Boury, M.; Laffitte, J.; Pinton, P.; Oswald, E.; Oswald, I. The food contaminant deoxynivalenol exacerbates the genotoxicity of gut microbiota. mBio 2017, 8. [Google Scholar] [CrossRef]
- Cani, P.; Everard, A. Talking microbes: When gut bacteria interact with diet and host organs. Mol. Nutr. Food Res. 2016, 60, 58–66. [Google Scholar] [CrossRef]
- Claus, S.; Guillou, H.; Ellero-simatos, S. The gut microbiota: A major player in the toxicity of environmental pollutants? Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef]
- Goodrich, J.; Davenport, E.; Waters, J.; Clark, A.; Ley, R. Cross-species comparisons of host genetic associations with the microbiome. Science 2016, 352, 532–535. [Google Scholar] [CrossRef]


{kind=link}
| Mycotoxins | In Vitro/In Vivo | Species | Dose | Reference |
|---|---|---|---|---|
| DON | In vivo | broilers | 10 mg/kg | [17] |
| pigs | 2.29 mg/kg | [18] | ||
| In vitro | Caco-2 | 50, 100, 200 ng/mL | [19] | |
| the jejunal explant | 10, 30 μM | [20] | ||
| AFB1 | In vivo | broilers | 0.6 mg/kg | [21] |
| In vitro | Caco-2 | 1–100 μM | [22] | |
| AFM1 | In vitro | Caco-2 | 0.12 M, 12 M | [23] |
| OTA | 1–100 μM | [22] | ||
| FB1 | 1–100 μM | [22] | ||
| T2 | 1–100 μM | [22] | ||
| FB1 | In vivo | pigs | 6 mg/kg | [24] |
| Mycotoxin | Species | Toxicity | References |
|---|---|---|---|
| DON | Broilers, rats, pigs | The villi of the duodenum and jejunum become shorter and thinner, flattened and fused, and apical necrosis | [17,18,25,26,27,28,29,30] |
| Caco-2 | (1) Decompose the Caco-2 cell monolayer; (2) Decrease the epithelial resistance; (3) Increase the permeability of the labeling molecule, such as Lucifer Yellow and FITC-labeled dextran; (4) Reduce the TEER. | [27] | |
| the jejunal explant | (1) The villi are flattened; (2) The adhesion and the lamina propria are weakened; (3) The epithelial cells are shed; (4) The scrotum is dilated. | [20] | |
| AFB1 | broilers | (1) Reduce jejunal villus density and intestinal absorption area; (2) Reduce jejunal villus height and villus height/crypt rate; (3) Reduce the goblet cell number; (4) Reduce the microvilli in intestinal absorption cells; (5) Mitochondrial vacuolization; (6) Mitochondrial ridges disappear; (7) Connection complex and terminal network disappear. | [21,31,32,33] |
| AFM1+OTA | Caco-2 | Reduce transepithelial resistance | [23] |
| DON+ZEA | pigs | Reduce the number of goblet cells in the cecum and descending colon | [34] |
| DON+FB | pigs | (1) Villus atrophy and fusion; (2) Reduce the jejunum villus height and cell proliferation; (3) Decrease in goblet cells and lymphocytes. | [24] |
| Mycotoxins | In Vitro/In Vivo | Species | Dose | Reference |
|---|---|---|---|---|
| FBs | In vivo | broilers | 25.4 mg (FB1+FB2)/kg feed | [42] |
| DON | In vivo | Broilers | 4.6 mg/kg feed | [42] |
| mouse | 12 mg/kg | [43] | ||
| In vitro | human goblet cells (HT29-16E)/porcine small intestine explants | 1, 10, 100 μM | [44] | |
| ZEA | In vivo | mouse | 0.5 mg/kg | [43] |
| Mycotoxins | In Vitro/In Vivo | Species | Dose | Reference |
|---|---|---|---|---|
| AFB1 | In vivo | broilers | 0.6 mg/kg | [50] |
| DON | In vivo | mouse | 10 μg/kg BW | [51] |
| pigs | 3 mg/kg | [24] | ||
| 2.2–2.5 mg DON/kg feed | [52] | |||
| 1.2–2.0 mg/kg | [53] | |||
| In vitro | human epithelial cells | 0, 25, 500, 1000 bg/mL | [54] | |
| Cac0-2 | 50, 500, 5000 ng/mL | [55] | ||
| 250–1000 ng/mL | [56] | |||
| FB1 | In vivo | pigs | 6 mg/kg | [24] |
| ZEA | In vivo | pigs | 1, 2, 3 mg/kg | [57] |
| Mycotoxin | Species | Toxicity | References |
|---|---|---|---|
| AFB1 | broilers | (1) Reduce the humoral immune function of the ileal mucosa in broilers; (2) Reduce the mRNA expression levels of immunoglobulins, such as IgA, pIgR, IgM, and IgG. | [50,58] |
| DON/FB1 | pigs | Reduce the IgG content in serum and lymphocyte proliferation in piglets | [24] |
| ZEA | pigs | (1) Increase the levels of IgG and IgM in serum; (2) Decrease proportion of neutrophils indicates. | [57] |
| DON | human epithelial intestine 407 cells | Increase the level of IL-8 in human intestinal epithelial cells | [54] |
| Caco-2 | (1) A dose-dependent increase in IL-8 was shown in Caco-2 cells; (2) Increase the expression of IL-1-α, IL-1β, and TNF-α. | [55,56,59,60] | |
| pigs | (1) DON do not alter the expression of TGF-β, IFN-γ, IL-4, and IL-6 mRNA in the pig ileum following a short, low-dose regimen; (2) Reduce the expression of IL-1β and IL-8 in the pig ileum following a longer-term, low dose regimen. (3) Short-term, high-dose exposure to DON significantly upregulate TNF-α, IL-1β, IFN-γ, IL-6, and IL-10 levels in the ileum and jejunum of pigs | [24,52,53] | |
| mouse | (1) Induce the recruitment of regulatory B cells; (2) Activation of regulatory T cells and dendritic cells in the mesenteric lymph nodes. | [51] |
| Mycotoxins | Species | Dose | Reference |
| AFB1 | rats | 5, 25, 75 μg/kg BW | [64] |
| OTA | rats | 0, 70, 210 μg/kg BW | [65] |
| ZEA | pigs | 40 μg/kg BW | [66] |
| DON | pigs | 12 μg/kg BW | [66] |
| broilers | 0, 2.5, 5, 10 mg/kg feed | [67] | |
| rats | 100 μg/kg BW | [68] | |
| FBs | pigs | 0.41 mg/kg FB1 + 0.17 mg/kg FB2 | [69] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. https://doi.org/10.3390/ijms20112777
Ren Z, Guo C, Yu S, Zhu L, Wang Y, Hu H, Deng J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. International Journal of Molecular Sciences. 2019; 20(11):2777. https://doi.org/10.3390/ijms20112777
Chicago/Turabian StyleRen, Zhihua, Chaoyue Guo, Shumin Yu, Ling Zhu, Ya Wang, Hui Hu, and Junliang Deng. 2019. "Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function" International Journal of Molecular Sciences 20, no. 11: 2777. https://doi.org/10.3390/ijms20112777
APA StyleRen, Z., Guo, C., Yu, S., Zhu, L., Wang, Y., Hu, H., & Deng, J. (2019). Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. International Journal of Molecular Sciences, 20(11), 2777. https://doi.org/10.3390/ijms20112777