1. Introduction
The genus
Dendrobium in the subfamily Epidendroideae, Orchidaceae, which consists of approximately 1450 species, is one of the largest genera in Orchidaceae. Most of the species in this genus are epiphytes with high ornamental, medicinal, and commercial value and are favoured by botanists and plant enthusiasts [
1]. As one of the largest Orchidaceae genera, most (72%) of the 61
Dendrobium species that are known to be self-pollinated show self-sterility [
2]. Nearly half of orchid self-incompatibility (SI) species are from
Dendrobium. So, the high SI rate in
Dendrobium species might contribute to their high level of species diversity.
SI is a type of reproductive barrier within species which can promote outcrossing and maintain heterozygosity. SI, with multiple independent origins, exists in approximately 40% of flowering plants, based on the plant species tested [
3,
4]. SI is closely related to the formation and maintenance of the high diversity and adaptation of angiosperm species [
5,
6].
The SI response is controlled by a single highly polymorphic locus, the S-locus, in most reported species. There are at least two closely linked genes (the male and female S-determinants) located at the S-locus, and self and non-self modes are discriminated by a specific molecular interaction between these S-determinants [
7]. Based on the modes of genetic control of the pollen SI phenotype, SI can be classified as gametophytic SI (GSI) or sporophytic SI (SSI), in which the pollen SI phenotype is controlled by the haploid pollen itself (Papaveraceae, Solanaceae, Rosaceae, and Plantaginaceae) and diploid donor tissues (anther tapetum in
Brassica of the Brassicaceae) [
5], respectively. There are three types of well-studied SI systems with molecular characterization: Firstly, the self-recognition mechanism of SSI in Brassicaceae, especially in
Brassica. The male S-determinant is the S-locus protein 11 (SP11) or the S-locus cysteine-rich protein (SCR), a small secreted peptide that localizes in the pollen coat [
8,
9]. The female S-determinant is the S-locus receptor kinase (SRK), a Ser/Thr receptor kinase localized to the plasma membrane of the papilla cells of the stigma [
10]. The interaction between these two S-determinants from the same S-haplotype induces the incompatibility response in the stigma, leading to self-pollen rejection [
11,
12,
13] which belongs to the receptor and ligand interaction type. Secondly, there is the self-recognition mechanism of gametophytic self-incompatibility (GSI) in Papaveraceae, mainly in
Papaver rhoeas. The interaction between the male S-determinant (
P. rhoeas pollen S,
PrpS, pollen surface receptor) and the female S-determinant (
P. rhoeas style S,
PrsS, small protein secreted by papilla cells of the stigma) from the same S-haplotype induces the SI response and belongs to the novel “receptor”–ligand interaction type [
14]. Thirdly, the non-self-recognition mechanism of GSI in Solanaceae, Rosaceae, and Plantaginaceae, is different from the above two types. The male S-determinant is made up of multiple
S-locus F-box (
SLF) genes [
15], encoding the F-box proteins [
16], and the female S-determinant S-RNase is mainly present as a glycoprotein in the style tissue. Each SLF protein interacts specifically with one or more S-RNase proteins of other S-haplotypes. The set of SLF genes encoded by a single S-haplotype allows pollen to recognize and detoxify multiple allelic variants of non-self S-RNase [
15,
17], which is called the collaborative non-self-recognition model.
In addition to the differences between the molecular mechanisms among these three SI systems, the pollen tube morphologies of SI also show significant differences. In Solanaceae, the growth of the incompatible pollen tube is arrested by the time it reaches approximately one-third of the way through the style [
18]. The rejection of pollen in
Papaver rhoeas occurs on the stigmatic surface. Inhibition of incompatible pollen in
P. rhoeas is rapid compared with the relatively slow inhibition in Solanaceae, acting within several minutes [
18]. These differences have led to suggestions that GSI systems have evolved independently several times. In
Brassica, all self-pollen grains fail to adhere strongly to the papilla [
19,
20], and hydration is arrested before the pollen becomes fully turgid [
21]. Different cessation positions of pollen tube growth suggest that they are controlled by different molecular mechanisms. In addition to these three types of SI pollen tube morphology, there is also ovarian SI, in which pollen is rejected at the ovarian interface and its molecular mechanism is unknown [
22].
Seed and fruit setting, the growth of pollen tubes, seed filling, seed germination, and seedling development are influenced by SI in Orchidaceae. Therefore, the capsule set, seed filling, and growth of pollen tubes following self- and cross-pollination are the primary SI indicators [
23]. Due to the long time between pollination and seed maturation—mostly between 6 months and 1 year—the growth of pollen tubes and the capsule set rate are the most important and feasible markers of the SI response. Studies performed on
Pleurothallidinae and
Dendrobium [
2,
23,
24,
25] suggest that the sites of incompatibility reactions vary among different groups of orchids, implying that different SI molecular mechanisms exist in Orchidaceae. In some species, such as
Masdevallia infracta,
Octomeria,
Stelis,
Specklinia, and
Anathallis spp. (except
A. microphyta), pollen after pollination does not germinate [
26], and self-pollination of 26 species of
Restrepia (Orchidaceae) revealed that pollen tubes grow only into the top third of the ovary [
23], while
Pleurothallis adamantinensis and
P. fabiobarrosii are strictly self-incompatible because pollen tube growth ceases near the base of the style [
27]. We identified two SI species in
Dendrobium:
Dendrobium longicornu and
D. chrysanthum. The growth of the self-pollinated pollen tube in both species ceases at the top of the style [
28].
Even studies have shown a high SI species rate in
Dendrobium, the morphology of pollen tube growth, its distribution pattern, and its ancestral status in the
Dendrobium phylogenetic tree remain unclear. As previously mentioned, we have already identified two SI species (
D. longicornu and
D. chrysanthum) by investigating the growth of pollen tubes using fluorescence microscopy [
28]. In this study, in order to know the potential pollen tube morphology in
Dendrobium, we carried out a systematic investigation on the capsule set rate and pollen tube growth over 2 years. Thereafter, we analysed the distribution pattern of the capsule set rate after pollination in the
Dendrobium phylogenetic tree, the distribution pattern of pollen tube growth of SI species in the
Dendrobium phylogenetic tree, and its ancestral status. We conclude that there are several different sites at which the growth of the pollen tube bundle of self-pollinated species stops and the diverse pollen tube bundle phenotypes in
Dendrobium SI species, suggesting that different SI molecular bases might exist between SI species with different pollen morphologies, and that the SI morphologies of the growth of the pollen tube bundle that stopped at the upper one-third of the style might be the main SI type.
2. Results
2.1. The Results Obtained from Artificial Pollination
Self- and cross-pollination were performed in 26
Dendrobium species over a 2-year period (
Table 1). Flowering time for a single flower is generally longer than 6 days, and the longest is 21 days, such as in
D. cariniferum. Flowering times of most species are between 8 and 11 days, such as in
D. catenatum,
D. chrysanthum, and
D. longicornu, while those of
D. farmer and
D. stuposum are the shortest, only 6 days.
In regard to the capsule set rate (CSR), there were some self-sterile species, such as D. aphyllum, D. densiflorum, D. devonianum, D. farmer, D. hancockii, D. jenkinsii, D. longicornu, D. moniliforme, D. polyanthum, and D. stuposum. Of these species, we also found a CSR with cross-pollination of zero in D. devonianum and D. longicornu. However, the CSR was 100% in D. longicornu planted in Malipo, Yunnan Province, China. Our hypothesis is that these two species might not be adapted to the environments of Shenzhen, even after being planted for more than 8 years. In addition, the cross-pollinated capsule set rate (CPCSR) for most species is higher than 50% and reaches nearly 100%, except for D. moniliforme and D. stuposum, which are 34–41% and 37.5–50%, respectively.
We also found some species with a low self-pollinated capsule set rate (SPCSR) (1–24%). These are D. aduncum, D. cariniferum, D. catenatum, D. chrysanthum, D. fimbriatum, D. gratiosissimum, D. harveyanum, D. lindleyi, D. thyrsiflorum, and D. unicum. In these species, the CPCSRs of D. aduncum, D. cariniferum, D. catenatum, and D. unicum are between 0% and 60%. The CSR of D. unicum showed very significant differences between the two years—the CPCSR was 60% in 2015 and 0% in 2016, while the SPCSR was 0% in 2015 and 21.4% in 2016.
Six species, D. chrysotoxum, D. crystallinum, D. denneanum, D. hainanense, D. hercoglossum, and D. linawianum, had a relatively high CSR—35–100% for self-pollination and 41–100% for cross-pollination. Dendrobium hercoglossum also showed large differences in CSR values between the 2 years, with 0% and 57% for self-pollination in 2015 and 2016, respectively, and 36% and 15.5% for cross-pollination in 2015 and 2016, respectively. These differences might also be caused by environmental influences.
2.2. Pollen Tube Growth Results
Based on the results from artificial pollination and the phylogenetic tree of Dendrobium, we selected the species containing all types of CSR and phylogenetic clades and observed pollen tube growth using fluorescence microscopy after self- and cross-pollination over 2 years.
A previous report of
D. chrysanthum and
D. longicornu (Malipo, Yunnan) [
28] stated that
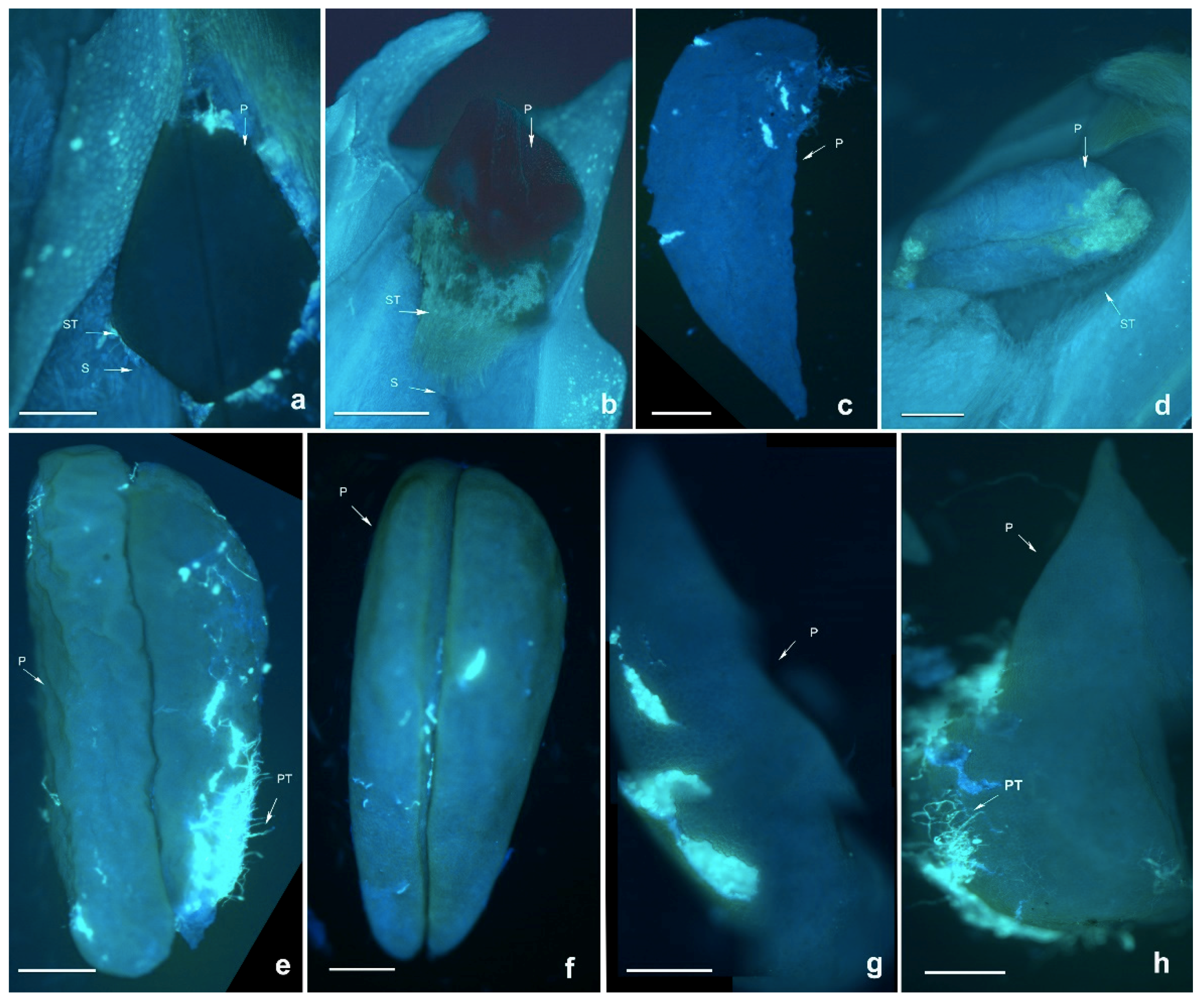
D. longicornu planted in Shenzhen had no seeds after self- and cross-pollination. The pollen tube grew very slowly and was short, with a membrane-like substance coating the pollinium (
Figure 1); which might inhibit the germination of the pollen tube and hinder the pollen tube growth, providing resistance against the unadaptable environment.
Dendrobium lindleyi is a self-sterile species based on artificial pollination (
Table 1). Compared with the pollen tube bundle (PTB) of cross-pollinated plants, the PTB of self-pollinated plants grew more slowly up to 48 h after self-pollination (HASP), when the PTB was at the bottom of the style or had just reached the ovary. Next, the PTB stopped growing. Finally, flower abscission happened. At 24 HASP, black spots began to appear which increased in number over time. This might be due to the degradation of callose in the pollen tube which cannot make the pollen tube fluorescent, leading to the SI phenotype. However, the PTB of cross-pollinated species grew much faster, reaching the ovaries in 36 or 48 HASP (
Figure 2).
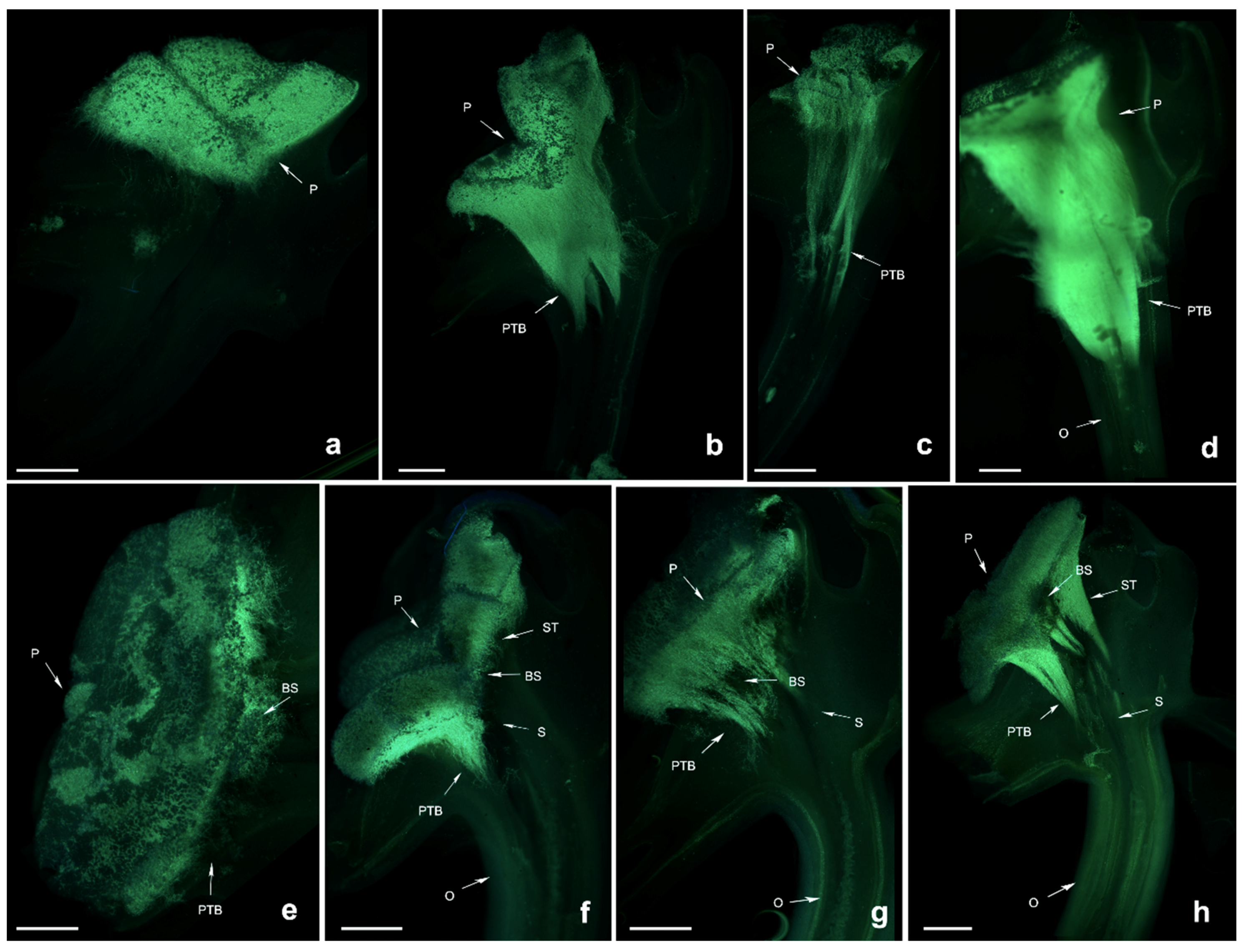
Pollen tubes of
D. devonianum showed similar growth rates after self- and cross-pollination, and black spots appeared in the pollinium and pollen tubes at both 96 HASP and HACP (hours after cross pollination), when the PTB grew up to the upper end of the style (
Figure 3). The zone of black spots became larger at 120 HASP and HACP, indicating the degradation of callose, meaning that the pollen tube cannot be stained. This might be a different phenotype from that of
D. longicornu, showing the lack of adaptability to the environment. These results imply that a style substance or release of some materials from the style already exists when the pollen tube grows up to a certain position in the style that degrades the pollen tube, which shows the self- and cross-incompatibility phenotype.
Dendrobium denneanum is a self- and cross-fertilised species that is present in most individuals based on the artificial pollination results. During pollen tube growth of self-sterile individuals, black spots were present after 96 h of self-pollination, and the zone of black spots became larger until the pollen tube grew into the ovary (
Figure 4). This is similar to
D. devonianum, except that the growth of the pollen tube in
D. devonianum stops at the upper end of the style. This is also similar to pollen tube growth in
D. lindleyi, but the black spots appear later than in
D. lindleyi.
Because of the significantly different CSR results in
D. unicum between the 2 years, we analyse the pollen tube growth of self-sterile plants. The growth of PTB in
D. unicum was similar to that in
D. devonianum,
D. denneanum, and
D. lindleyi, and all of them had black spots (
Figure 5). However, the growth cessation position of PTB was at a certain position of the style in
D. unicum and
D. devonianum, while it was at the ovary and the bottom of the style or the upper part of the ovary in
D. denneanum and
D. lindleyi, respectively
.Based on the artificial pollination results,
Dendrobium densiflorum is a self-sterile species. The pollen tube grew fast after cross-pollination, especially at 48 HACP when the PTB had grown into the ovary (
Figure 6). Although the self-pollination PTB grew much more slowly, there was no growth 48 h after self-pollination, and the growth of PTB stopped at the surface of the stigma or when just entering the style (
Figure 6). We speculate that some substance from the style blocked the growth of the pollen tube, or the blocking response occurred earlier on and the phenotype appearance was delayed.
Dendrobium catenatum has partial self- and partial cross-sterile characteristics. The SPCSR was 11% and 10% in 2015 and 2016, while the CPCSR was 23% and 40% in 2015 and 2016, respectively (
Table 1). For self-incompatible individuals, PTB growth stops at the upper end of the style 48 or 72 h after self-pollination. At nearly the same time, black spots appear in the pollinium and the pollen tube and increase in number over time. This phenotype is also similar to those of
D. unicum,
D. denneanum,
D. lindleyi, and
D. devonianum, although pollen tube growth positions and the timing of the appearance of black spots are different.
Dendrobium moniliforme is a self-sterile and partial cross-fertile species. The SPCSR was 0% over the 2 years, and the CPCSR was 34% and 41% in 2015 and 2016, respectively (
Table 1). We chose the self-sterile and cross-fertile plants by observing the growth of the pollen tube. The PTB values between self- and cross-pollination were significantly different 72 h after pollination, and PTB growth stopped at the upper end of the style (
Figure 7).
Dendrobium hancockii is an artificially pollinated self-sterile species with 0% SPCSR and 100% CPCSR (
Table 1). There was no difference in PTB growth between self- and cross-pollination, and the PTB of self-pollinated plants also grew into the ovaries. Surprisingly, black spots also appeared 72 h after self-pollination, and the black zone increased in size until no pollen tube was observed 120 h after self-pollination (
Figure 8). Pollen tube growth in
D. jenkinsii after self- and cross-pollination was similar to that of
D. hancockii and
D. lindleyi (
Figure 9), suggesting that they might have the same SI molecular mechanism.
Based on the results of self- and cross-pollination and pollen tube growth, we observed a high diversity of self- and cross-sterility and thus many different types of self-incompatible pollen tube morphologies. Therefore, what is the distribution pattern and what is the ancestral status of this morphology?
2.3. Distribution Pattern and Ancestral Status of Self-Incompatibility and Self-Compatibility in Asian Dendrobium Species
We combined the CSR and pollen tube growth results after pollination and identified self-incompatible species and self-compatible species. using the reported
Dendrobium self-incompatible species [
2] and species in this study, we analysed the self-incompatibility and self-compatibility distribution pattern and ancestral status of the mating system in
Dendrobium from Asia.
There were 61 species used in this analysis. Twenty-six species were from this study, and the others were from unpublished data and data from Johansen [
2]. As we were limited by the materials, we focused on the Asian clade of
Dendrobium.
From the distribution of selected species, the samples represented all of the Asian clade (
Figure 10). The selected self-compatibility species were located from the top to the bottom of the Asian
Dendrobium clade, in the order of
D. hainanense,
D. terminale,
D. exile,
D. bellatulum,
D. infundibulum,
D cariniferum,
D. formosum,
D. trigonopus,
D. capillipes,
D.jiaolingense,
D. linawianum,
D. hercoglossum,
D. heterocarpum,
D. findlayanum,
D. scoriarum,
D. catenatum,
D. aduncum,
D. unicum,
D. trantuanii,
D. wardianum,
D. pendulum,
D. crystallinum,
D. lituiflorum,
D. loddigesii,
D. polyanthum,
D. brymerianum,
D. crepidatum,
D. denneanum,
D. gibsonii,
D. fimbriatum,
and D. chrysotoxum. For the self-incompatibility species, their distribution was the same as that of self-compatibility species. From the top to the bottom of the Asian
Dendrobium clade, they were
D. leonis,
D. spatella,
D. pachyphyllum,
D. crumenatum,
D. densiflorum,
D. thyrsiflorum,
D. farmer,
D. longicornu,
D. christyanum,
D. draconis,
D. virgineum,
D. secundum,
D. jenkinsii,
D. lindleyi,
D. denudans,
D. porphyrochilum,
D. strongylanthum,
D. signatum,
D. moniliforme,
D. falconeri,
D. gratiosissimum,
D. devonianum,
D. chrysanthum,
D. moschatum,
D. ellipsophyllum,
D. stuposum,
D. bicameratum,
D. pulchellum,
D. harveyanum,
and D. hancockii. Moreover, nearly each subclade had self-incompatibility and self-compatibility species at the same time.
There were four different self-incompatible phenotypes of pollen tube growth as follows: (1) Membrane-like body coating the pollinium hindered pollinium germination and pollen tube growth, such as in D. longicornu (Shenzhen); (2) pollen tube growth stopped at the upper end of the style, such as in D. chrysanthum, D. moniliforme, D. longicornu (Malipo), and D. densiflorum; (3) no difference in pollen tube growth between self- and cross-pollinated plants, but black spots appeared on the pollen tube when it grew to the bottom of the style or just into the ovary, causing degradation in the self-pollinated pollen tube, for example, in D. hancockii, D. jenkinsii, and D. lindleyi; (4) pollen tube growth stopped at a certain position of the style and black spots appeared in D. devonianum and D. unicum. The second SI type had a wider phylogenetic distribution, covering the bottom and top locations of the phylogenetic tree, and might be the main SI phenotype in Dendrobium.
Based on the ancestral status reconstruction results, self-compatibility was the ancestral status in most subclades, but this was equivocal in the Dendrobium clade.
3. Discussion
3.1. Artificial Pollination and CSR
In this study, 26 species were investigated over 2 years. For certain species with a limited number of individuals, there were comparatively less pollinated flowers, only eight to ten flowers, so more pollination may have been required to reveal the real situation. Nevertheless, we carried out the pollination experiments over 2 years to make up for the lack of quantity. For most species with many individuals, there were sufficient pollinated flowers, i.e., more than 100 flowers. We converted the results from more than 100 flowers to 100 flowers for normalization.
We identified nine species with complete self-sterility in this study. These were D. aphyllum, D. densiflorum, D. devonianum, D. farmer, D. hancockii, D. jenkinsii, D. longicornu (Malipo), D. moniliforme, and D. stuposum, including the cross-sterility species D. devonianum and the partial cross-sterility species D. moniliforme, D. aphyllum, D. stuposum, and D. jenkinsii.
There were species with different results between the 2 years, such as D. unicum, for which the SPCSR was 0% in 2015 and 21.4% in 2016, while the number of pollinated flowers in 2016 was less than that in 2015, at 52 in 2015 and 28 in 2016. This difference might have been caused by the different levels of pollination of plants between the 2 years or changes in the environment. Because of the variation in self-sterility, we treated the species as partially self-incompatible in the following analysis.
One species was also reported by Johansen [
2]. The
D. lindleyi results reported by Johansen showed self- and cross-incompatibility. However, the results from our study showed self-incompatible and cross-compatible species, which was different from the results of Johansen. In the study by Johansen, cross-pollination was carried out in only two flowers from one individual, which was unrepresentative. In our study, we carried out a pollination experiment in at least 100 flowers. Therefore, we used our results for the next analysis.
3.2. Diversity of Pollen Tube Growth
We analysed nine species containing all the pollen tube growth types from 26 Dendrobium species. The high diversity observed, such as the membrane-like body coating of the pollinium that inhibits germination and limits growth in D. longicornu (Shenzhen), exceeded our expectations. For D. devonianum, the pollen tubes of self- and cross-pollinated species grew into the style and stopped at a certain position; then, black spots appeared on the pollen tube and the pollinium, leading to self- and cross-incompatibility. These spots might be another phenotype that is resistant to adverse situations, in contrast to D. longicornu. The third phenotype was found in D. jenkinsii, D. hancockii, and D. lindleyi. The pollen tube of self-pollinated species grew into the ovary or at the bottom of the style, and then black spots appeared, but the appearance time was earlier than that in D. devonianum, which is 72 and 96 h, respectively. In addition, pollen tube growth stopped at the upper part of the style after self-pollination in D. chrysanthum, D. densiflorum, D. longicornu (Malipo), and D. thyrsiflorum. In this type, the time at which pollen tube growth stopped in D. longicornu was much longer than that in the other three species (96 h and 24–48 h, respectively). For the different times of pollen tube growth cessation, we speculate that the SI determinants of D. longicornu might be different from that of D. chrysanthum, D. densiflorum, and D. thyrsiflorum.
Pollen tube growth stopped at different positions in the style or ovary, which is consistent with previous reports in other orchids. For example, pollen tube growth stopped at certain style positions in
P. adamantinensis and
P. fabiobarrosii, while it stopped at the ovary in
Restrepia species, and the pollinia did not germinate in
M. infracta,
Octomeria,
Stelis, and
Specklinia [
2,
23,
26,
27]. This suggests that there are a variety of SI determinants in the Orchidaceae, even in
Dendrobium.
3.3. Distribution Pattern of Self-Incompatibility and Self-Compatibility
Next, we investigated how the different degrees of self- and cross-incompatibility and types of pollen tube growth states after self-pollination are distributed in the Dendrobium phylogenetic tree as well as investigating their ancestral statuses.
We concluded that the SI type represented by D. chrysanthum was distributed from the bottom to the top of the Dendrobium phylogenetic tree, which might be the main SI type of Dendrobium, while other SI types were distributed in certain phylogenetic positions. Therefore, we consider that SI in Dendrobium has undergone many independent evolutionary processes, controlled by multiple SI determinants.
By reconstructing the ancestral status of SI in the Asian
Dendrobium clade, we found that self-compatibility was the SI ancestral status in most Asian
Dendrobium subclades, which was consistent with the findings of Xu but inconsistent with those in Cruciferae and Solanaceae [
29,
30]. As the majority of orchid species are self-compatible, species at the root of the Orchidaceae phylogenetic tree were also self-compatible and
Dendrobium species evolved 36.7 million years ago (Mya) [
31]. Therefore, we suggest that SI in
Dendrobium also evolved recently, and might be
Dendrobium- or Orchidaceae-specific.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}