A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress

 ,
,  , , ,
, , ,  and
and 
Abstract
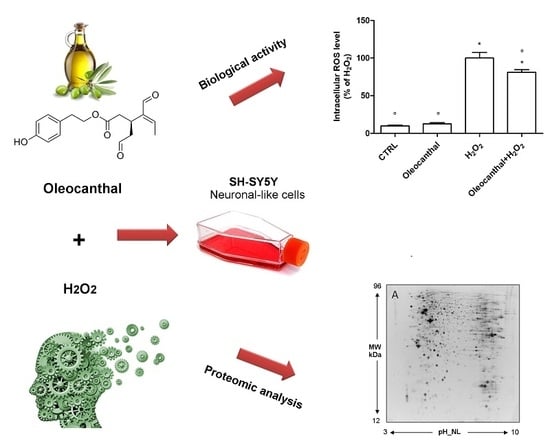
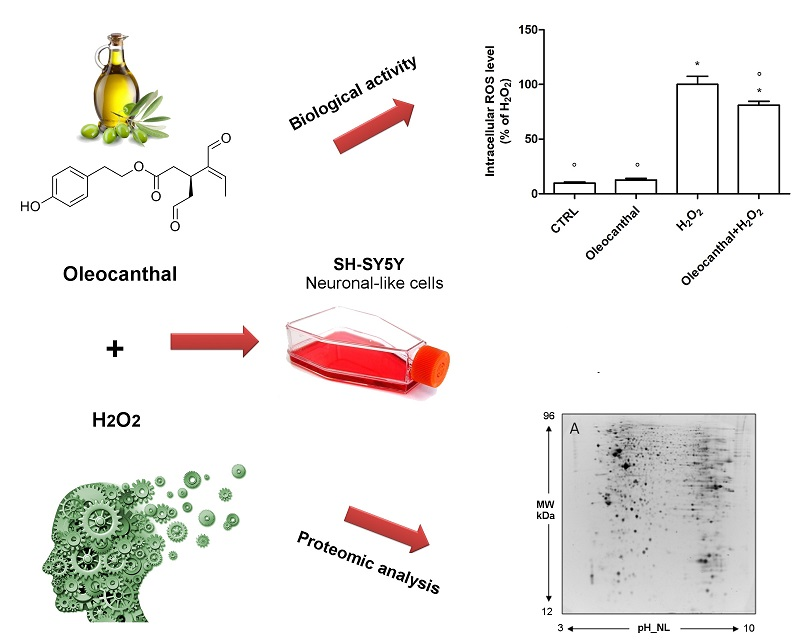
1. Introduction
2. Results
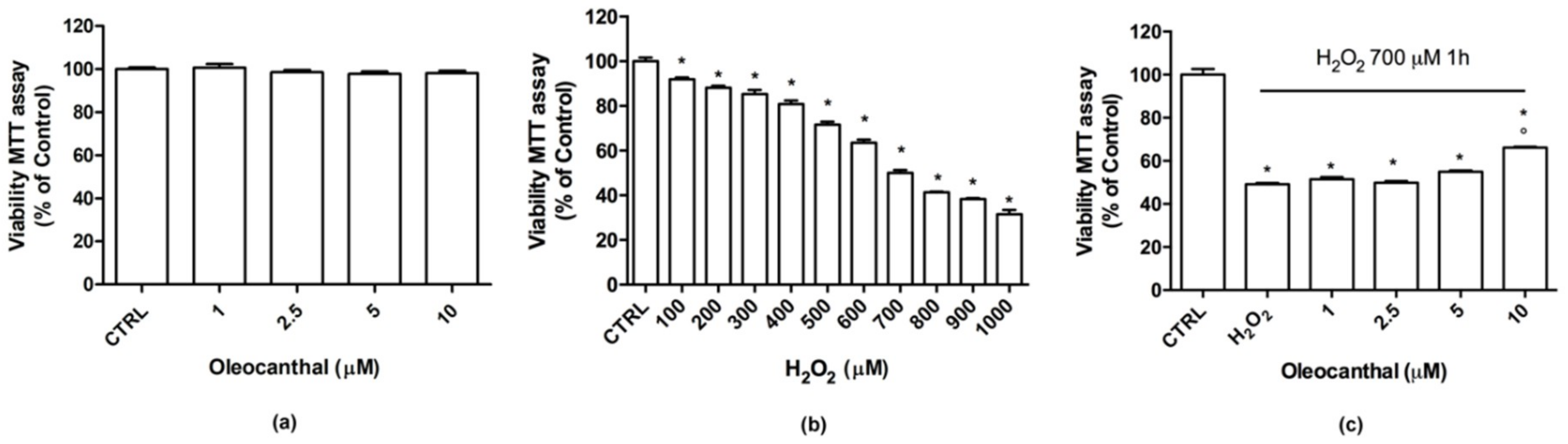
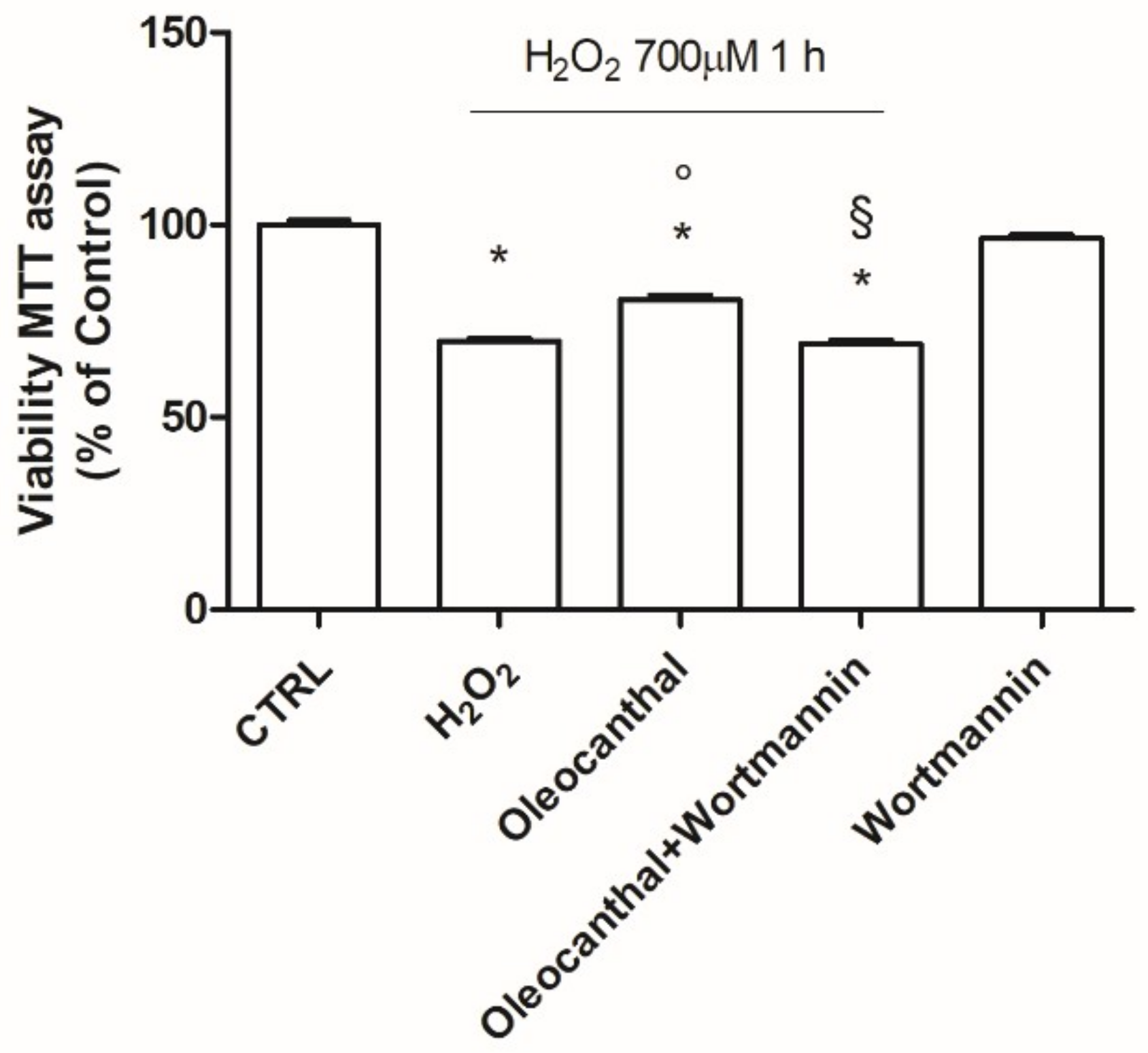
2.1. Neuroprotective Effects of Oleocanthal against H2O2-Induced Damage
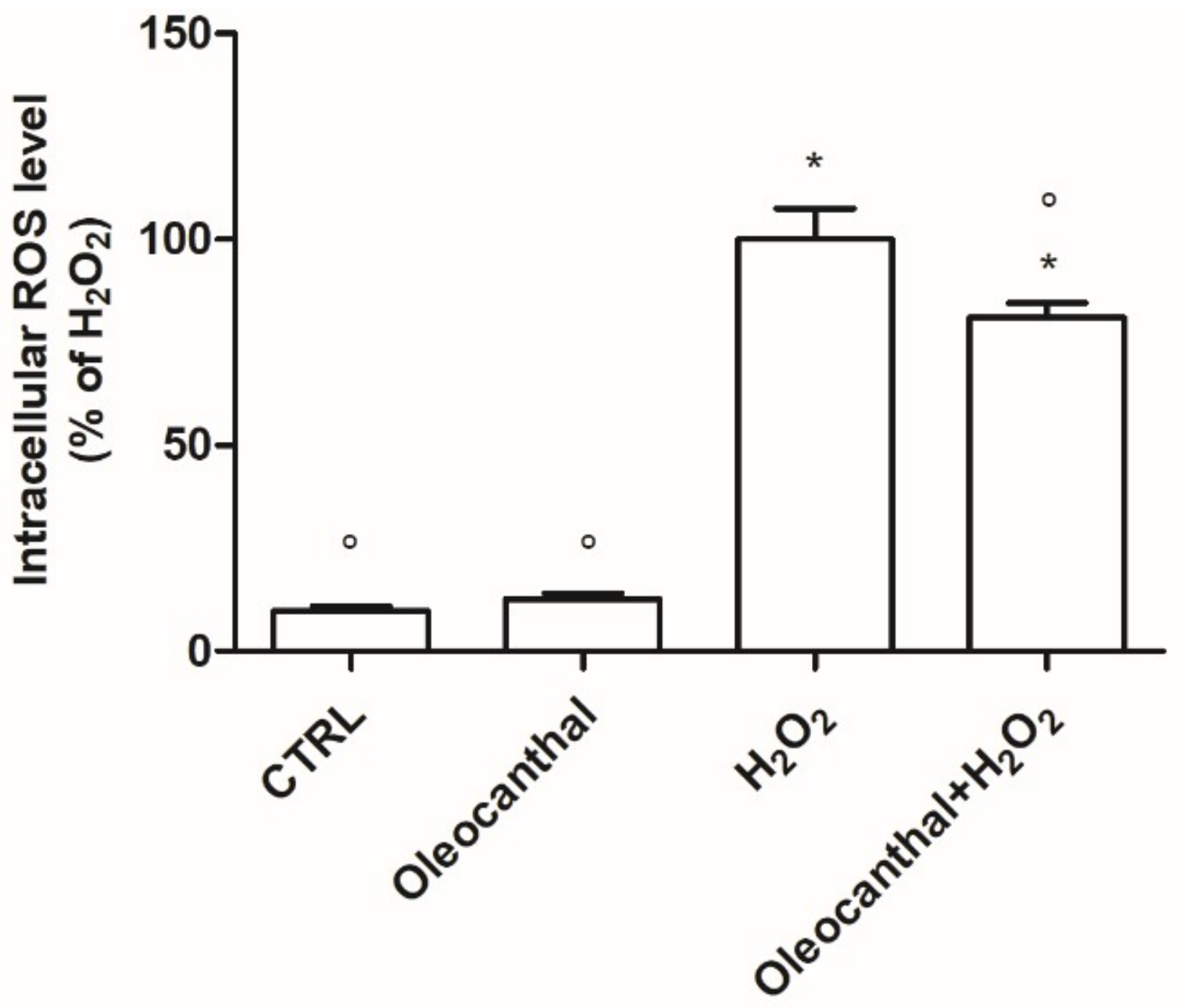
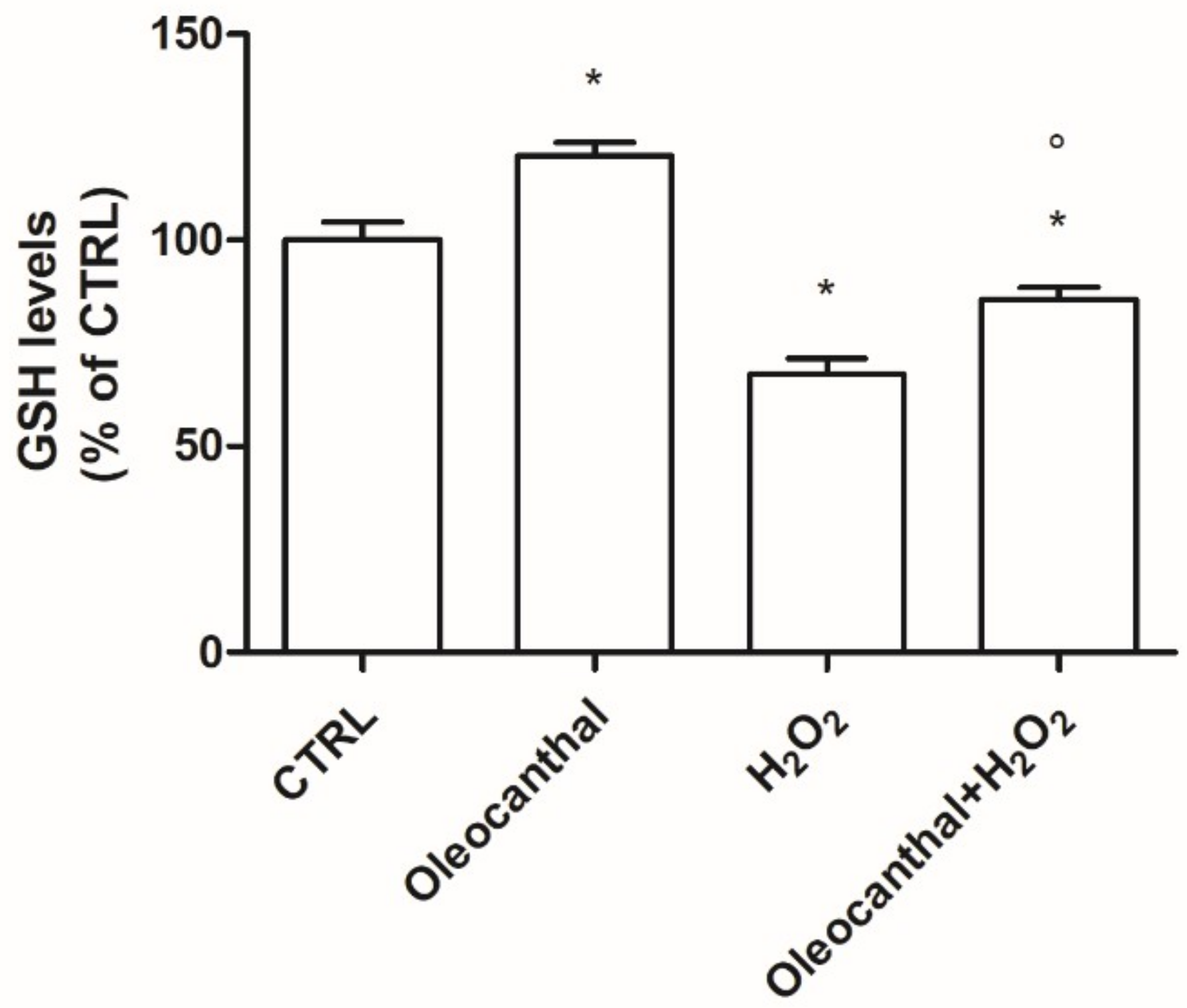
2.2. Antioxidant Activity of Oleocanthal against H2O2-Induced Oxidative Stress
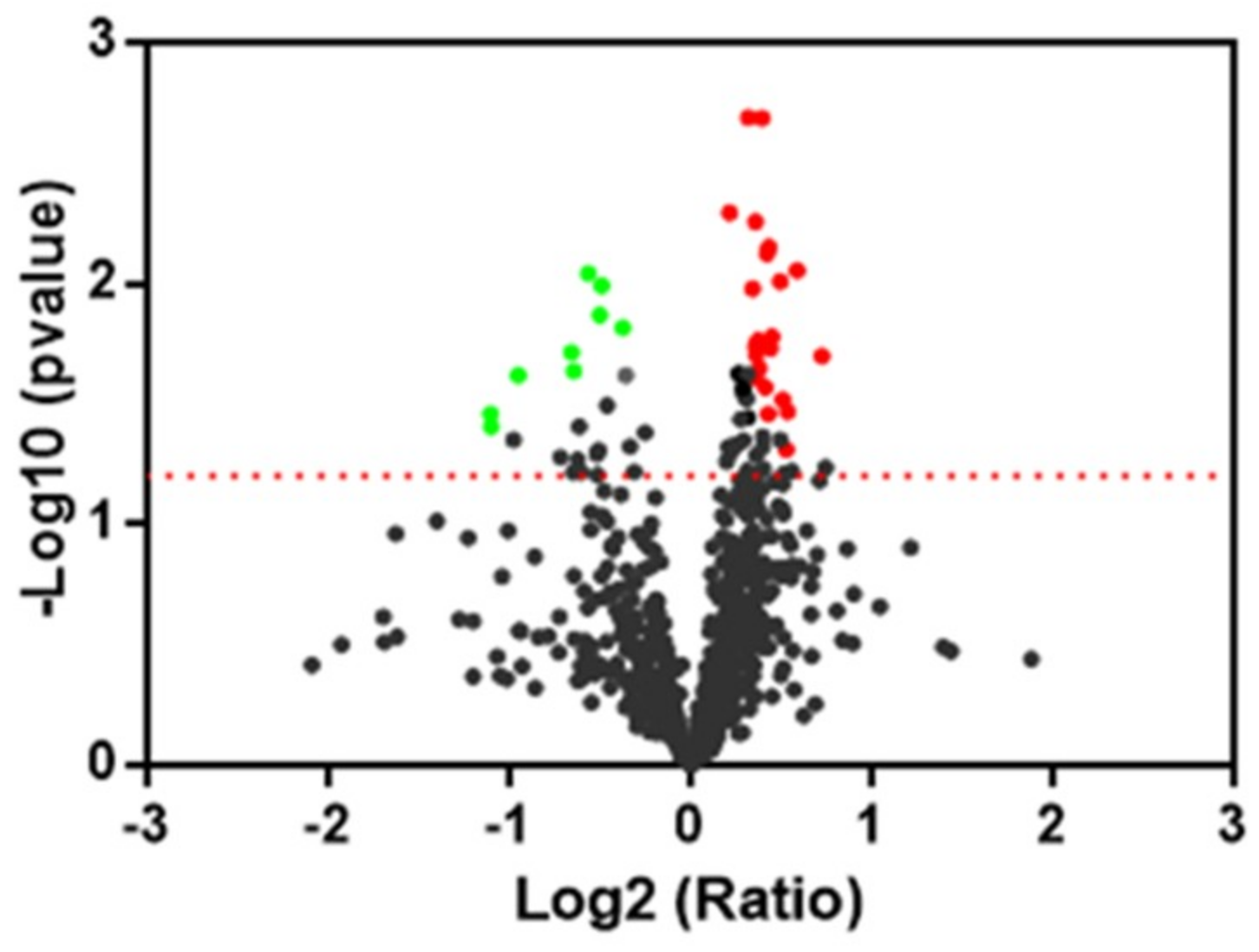
2.3. Protein Expression Analysis
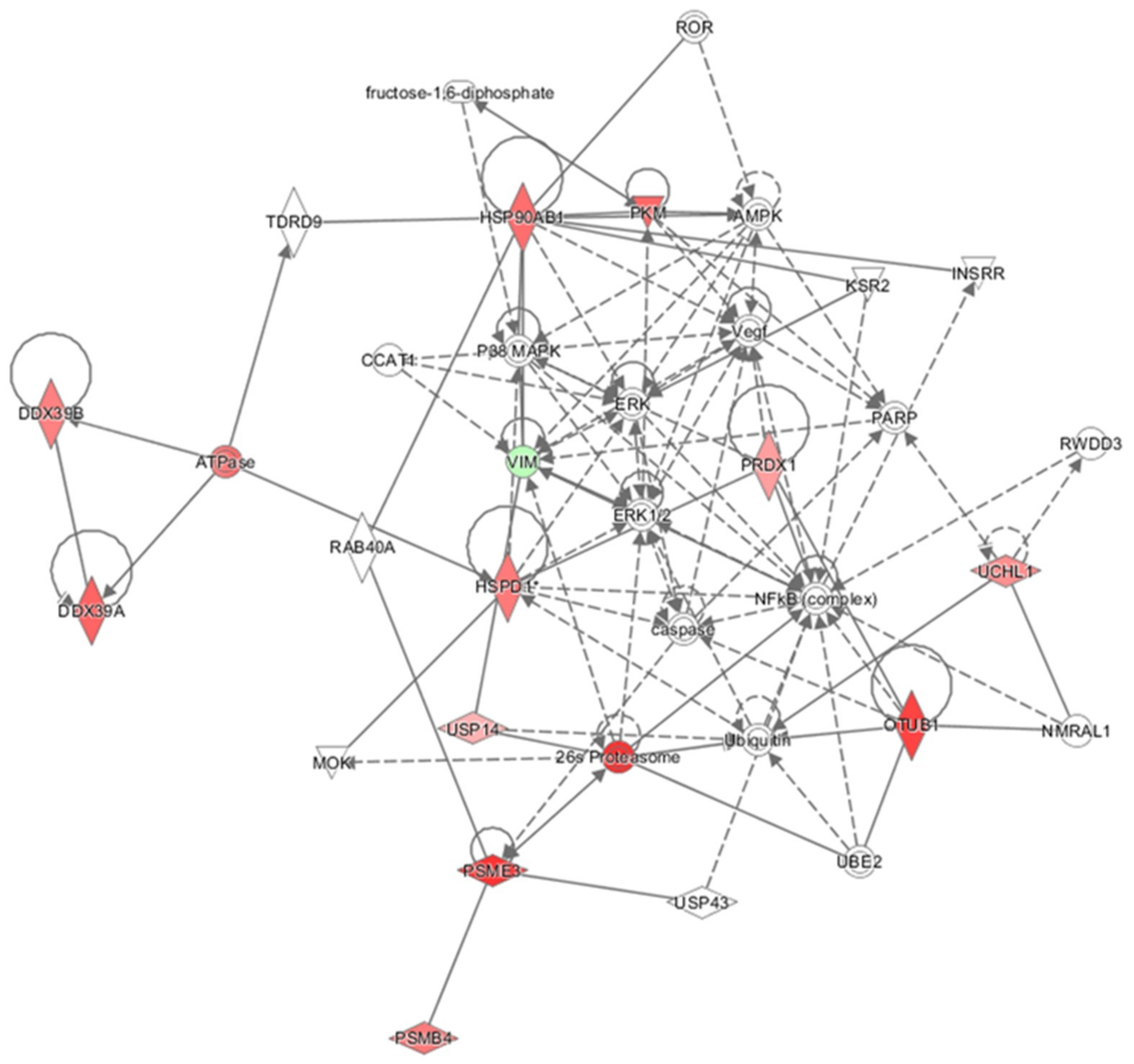
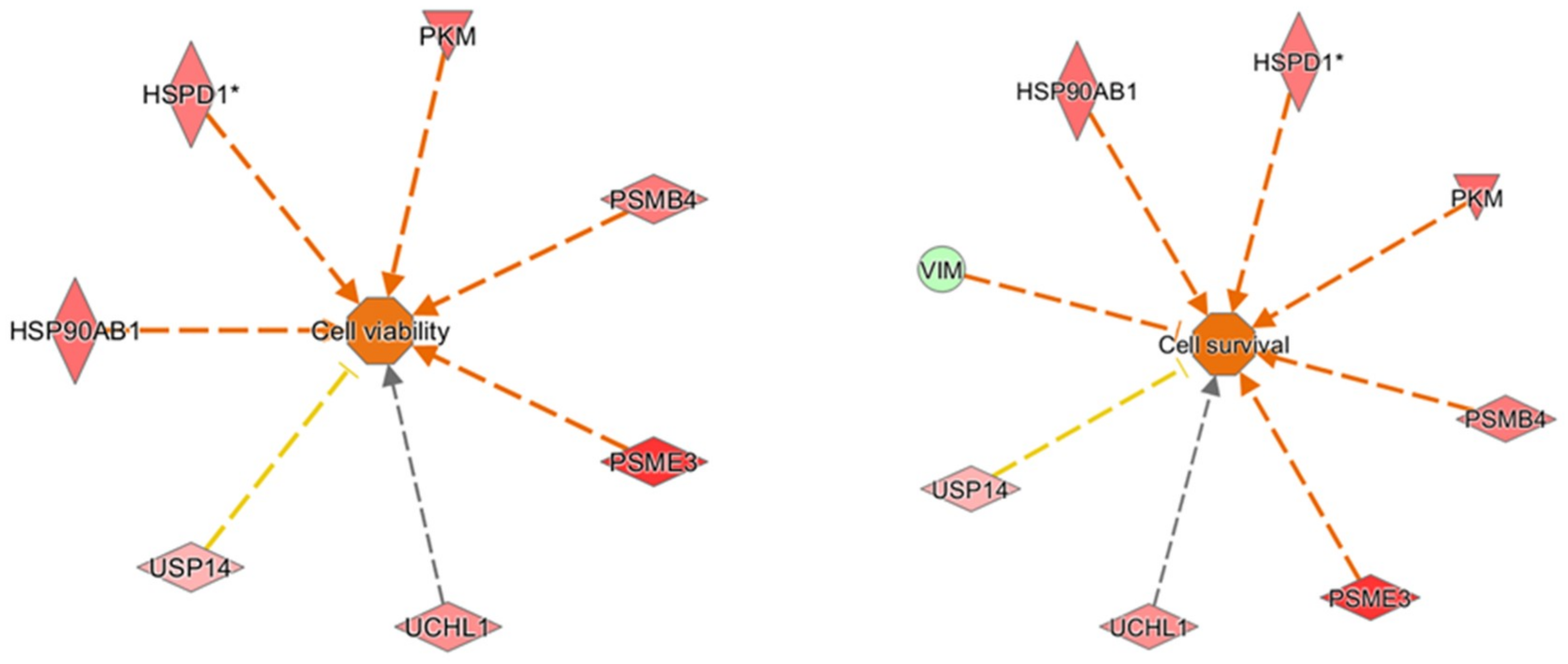
2.4. Pathway and Network Analysis
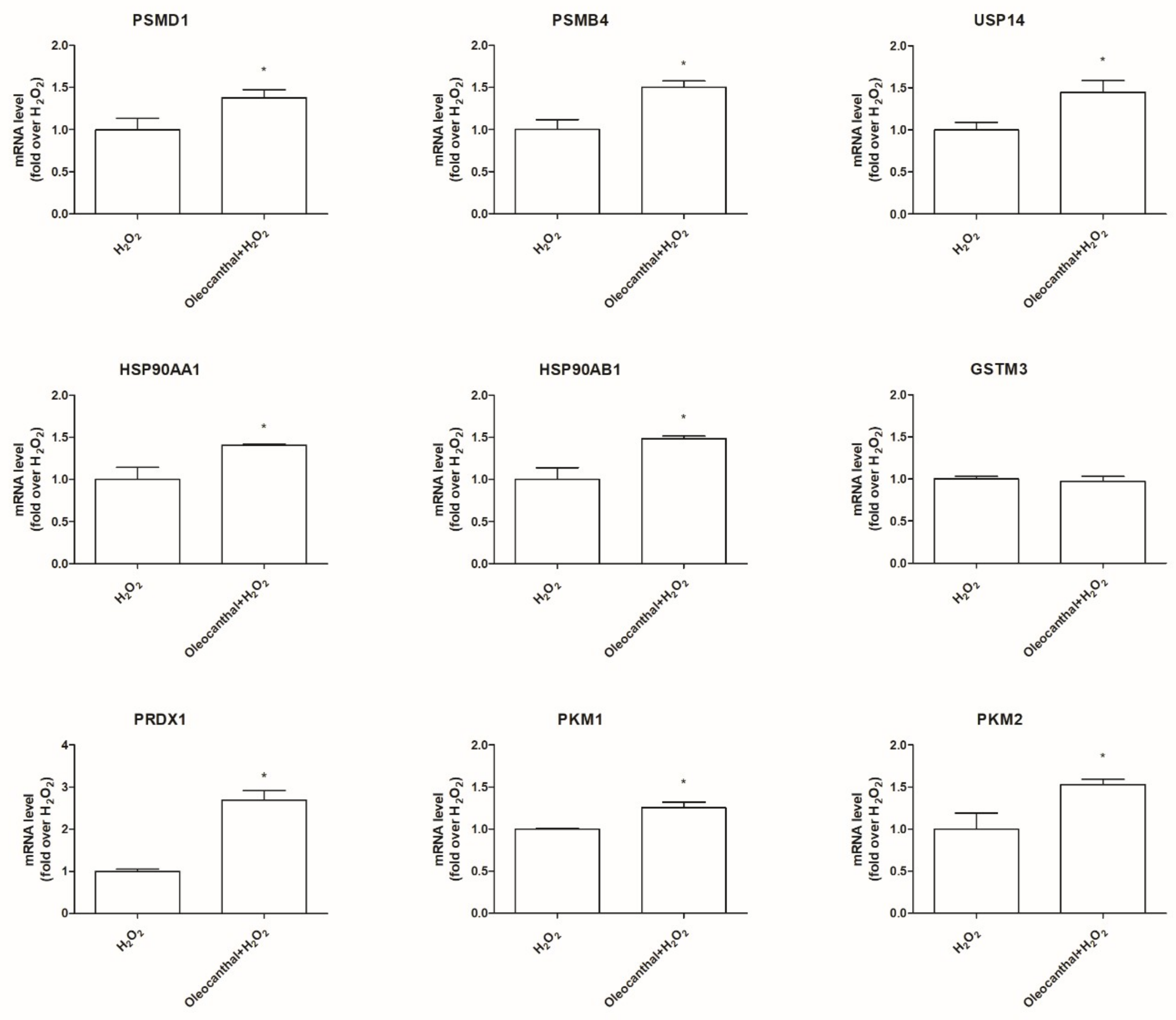
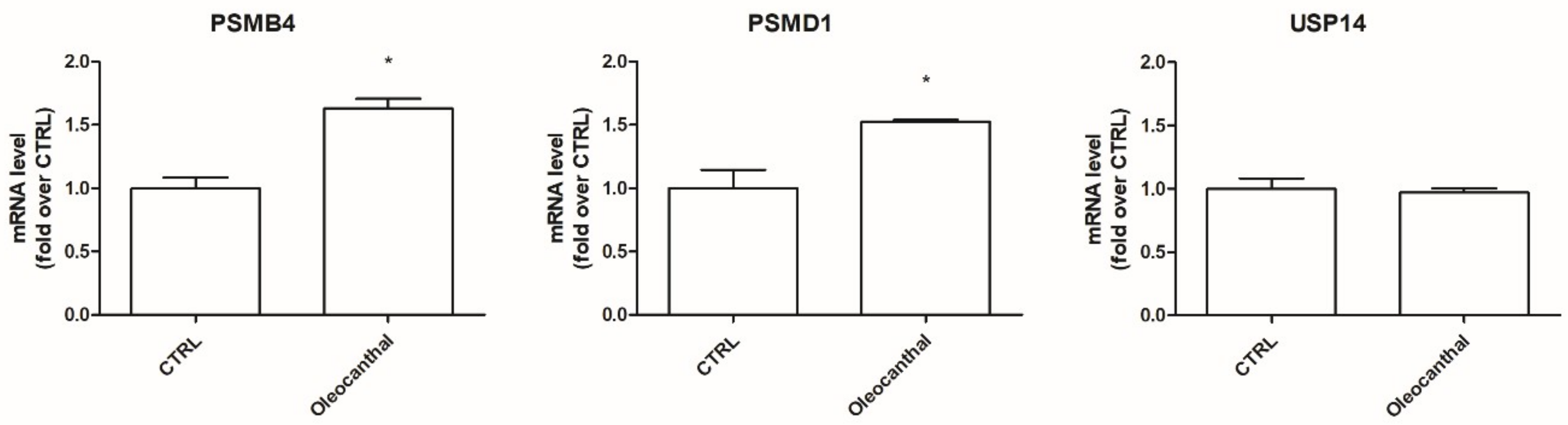
2.5. Transcriptional Validation of Proteomic Data Using RT-Polymerase Chain Reaction (PCR) Assays
2.6. Role of Oleocanthal in the Modulation of the Ubiquitin–Proteasome System
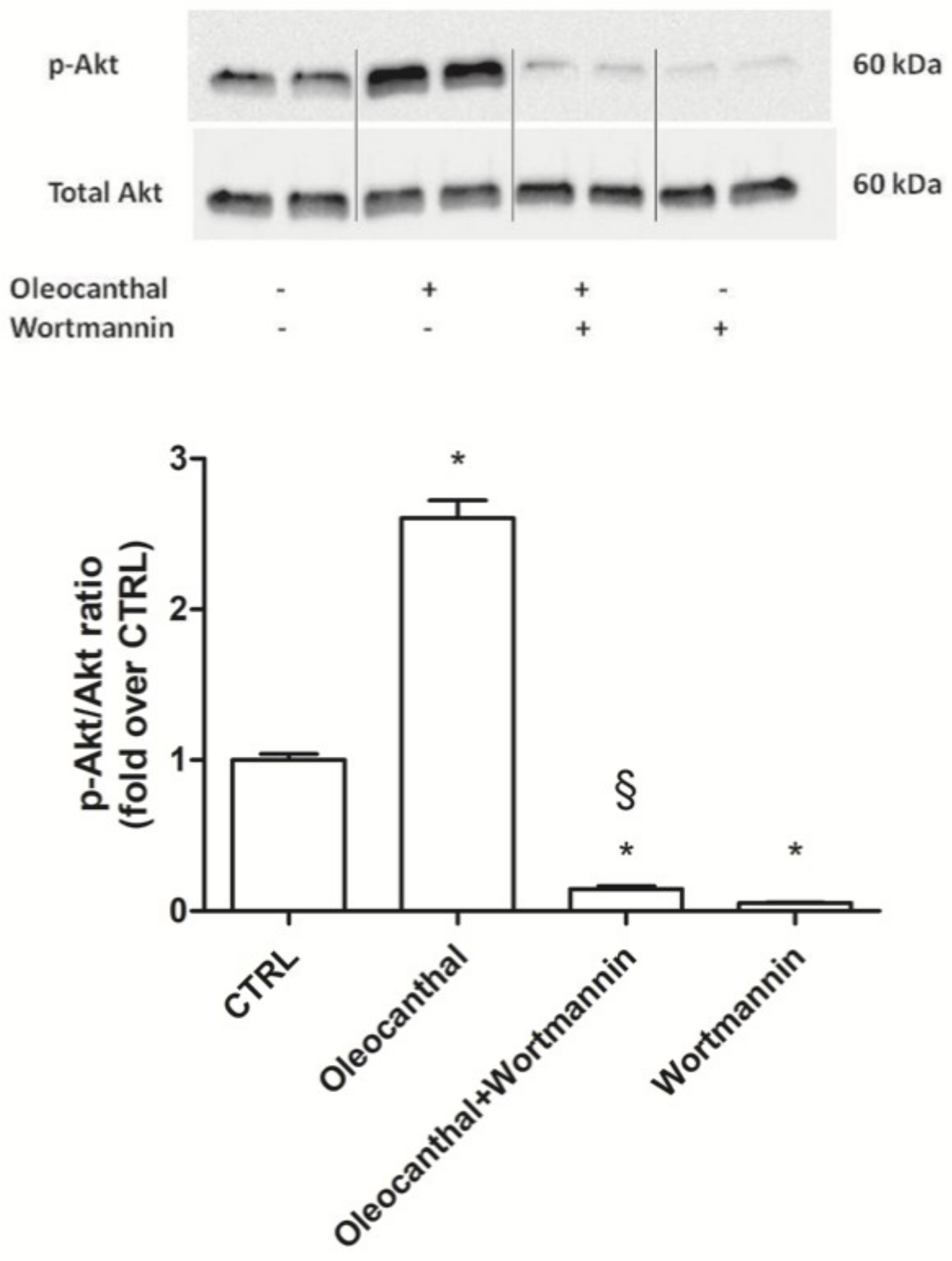
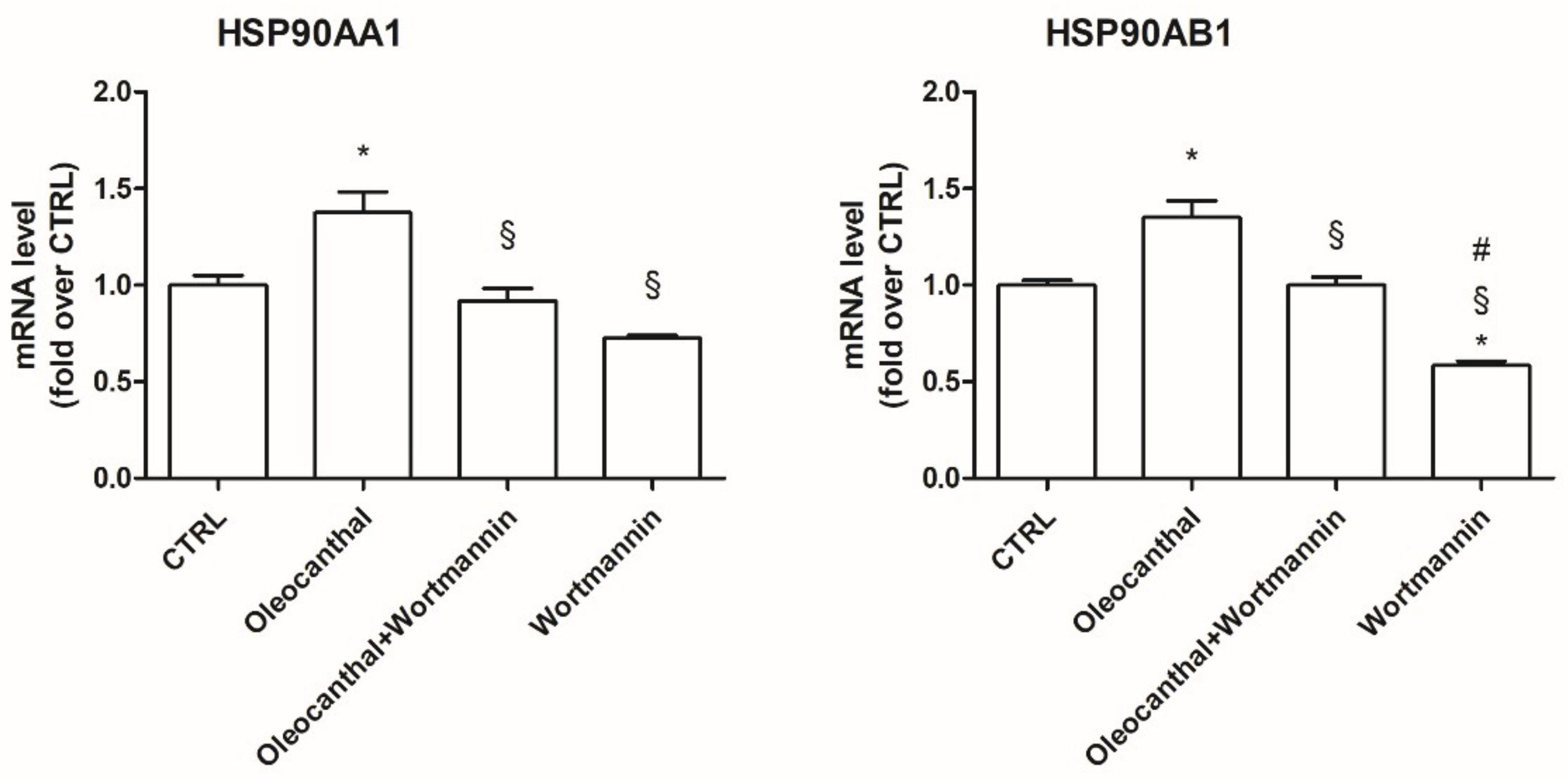
2.7. Involvement of HSP90 in Oleocanthal Protection against Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatments
4.3. MTT Assay
4.4. Intracellular ROS Production Assay
4.5. Intracellular GSH Levels Assay
4.6. RNA Extraction
4.7. Analysis of mRNA Levels by Reverse Transcriptase Polymerase Chain Reaction
4.8. Western Blotting
4.9. Sample Preparation for Proteomic Analysis
4.10. 2DE Analysis
4.11. 2DE Statistical Analysis
4.12. Spot Digestion and Protein Identification
4.13. Network Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2DE | Two-dimensional electrophoresis |
| AD | Alzheimer’s disease |
| ANOVA | Analysis of variance |
| BBB | Blood–brain barrier |
| BSA | Bovine serum albumin |
| CID | Collision-induced dissociation |
| DCFH-DA | 2,7-Dichlorodihydrofluorescein diacetate |
| DDA | Data dependent acquisition mode |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| DTT | Ditiothreitol |
| ECL | Enhanced chemiluminescence |
| FMHW | Full width at half maximum |
| GSH | Reduced glutathione |
| Gstm3 | Glutathione S-transferase mu 3 |
| IEF | Isoelectrofocusing |
| IPA | Ingenuity pathways aanalysis |
| MCB | Monochlorobimane |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5diphenyl-tetrazolium bromide) |
| NOX | Nicotinamide adenine dinucleotide phosphate oxidase |
| PBS | Phosphate-buffered saline |
| PCR | Polymerase chain reaction |
| PD | Parkinson’s disease |
| Prdx | Peroxiredoxin |
| Prk | Pyruvate kinase |
| Psmb4 | Proteasome subunit β 4 |
| Psmd1 | Proteasome 26S subunit non-ATPase 1 |
| RA | All trans retinoic acid |
| RIPA | Radioimmunoprecipitation assay |
| ROS | Reactive oxygen species |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| Usp14 | Ubiquitin carboxyl-terminal hydrolase 14 |
References
- Hampel, H.; Frank, R.; Broich, K.; Teipel, S.J.; Katz, R.G.; Hardy, J.; Herholz, K.; Bokde, A.L.; Jessen, F.; Hoessler, Y.C.; et al. Biomarkers for Alzheimer’s disease: Academic, industry and regulatory perspectives. Nat. Rev. Drug Discov. 2010, 9, 560–574. [Google Scholar] [CrossRef] [PubMed]
- Mandel, S.; Grunblatt, E.; Riederer, P.; Gerlach, M.; Levites, Y.; Youdim, M.B. Neuroprotective strategies in Parkinson’s disease: An update on progress. CNS Drugs 2003, 17, 729–762. [Google Scholar] [CrossRef] [PubMed]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.L.; van Remmen, H.; Drake, J.A.; Yang, H.; Guo, Z.M.; Kewitt, K.; Walter, C.A.; Richardson, A. Does oxidative damage to dna increase with age? Proc. Natl. Acad. Sci. USA 2001, 98, 10469–10474. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I. Neuroprotection by dietary antioxidants: New age of research. Die Nahrung 2002, 46, 381–382. [Google Scholar] [CrossRef]
- Tatton, W.G.; Chalmers-Redman, R.; Brown, D.; Tatton, N. Apoptosis in parkinson’s disease: Signals for neuronal degradation. Ann. Neurol. 2003, 53 (Suppl. 3), S61–S72. [Google Scholar] [CrossRef]
- Jenner, P.; Olanow, C.W. Oxidative stress and the pathogenesis of Parkinson’s disease. Neurology 1996, 47, 161S–170S. [Google Scholar] [CrossRef]
- Henchcliffe, C.; Beal, M.F. Mitochondrial biology and oxidative stress in Parkinson disease pathogenesis. Nat. Clin. Pract. Neurol. 2008, 4, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Boveris, A. Brain mitochondrial dysfunction and oxidative damage in Parkinson’s disease. J. Bioenerg. Biomembr. 2009, 41, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, A.; Perry, G.; Aliev, G.; Hirai, K.; Takeda, A.; Balraj, E.K.; Jones, P.K.; Ghanbari, H.; Wataya, T.; Shimohama, S.; et al. Oxidative damage is the earliest event in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2001, 60, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, A.; Perry, G.; Pappolla, M.A.; Wade, R.; Hirai, K.; Chiba, S.; Smith, M.A. RNA oxidation is a prominent feature of vulnerable neurons in Alzheimer’s disease. J. Neurosci. 1999, 19, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Castellani, R.J.; Harris, P.L.; Sayre, L.M.; Fujii, J.; Taniguchi, N.; Vitek, M.P.; Founds, H.; Atwood, C.S.; Perry, G.; Smith, M.A. Active glycation in neurofibrillary pathology of Alzheimer disease: N(epsilon)-(carboxymethyl) lysine and hexitol-lysine. Free Radic. Biol. Med. 2001, 31, 175–180. [Google Scholar] [CrossRef]
- Shlisky, J.; Bloom, D.E.; Beaudreault, A.R.; Tucker, K.L.; Keller, H.H.; Freund-Levi, Y.; Fielding, R.A.; Cheng, F.W.; Jensen, G.L.; Wu, D.; et al. Nutritional considerations for healthy aging and reduction in age-related chronic disease. Adv. Nutr. 2017, 8, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Féart, C.; Samieri, C.; Barberger-Gateau, P. Mediterranean diet and cognitive function in older adults. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Loughrey, D.G.; Lavecchia, S.; Brennan, S.; Lawlor, B.A.; Kelly, M.E. The impact of the Mediterranean diet on the cognitive functioning of healthy older adults: A systematic review and meta-analysis. Adv. Nutr. 2017, 8, 571–586. [Google Scholar] [PubMed]
- Angeloni, C.; Malaguti, M.; Barbalace, M.C.; Hrelia, S. Bioactivity of olive oil phenols in neuroprotection. Int. J. Mol. Sci. 2017, 18, 2230. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, F.; Maccaglia, A.; Pietraforte, D.; Turco, L.; Minetti, M. Phenolic antioxidants and the protection of low density lipoprotein from peroxynitrite-mediated oxidations at physiologic CO2. J. Agric. Food Chem. 2004, 52, 2866–2874. [Google Scholar] [CrossRef] [PubMed]
- Petroni, A.; Blasevich, M.; Salami, M.; Papini, N.; Montedoro, G.F.; Galli, C. Inhibition of platelet aggregation and eicosanoid production by phenolic components of olive oil. Thromb. Res. 1995, 78, 151–160. [Google Scholar] [CrossRef]
- Maiuri, M.C.; de Stefano, D.; di Meglio, P.; Irace, C.; Savarese, M.; Sacchi, R.; Cinelli, M.P.; Carnuccio, R. Hydroxytyrosol, a phenolic compound from virgin olive oil, prevents macrophage activation. Naunyn. Schmiedebergs Arch. Pharmacol. 2005, 371, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Qosa, H.; Batarseh, Y.S.; Mohyeldin, M.M.; El Sayed, K.A.; Keller, J.N.; Kaddoumi, A. Oleocanthal enhances amyloid-β clearance from the brains of TgSwDI mice and in vitro across a human blood-brain barrier model. ACS Chem. Neurosci. 2015, 6, 1849–1859. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, V.; Sacchi, R. Oleocanthal in olive oil: Between myth and reality. Mol. Nutr. Food Res. 2006, 50, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Servili, M.; Esposto, S.; Fabiani, R.; Urbani, S.; Taticchi, A.; Mariucci, F.; Selvaggini, R.; Montedoro, G.F. Phenolic compounds in olive oil: Antioxidant, health and organoleptic activities according to their chemical structure. Inflammopharmacology 2009, 17, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, G.K.; Keast, R.S.; Morel, D.; Lin, J.; Pika, J.; Han, Q.; Lee, C.H.; Smith, A.B.; Breslin, P.A. Phytochemistry: Ibuprofen-like activity in extra-virgin olive oil. Nature 2005, 437, 45–46. [Google Scholar] [CrossRef] [PubMed]
- Abuznait, A.H.; Qosa, H.; Busnena, B.A.; El Sayed, K.A.; Kaddoumi, A. Olive-oil-derived oleocanthal enhances β-amyloid clearance as a potential neuroprotective mechanism against Alzheimer’s disease: In vitro and in vivo studies. ACS Chem. Neurosci. 2013, 4, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.C.; Margarucci, L.; Tosco, A.; Riccio, R.; Casapullo, A. New insights on the interaction mechanism between tau protein and oleocanthal, an extra-virgin olive-oil bioactive component. Food Funct. 2011, 2, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.; Roth, W.; Lacor, P.; Smith, A.B.; Blankenship, M.; Velasco, P.; de Felice, F.; Breslin, P.; Klein, W.L. Alzheimer’s-associated Aβ oligomers show altered structure, immunoreactivity and synaptotoxicity with low doses of oleocanthal. Toxicol. Appl. Pharmacol. 2009, 240, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sperry, J.B.; Crowe, A.; Trojanowski, J.Q.; Smith, A.B.; Lee, V.M. Inhibition of tau fibrillization by oleocanthal via reaction with the amino groups of tau. J. Neurochem. 2009, 110, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Rosignoli, P.; Fuccelli, R.; Fabiani, R.; Servili, M.; Morozzi, G. Effect of olive oil phenols on the production of inflammatory mediators in freshly isolated human monocytes. J. Nutr. Biochem. 2013, 24, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Piras, S.; Furfaro, A.L.; Brondolo, L.; Passalacqua, M.; Marinari, U.M.; Pronzato, M.A.; Nitti, M. Differentiation impairs bach1 dependent ho-1 activation and increases sensitivity to oxidative stress in SH-SY5Y neuroblastoma cells. Sci. Rep. 2017, 7, 7568. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Sinha, I.; Calcagnotto, A.; Trushin, N.; Haley, J.S.; Schell, T.D.; Richie, J.P. Oral supplementation with liposomal glutathione elevates body stores of glutathione and markers of immune function. Eur. J. Clin. Nutr. 2018, 72, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.G.; Matts, R.L. Hsp90 functions to balance the phosphorylation state of Akt during C2C12 myoblast differentiation. Cell Signal. 2005, 17, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Medina, E.; Vargas, J.; Brenes, M.; De Castro, A. In vitro activity of olive oil polyphenols against helicobacter pylori. J. Agric. Food Chem. 2007, 55, 680–686. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Carrasco-Pancorbo, A.; Nevedomskaya, E.; Mayboroda, O.A.; Deelder, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Exploratory analysis of human urine by LC-ESI-TOF MS after high intake of olive oil: Understanding the metabolism of polyphenols. Anal. Bioanal. Chem. 2010, 398, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.N.; Marques, C.; Rodrigues, E.; Henderson, C.J.; Wolf, C.R.; Pereira, P.; Gama, M.J. Ubiquitin-proteasome system impairment and MPTP-induced oxidative stress in the brain of C57BL/6 wild-type and GSTP knockout mice. Mol. Neurobiol. 2013, 47, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell. Biol. 2005, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, N.; Piccirillo, R.; Hourez, R.; Venkatraman, P.; Goldberg, A.L. Cathepsins L and Z are critical in degrading polyglutamine-containing proteins within lysosomes. J. Biol. Chem. 2012, 287, 17471–17482. [Google Scholar] [CrossRef] [PubMed]
- Coux, O.; Tanaka, K.; Goldberg, A.L. Structure and functions of the 20s and 26s proteasomes. Annu. Rev. Biochem. 1996, 65, 801–847. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, C.; Tabrizi, S.J. The ubiquitin-proteasome system in neurodegeneration. Antioxid. Redox Signal. 2014, 21, 2302–2321. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Hara, K.; Sethi, K.D.; Morgan, J.C.; Borlongan, C.V. Increased 8-OHDG levels in the urine, serum, and substantia nigra of hemiparkinsonian rats. Brain Res. 2007, 1133, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Lu, Y.; Prado, M.A.; Shi, Y.; Tian, G.; Sun, S.; Elsasser, S.; Gygi, S.P.; King, R.W.; Finley, D. Usp14 deubiquitinates proteasome-bound substrates that are ubiquitinated at multiple sites. Nature 2016, 532, 398–401. [Google Scholar] [CrossRef] [PubMed]
- van Assema, D.M.; Lubberink, M.; Bauer, M.; van der Flier, W.M.; Schuit, R.C.; Windhorst, A.D.; Comans, E.F.; Hoetjes, N.J.; Tolboom, N.; Langer, O.; et al. Blood-brain barrier p-glycoprotein function in Alzheimer’s disease. Brain 2012, 135, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Batarseh, Y.S.; Mohamed, L.A.; Al Rihani, S.B.; Mousa, Y.M.; Siddique, A.B.; El Sayed, K.A.; Kaddoumi, A. Oleocanthal ameliorates amyloid-β oligomers’ toxicity on astrocytes and neuronal cells: In vitro studies. Neuroscience 2017, 352, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 2008, 283, 18473–18477. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Gehrmann, M.; Brunet, M.; Multhoff, G.; Garrido, C. Intracellular and extracellular functions of heat shock proteins: Repercussions in cancer therapy. J. Leukoc. Biol. 2007, 81, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Taiyab, A.; Sreedhar, A.S.; Rao, C.M. Hsp90 inhibitors, GA and 17AAG, lead to ER stress-induced apoptosis in rat histiocytoma. Biochem. Pharmacol. 2009, 78, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Fujita, N.; Tsuruo, T. Modulation of Akt kinase activity by binding to Hsp90. Proc. Natl. Acad. Sci. USA 2000, 97, 10832–10837. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Tiong, C.X.; Bian, J.S. Hydrogen sulfide protects SH-SY5Y cells against 6-hydroxydopamine-induced endoplasmic reticulum stress. Am. J. Physiol. Cell Physiol. 2012, 303, C81–C91. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.A.; Tindale, L.; Cumming, R.C. Age-dependent metabolic dysregulation in cancer and Alzheimer’s disease. Biogerontology 2014, 15, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Wärmländer, S.K.; Gräslund, A.; Abrahams, J.P. Non-chaperone proteins can inhibit aggregation and cytotoxicity of Alzheimer amyloid β peptide. J. Biol. Chem. 2014, 289, 27766–27775. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Yang, K.S.; Kang, S.W.; Woo, H.A.; Chang, T.S. Controlled elimination of intracellular H2O2: Regulation of peroxiredoxin, catalase, and glutathione peroxidase via post-translational modification. Antioxid. Redox Signal. 2005, 7, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Immenschuh, S.; Baumgart-Vogt, E. Peroxiredoxins, oxidative stress, and cell proliferation. Antioxid. Redox Signal. 2005, 7, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Fountoulakis, M.; Cairns, N.; Lubec, G. Protein levels of human peroxiredoxin subtypes in brains of patients with Alzheimer’s disease and down syndrome. J. Neural Transm. Suppl. 2001, 223–235. [Google Scholar]
- Chang, R.; Wang, E. Mouse translation elongation factor eEF1A-2 interacts with Prdx-I to protect cells against apoptotic death induced by oxidative stress. J. Cell Biochem. 2007, 100, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Majd, S.; Power, J.H.T. Oxidative stress and decreased mitochondrial superoxide dismutase 2 and peroxiredoxins 1 and 4 based mechanism of concurrent activation of ampk and mtor in Alzheimer’s disease. Curr. Alzheimer Res. 2018, 15, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Schreibelt, G.; van Horssen, J.; Haseloff, R.F.; Reijerkerk, A.; van der Pol, S.M.; Nieuwenhuizen, O.; Krause, E.; Blasig, I.E.; Dijkstra, C.D.; Ronken, E.; et al. Protective effects of peroxiredoxin-1 at the injured blood-brain barrier. Free Radic. Biol. Med. 2008, 45, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.A.; Nelson, K.J.; Haynes, A.C.; Lee, J.; Reisz, J.A.; Graff, A.H.; Clodfelter, J.E.; Parsonage, D.; Poole, L.B.; Furdui, C.M.; et al. Novel hyperoxidation resistance motifs in 2-Cys peroxiredoxins. J. Biol. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pasban-Aliabadi, H.; Esmaeili-Mahani, S.; Abbasnejad, M. Orexin-A protects human neuroblastoma SH-SY5Y cells against 6-hydroxydopamine-induced neurotoxicity: Involvement of PKC and PI3K signaling pathways. Rejuvenation Res. 2017, 20, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Lee, B.D.; Sok, D.E.; Ma, J.Y.; Kim, M.R. Neuroprotective action of N-acetyl serotonin in oxidative stress-induced apoptosis through the activation of both TrkB/CREB/BNDF pathway and Akt/Nrf2/antioxidant enzyme in neuronal cells. Redox Biol. 2017, 11, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.L.; Durham, H.D.; Török, Z.; Crul, T.; Vígh, L. The central role of heat shock factor 1 in synaptic fidelity and memory consolidation. Cell Stress Chaperones 2016, 21, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Pfister, J.A.; Mallick, S.; D’Mello, S.R. HSF1 protects neurons through a novel trimerization- and HSP-independent mechanism. J. Neurosci. 2014, 34, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Ginet, V.; Sun, Y.; Koike, M.; Zhou, K.; Li, T.; Li, H.; Li, Q.; Wang, X.; Uchiyama, Y.; et al. Neuroprotection by selective neuronal deletion of Atg7 in neonatal brain injury. Autophagy 2016, 12, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Malaguti, M.; Rizzo, B.; Barbalace, M.C.; Fabbri, D.; Hrelia, S. Neuroprotective effect of sulforaphane against methylglyoxal cytotoxicity. Chem. Res. Toxicol. 2015, 28, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Motori, E.; Fabbri, D.; Malaguti, M.; Leoncini, E.; Lorenzini, A.; Hrelia, S. H2O2 preconditioning modulates phase ii enzymes through p38 MAPK and PI3K/Akt activation. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H2196–H2205. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Teti, G.; Barbalace, M.C.; Malaguti, M.; Falconi, M.; Hrelia, S. 17β-estradiol enhances sulforaphane cardioprotection against oxidative stress. J. Nutr. Biochem. 2017, 42, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Leoncini, E.; Malaguti, M.; Angelini, S.; Hrelia, P.; Hrelia, S. Role of quercetin in modulating rat cardiomyocyte gene expression profile. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1233–H1243. [Google Scholar] [CrossRef] [PubMed]
- Ciregia, F.; Bugliani, M.; Ronci, M.; Giusti, L.; Boldrini, C.; Mazzoni, M.R.; Mossuto, S.; Grano, F.; Cnop, M.; Marselli, L.; et al. Palmitate-induced lipotoxicity alters acetylation of multiple proteins in clonal β cells and human pancreatic islets. Sci. Rep. 2017, 7, 13445. [Google Scholar] [CrossRef] [PubMed]
- Ciregia, F.; Giusti, L.; da Valle, Y.; Donadio, E.; Consensi, A.; Giacomelli, C.; Sernissi, F.; Scarpellini, P.; Maggi, F.; Lucacchini, A.; et al. A multidisciplinary approach to study a couple of monozygotic twins discordant for the chronic fatigue syndrome: A focus on potential salivary biomarkers. J. Transl. Med. 2013, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- Ciregia, F.; Giusti, L.; Ronci, M.; Bugliani, M.; Piga, I.; Pieroni, L.; Rossi, C.; Marchetti, P.; Urbani, A.; Lucacchini, A. Glucagon-like peptide 1 protects ins-1e mitochondria against palmitate-mediated β-cell dysfunction: A proteomic study. Mol. Biosyst. 2015, 11, 1696–1707. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # Spot | Protein Name | Gene | ID | Peptide (n) | Unique | Coverage (%) | MW (th) | pI (th) |
|---|---|---|---|---|---|---|---|---|
| Metabolism | ||||||||
| 381 | Guanosine 5′-monophosphate synthase (glutamine-hydrolyzing) | GMPS | P49915 | 32 | 32 | 52 | 76,716 | 6.42 |
| 521 | Pyruvate kinase PKM | PKM | P14618 | 36 | 6 | 59 | 57,937 | 7.96 |
| 559 | Amidophosphoribosyltransferase | PPAT | Q06203 | 4 | 4 | 10 | 57,398 | 6.3 |
| 693 | Gamma-enolase | ENO2 | P09104 | 13 | 13 | 46 | 47,269 | 4.91 |
| 856 | Cytosolic acyl coenzyme A thioester hydrolase | ACOT7 | O00154 | 3 | 3 | 14 | 27,041 | 6.51 |
| 876 | Alcohol dehydrogenase (NADP(+)) | AKR1A1 | P14550 | 14 | 14 | 50 | 36,573 | 6.32 |
| 939 | Malate dehydrogenase, cytoplasmic | MDH1 | P40925 | 7 | 7 | 32 | 36,426 | 6.91 |
| 950 | ADP-sugar pyrophosphatase | NUDT5 | Q9UKK9 | 6 | 6 | 26 | 24,328 | 4.87 |
| 1069 | Enoyl-CoA hydratase, mitochondrial | ECHS1 | P30084 | 13 | 13 | 43 | 31,387 | 8.34 |
| 1076 | NADH:ubiquinone oxidoreductase iron-sulfur protein 3, mitocondrial | NDUFS3 | O75489 | 8 | 8 | 32 | 30,242 | 6.98 |
| 1098 | Triosephosphate isomerase | TPI1 | P60174 | 12 | 12 | 57 | 26,669 | 6.45 |
| 2010 | Phosphoglycerate kinase 1 | PGK1 | P00558 | 41 | 35 | 72 | 44,615 | 8.3 |
| 2010 | Isocitrate dehydrogenasemitochondrial isoform2 | IDH2 | P48735 | 25 | 25 | 53 | 45,180 | 7.63 |
| Protein synthesis, modification secretion folding | ||||||||
| 296 | Acylamino-acid-releasing enzyme | APEH | P13798 | 15 | 15 | 24 | 81,225 | 5.29 |
| 708 | Adenosylhomocysteinase | AHCY | P23526 | 18 | 18 | 37 | 47,716 | 5.92 |
| 916 | Small glutamine-rich tetratricopeptide repeat-containing protein alpha | SGTA | O43765 | 8 | 8 | 27 | 34,063 | 4.79 |
| 940 | Ubiquitin thioesterase OTUB1 | OTUB1 | Q96FW1 | 5 | 5 | 23 | 31,284 | 4.85 |
| 1230 | Protein DJ-1 | PARK7 | Q99497 | 11 | 11 | 49 | 19,891 | 6.32 |
| 1812 | Platelet-activating factor acetylhydrolase IB subunit gamma | PAFAH1B3 | Q15102 | 6 | 6 | 27 | 25,734 | 6.33 |
| Protein degradation | ||||||||
| 498 | Ubiquitin carboxyl-terminal hydrolase 14 | USP14 | P54578 | 13 | 13 | 35 | 52,386 | 5.61 |
| 995 | Proteasome activator complex subunit 3 | PSME3 | P61289 | 10 | 10 | 39 | 29,506 | 5.69 |
| 1111 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 | UCHL1 | P09936 | 29 | 29 | 81 | 24,824 | 5.33 |
| Cytoskeleton, vesicle motility, transport, vesicle release | ||||||||
| 430 | Dihydropyrimidinase-related protein 3 | DPYSL3 | Q14195 | 29 | 24 | 57 | 73,911 | 5.94 |
| 436 | Dihydropyrimidinase-related protein 3 | DPYSL3 | Q14195 | 30 | 30 | 58 | 73,911 | 5.94 |
| 441 | Dihydropyrimidinase-related protein 3 | DPYSL3 | Q14195 | 29 | 24 | 58 | 73,911 | 5.94 |
| 1079 | Ran-specific GTPase-activating protein | RANBP1 | P43487 | 9 | 9 | 50 | 23,310 | 5.19 |
| 1425 | Cofilin-1 | CFL1 | P23528 | 8 | 8 | 51 | 18,502 | 8.22 |
| 1743 | F-actin-capping protein subunit β | CAPZB | P47756 | 17 | 17 | 56 | 30,629 | 5.69 |
| RNA synthesis/metabolism | ||||||||
| 590 | ATP-dependent RNA helicase DDX39A | DDX39A | O00148 | 16 | 16 | 35 | 49,130 | 5.46 |
| 621 | Spliceosome RNA helicase DDX39B | DDX39B | Q13838 | 18 | 18 | 39 | 48,991 | 5.44 |
| 624 | Heterogeneous nuclear ribonucleoprotein H | HNRNPH1 | P31943 | 25 | 13 | 55 | 49,229 | 5.89 |
| 836 | Heterogeneous nuclear ribonucleoprotein A3 | HNRNPA3 | P51991 | 24 | 23 | 49 | 39,595 | 9.1 |
| 876 | Heterogeneous nuclear ribonucleoprotein H3 | HNRNPH3 | P31942 | 12 | 12 | 58 | 36,926 | 6.37 |
| 915 | Heterogeneous nuclear ribonucleoprotein H3 | HNRNPH3 | P31942 | 8 | 8 | 40 | 35,239 | 6.36 |
| 1735 | Heterogeneous nuclear ribonucleoprotein A1 | HNRNPA1 | P09651 | 15 | 11 | 63 | 29,386 | 9.19 |
| Chromatin remodeling and histone | ||||||||
| 498 | Poly(U)-binding-splicing factor PUF60 isoforma1 | PUF60 | Q9UHX1 | 10 | 10 | 28 | 59,875 | 5.19 |
| Stress response | ||||||||
| 181 | 26S proteasome non-ATPase regulatory subunit 1 | PSMD1 | Q99460 | 16 | 16 | 27 | 10,2258 | 5.14 |
| 334 | Heat shock protein HSP 90-α | HSP90AA1 | P07900 | 12 | 6 | 19 | 84,660 | 4.94 |
| 335 | Heat shock protein HSP 90-β | HSP90AB1 | P08238 | 15 | 8 | 24 | 83,264 | 4.96 |
| 498 | 60 kDa heat shock protein. mitochondrial | HSPD1 | P10809 | 31 | 31 | 54 | 61,055 | 5.7 |
| 506 | 60 kDa heat shock protein. mitochondrial | HSPD1 | P10809 | 54 | 54 | 73 | 61,055 | 5.7 |
| 1153 | 60 kDa heat shock protein. mitochondrial | HSPD1 | P10809 | 4 | 4 | 61,055 | 5.7 | |
| 701 | DnaJ homolog subfamily A member 2 | DNAJA2 | O60884 | 15 | 15 | 35 | 45,746 | 6.06 |
| 995 | Proteasome activator complex subunit 3 | PSME3 | P61289 | 10 | 10 | 39 | 29,506 | 5.69 |
| 1087 | Peroxiredoxin-6 | PRDX6 | P30041 | 21 | 21 | 65 | 25,035 | 6 |
| 1090 | Peroxiredoxin-6 | PRDX6 | P30041 | 20 | 20 | 67 | 25,035 | 6 |
| 1122 | Heat shock protein β-1 | HSPB1 | P04792 | 21 | 21 | 77 | 22,783 | 5.98 |
| 1253 | Peroxiredoxin-2 | PRDX2 | P32119 | 30 | 30 | 59 | 21,892 | 5.66 |
| 1269 | Peroxiredoxin-2 | PRDX2 | P32119 | 20 | 20 | 59 | 21,892 | 5.66 |
| 1725 | Peroxiredoxin-4 | PRDX4 | Q13162 | 4 | 4 | 17 | 30,540 | 5.86 |
| 1731 | Peroxiredoxin-1 | PRDX1 | Q06830 | 21 | 21 | 72 | 22,110 | 8.27 |
| 1736 | Peroxiredoxin-1 | PRDX1 | Q06830 | 17 | 17 | 72 | 22,110 | 8.27 |
| 1741 | Thioredoxin-dependent peroxide reductase. mitochondrial | PRDX3 | P30048 | 11 | 11 | 46 | 27,693 | 7.68 |
| 1743 | F-actin-capping protein subunit β | CAPZB | P47756 | 17 | 17 | 56 | 30,629 | 5.69 |
| Miscellaneous | ||||||||
| 498 | Ubiquitin carboxyl-terminal hydrolase 14 | USP14 | P54578 | 13 | 13 | 35 | 52,386 | 5.61 |
| 715 | Programmed cell death protein 2-like | PDCD2L | Q9BRP1 | 3 | 3 | 13 | 39,417 | 4.71 |
| 826 | Guanine nucleotide-binding protein G(i) subunit α-2 | GNAI2 | P04899 | 16 | 13 | 50 | 40,451 | 5.34 |
| 843 | V-type proton ATPase subunit d 1 | ATP6V0D1 | P61421 | 5 | 5 | 20 | 40,329 | 4.89 |
| 843 | Nucleophosmin | NPM1 | P06748 | 14 | 14 | 52 | 32,575 | 4.64 |
| 950 | Annexin A5 | ANXA5 | P08758 | 6 | 6 | 18 | 35,937 | 4.93 |
| 1153 | High mobility group protein B1 | HMGB1 | P09429 | 4 | 4 | 24 | 24,894 | 5.6 |
| 1733 | Chloride intracellular channel protein 1 | CLIC1 | O00299 | 16 | 15 | 61 | 26,923 | 5.09 |
| 1743 | N(G).N(G)-dimethylarginine dimethylaminohydrolase 2 | DDAH2 | O95865 | 13 | 13 | 54 | 29,644 | 5.66 |
| 1767 | MICOS complex subunit MIC60 | IMMT | Q16891 | 41 | 41 | 60 | 82625 | 6.15 |
| 1813 | 28 kDa heat- and acid-stable phosphoprotein | PDAP1 | Q13442 | 3 | 3 | 13 | 20,630 | 8.84 |
| # Spot | ID | Gene | Protein Name | Ratio (OD) H2O2/Ctrl | p-Value |
|---|---|---|---|---|---|
| 181 | Q99460 | PSMD1 | 26S proteasome non-ATPase regulatory subunit 1 | 0.76 | 0.026 |
| 335 | P08238 | HSP90AB1 | Heat shock protein HSP 90-beta | 0.67 | 0.018 |
| 381 | P49915 | GMPS | GMP synthase [glutamine-hydrolyzing] | 0.75 | 0.014 |
| 430 | Q14195 | DPYSL3 | Dihydropyrimidinase-related protein 3 | 0.58 | 0.002 |
| 436 | Q14195 | DPYSL3 | Dihydropyrimidinase-related protein 3 | 0.74 | 0.001 |
| 441 | Q14195 | DPYSL3 | Dihydropyrimidinase-related protein 3 | 1.56 | 0.022 |
| 506 | P10809 | HSPD1 | 60 kDa heat shock protein. mitochondrial | 0.83 | 0.011 |
| 559 | Q06203 | PPAT | Amidophosphoribosyltransferase | 0.57 | 0.046 |
| 621 | Q13838 | DDX39B | Spliceosome RNA helicase DDX39B | 0.83 | 0.022 |
| 624 | P31943 | HNRNPH1 | Heterogeneous nuclear ribonucleoprotein H | 0.81 | 0.039 |
| 701 | DNAJA2 | O60884 | DnaJ homolog subfamily A member 2 | 0.77 | 0.017 |
| 708 | P23526 | AHCY | Adenosylhomocysteinase | 0.82 | 0.012 |
| 856 | O00154 | ACOT7 | Cytosolic acyl coenzyme A thioester hydrolase | 0.76 | 0.036 |
| 876 | P14550 | AKR1A1 | Alcohol dehydrogenase (NADP(+)) | 0.69 | 0.043 |
| 876 | P31942 | HNRNPH3 | Heterogeneous nuclear ribonucleoprotein H3 | ||
| 915 | P31942 | HNRNPH3 | Heterogeneous nuclear ribonucleoprotein H3 | 0.71 | 0.026 |
| 939 | P40925 | MDH1 | Malate dehydrogenase. cytoplasmic | 0.68 | 0.026 |
| 940 | Q96FW1 | OTUB1 | Ubiquitin thioesterase OTUB1 | 0.73 | 0.017 |
| 950 | P08758 | ANXA5 | Annexin A5 | 0.78 | 0.012 |
| 950 | Q9UKK9 | NUDT5 | ADP-sugar pyrophosphatase | ||
| 1069 | P30084 | ECHS1 | Enoyl-CoA hydratase. mitochondrial | 0.78 | 0.023 |
| 1079 | P43487 | RANBP1 | Ran-specific GTPase-activating protein | 0.80 | 0.035 |
| 1087 | P30041 | PRDX6 | Peroxiredoxin-6 | 5.84 | 0.007 |
| 1090 | P30041 | PRDX6 | Peroxiredoxin-6 | 0.53 | 0.032 |
| 1098 | P60174 | TPI1 | Triosephosphate isomerase | 0.68 | 0.004 |
| 1122 | P04792 | HSPB1 | Heat shock protein β-1 | 0.69 | 0.015 |
| 1179 | P28070 | PSMB4 | Proteasome subunit β type-4 | 0.80 | 0.036 |
| 1253 | P32119 | PRDX2 | Peroxiredoxin-2 | 0.17 | 4.4 × 10−7 |
| 1269 | P32119 | PRDX2 | Peroxiredoxin-2 | 2.27 | 3.08 × 10−5 |
| 1725 | Q13162 | PRDX4 | Peroxiredoxin-4 | 0.62 | 0.016 |
| 1731 | Q06830 | PRDX1 | Peroxiredoxin-1 | 0.33 | 4.28 × 10−6 |
| 1736 | Q06830 | PRDX1 | Peroxiredoxin-1 | 3.37 | 1.37 × 10−6 |
| 1741 | P30048 | PRDX3 | Thioredoxin-dependent peroxide reductase mitochondrial | 0.48 | 0.00041 |
| 1743 | P47756 | CAPZB | F-actin-capping protein subunit β | 0.80 | 0.042 |
| 1743 | O95865 | DDAH2 | N(G).N(G)-dimethylarginine dimethylaminohydrolase 2 | ||
| 1767 | Q16891 | IMMT | MICOS complex subunit MIC60 | 0.72 | 0.021 |
| 1812 | Q15102 | PAFAH1B3 | Platelet-activating factor acetylhydrolase IB subunit γ | 0.76 | 0.021 |
| # Spot | ID | Gene | Protein Name | Ratio M ± SD (OD) OC + H2O2/H2O2 | p-Value |
|---|---|---|---|---|---|
| 181 | Q99460 | PSMD1 | 26S proteasome non-ATPase regulatory subunit 1 | 1.29 | 0.034 |
| 296 | P13798 | APEH | Acylamino-acid-releasing enzyme | 1.28 | 0.009 |
| 334 | P08238 | HSP90AB1 | Heat shock protein HSP 90-β | 1.33 | 0.026 |
| 334 | P07900 | HSP90AA1 | Heat shock protein HSP 90-alpha | ||
| 498 | P54578 | USP14 | Ubiquitin carboxyl-terminal hydrolase 14 | 1.30 | 0.023 |
| 498 | Q9UHX1 | PUF60 | Poly(U)-binding-splicing factor PUF60 isoforma1 | ||
| 521 | P14618 | PKM | Pyruvate kinase PKM | 1.35 | 0.035 |
| 590 | O00148 | DDX39A | ATP-dependent RNA helicase DDX39A | 1.36 | 0.018 |
| 621 | Q13838 | DDX39B | Spliceosome RNA helicase DDX39B | 1.29 | 0.013 |
| 715 | Q9BRP1 | PDCD2L | Programmed cell death protein 2-like | 0.77 | 0.013 |
| 836 | P51991 | HNRNPA3 | Heterogeneous nuclear ribonucleoprotein A3 | 0.64 | 0.023 |
| 843 | P61421 | ATP6V0D1 | V-type proton ATPase subunit d 1 | 0.64 | 0.019 |
| 843 | P06748 | NPM1 | Nucleophosmin | ||
| 940 | Q96FW1 | OTUB1 | Ubiquitin thioesterase OTUB1 | 1.45 | 0.036 |
| 1109 | P21266 | GSTM3 | Glutathione S-transferase Mu 3 | 1.23 | 0.036 |
| 1111 | P09936 | UCHL1 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 | 1.25 | 0.002 |
| 1153 | P09429 | HMGB1 | High mobility group protein B1 | 0.43 | 0.049 |
| 1179 | P28070 | PSMB4 | Proteasome subunit β type-4 | 1.30 | 0.024 |
| 1736 | Q06830 | PRDX1 | Peroxiredoxin-1 | 1.21 | 0.029 |
| Master Regulator | Molecule Type | Predicted Activation State | Activation z-Score | p-Value |
|---|---|---|---|---|
| GPR68 | G-protein complex | Inhibited | −2.12 | 1.09 × 10−4 |
| MARK2 | Kinase | Inhibited | −2.33 | 2.67 × 10−4 |
| miR-149-5p | Mature microRNA | Inhibited | −2.53 | 1.36 × 10−4 |
| ATG7 | Enzyme | Inhibited | −2.65 | 7.88 × 10−6 |
| Smad2/3-Smad4 | Complex | Inhibited | −2.83 | 3.02 × 10−4 |
| PDE2A | Enzyme | Activated | +2.53 | 4.07 × 10−5 |
| ELMO1 | Other | Activated | +2.53 | 8.06 × 10−5 |
| DOCK5 | Other | Activated | +2.53 | 8.14 × 10−5 |
| FARP2 | Other | Activated | +2.53 | 1.07 × 10−4 |
| ARHGEF6/7 | Group | Activated | +2.53 | 1.07 × 10−5 |
| BAIAP2 | Kinase | Activated | +2.33 | 2.44 × 10−4 |
| RAC1 | Enzyme | Activated | +2.33 | 2.47 × 10−4 |
| Hif | Complex | Activated | +2.33 | 3.28 × 10−4 |
| ADORA1 | G-protein complex | Activated | +2.33 | 4.12 × 10−4 |
| NOX4 | Enzyme | Activated | +2.31 | 3.59 × 10−6 |
| NFE2L2 | Transcription factor | Activated | +2.23 | 1.07 × 10−5 |
| MYCN | Transcription factor | Activated | +2.00 | 3.85 × 10−5 |
| HSF1 | Transcription factor | Activated | +2.00 | 1.33 × 10−3 |
| Gene | 5′-Forward-3′ | 5′-Reverse-3′ |
|---|---|---|
| GSTM3 | TTGAGGCTTTGGAGAAAATC | TGAAAAGAGCAAAGCAAGAG |
| HSP90AA1 | ATATCACAGGTGAGACCAAG | GTGACTGACACTAAAGTCTTC |
| HSP90AB1 | TCTATTACATCACTGGTGAGAG | CTCTTCCCATCAAATTCCTTG |
| PSMB4 | ACGCTGATTTCCAGTATTTG | CCATGAATGAATAGCTCTAGG |
| PSMD1 | CCAGTTATTGGATAACCCAG | CTCCAATAGAGAGTGGTTTG |
| USP14 | AGCCCTTAGAGATTTGTTTG | ATCCTGTTGAAGATACTGTCC |
| PKM1 | GAGGCAGCCATGTTCCAC | TGCCAGACTCCGTCAGAACT |
| PKM2 | CAGAGGCTGCCATCTACCAC | CCAGACTTGGTGAGGACGAT |
| PRDX1 | GGGTCAATACACCTAAGAAAC | CTTCATCAGCCTTTAAGACC |
| 18S rRNA | CAGAAGGATGTAAAGGATGG | TATTTCTTCTTGGACACACC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giusti, L.; Angeloni, C.; Barbalace, M.C.; Lacerenza, S.; Ciregia, F.; Ronci, M.; Urbani, A.; Manera, C.; Digiacomo, M.; Macchia, M.; et al. A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 2329. https://doi.org/10.3390/ijms19082329
Giusti L, Angeloni C, Barbalace MC, Lacerenza S, Ciregia F, Ronci M, Urbani A, Manera C, Digiacomo M, Macchia M, et al. A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress. International Journal of Molecular Sciences. 2018; 19(8):2329. https://doi.org/10.3390/ijms19082329
Chicago/Turabian StyleGiusti, Laura, Cristina Angeloni, Maria Cristina Barbalace, Serena Lacerenza, Federica Ciregia, Maurizio Ronci, Andrea Urbani, Clementina Manera, Maria Digiacomo, Marco Macchia, and et al. 2018. "A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress" International Journal of Molecular Sciences 19, no. 8: 2329. https://doi.org/10.3390/ijms19082329
APA StyleGiusti, L., Angeloni, C., Barbalace, M. C., Lacerenza, S., Ciregia, F., Ronci, M., Urbani, A., Manera, C., Digiacomo, M., Macchia, M., Mazzoni, M. R., Lucacchini, A., & Hrelia, S. (2018). A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress. International Journal of Molecular Sciences, 19(8), 2329. https://doi.org/10.3390/ijms19082329