Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses



Abstract
1. Introduction
2. Results
2.1. Identification of WRKY Gene Family in Longan
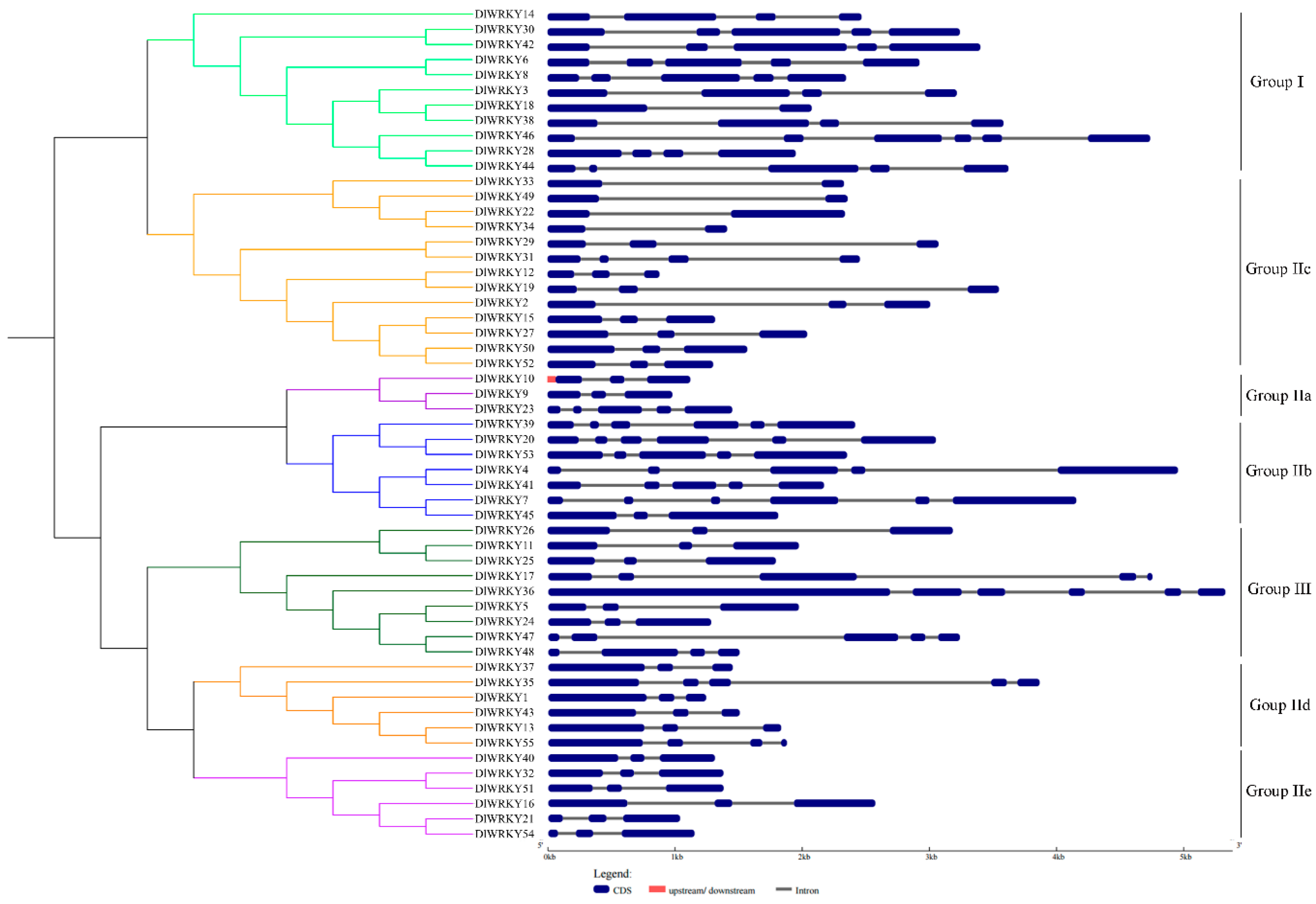
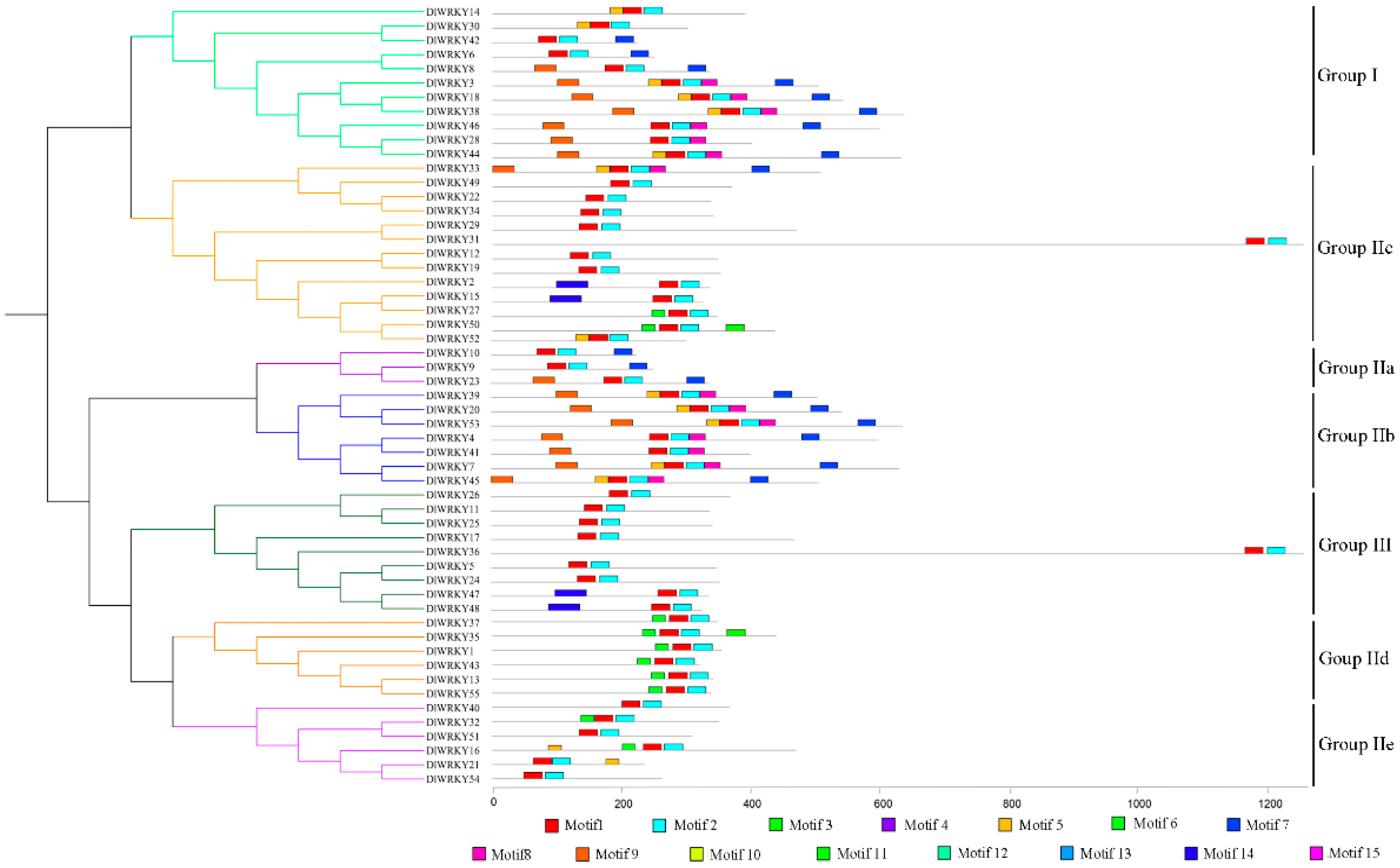
2.2. Phylogenetic Analysis of DlWRKY
2.3. Multiple Sequence Alignment and Structure Analysis
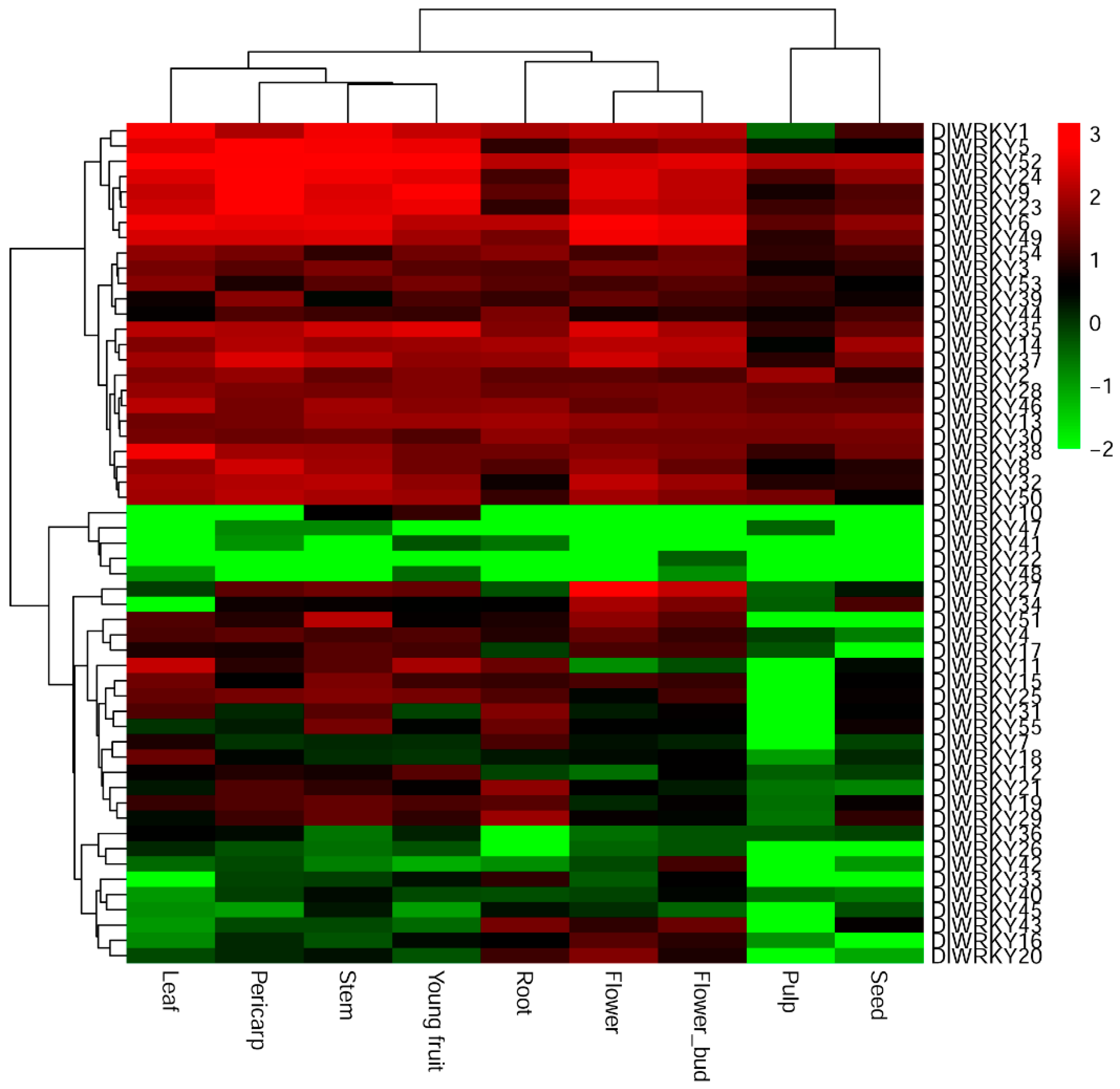
2.4. Tissue-Specific Expression Patterns of DlWRKY
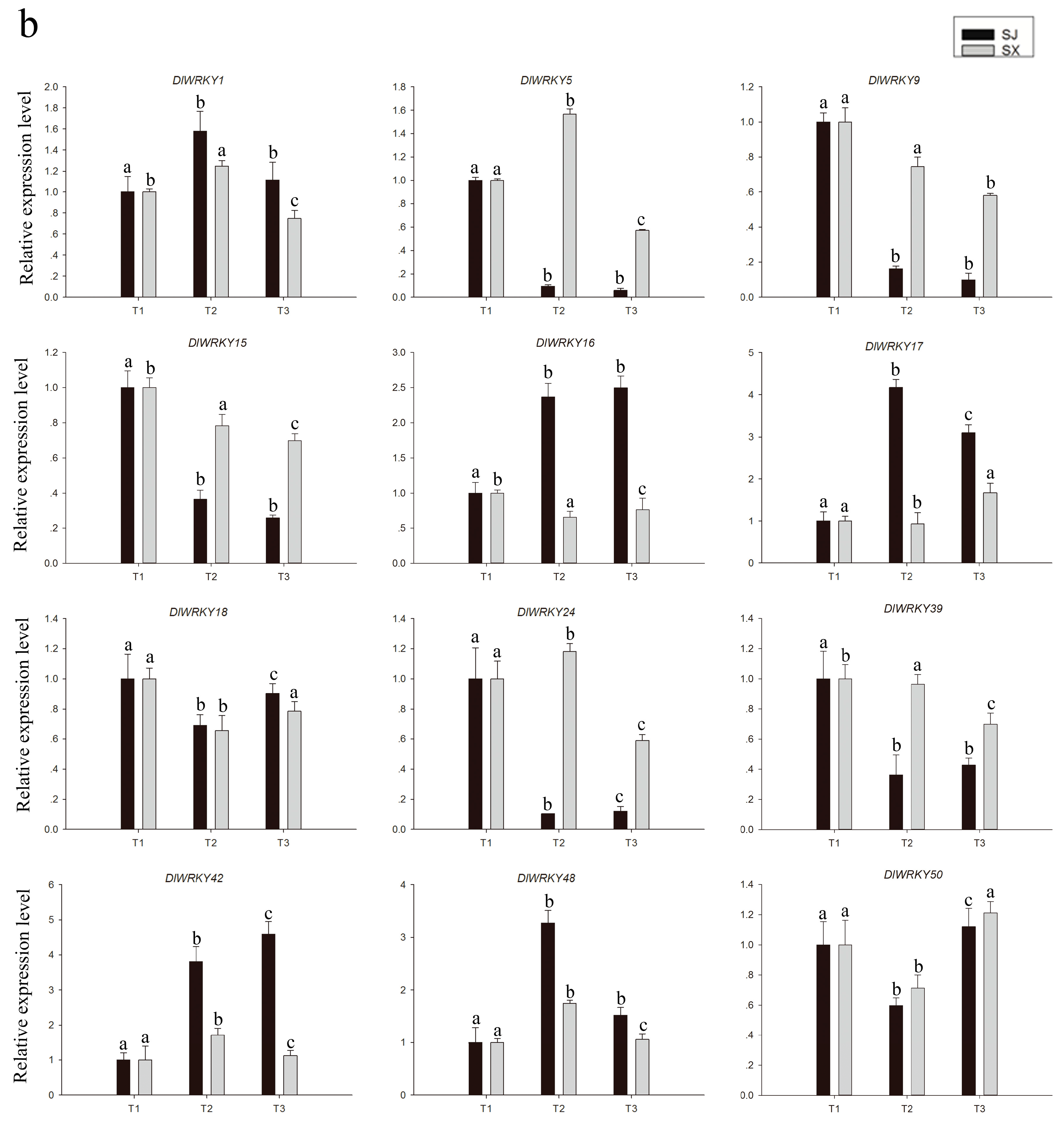
2.5. Comparative Expression Profiles of Two Longan Species during the Flowering Process
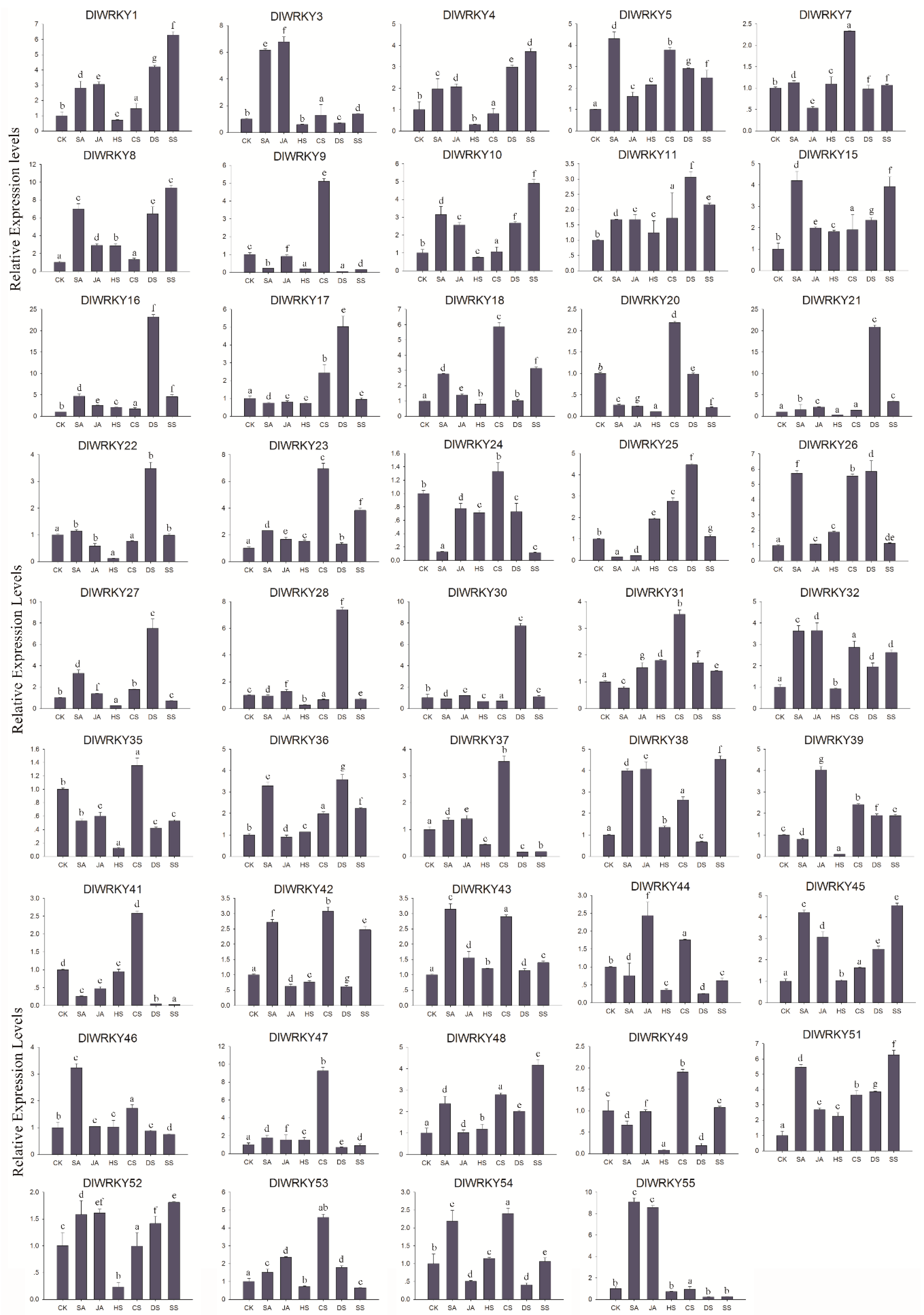
2.6. Differential Expression of DlWRKY Genes in Response to Stress and Hormonal Treatments
2.7. Analysis Related Cis-Elements in the Candidate DlWRKY Genes
3. Discussion
4. Materials
4.1. Identification of Longan WRKY Genes
4.2. Sequence Alignment, Phylogenetic Analysis, and Cis-Elements in the Promoters
4.3. Protein Feature Analysis
4.4. Expression Analysis of Longan WRKY Genes in Various Tissues and Different Flowering Stages
4.5. Stress and Hormonal Treatments and Expression Profiling Using qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviation
| HMM | Hidden Markov model |
| NJ | Neighbor-joining |
| GSDS | Gene Structure Display Server |
| MW | The molecular weight |
| ORF | Open reading frame |
| pI | Isoelectric point |
| NCBI | National Center of Biotechnology Information |
| qRT-PCR | Quantitative real-time reverse transcription polymerase chain reaction |
| RNA-seq | RNA sequencing |
| SA | Salicylic acid |
| JA | Jasmonic acid |
References
- Matsumoto, T.K. Genes uniquely expressed in vegetative and potassium chlorate induced floral buds of Dimocarpus longan. Plant Sci. 2006, 170, 500–510. [Google Scholar] [CrossRef]
- You, X.; Wang, L.; Liang, W.; Gai, Y.; Wang, X.; Chen, W. Floral reversion mechanism in longan (Dimocarpus longan lour.) revealed by proteomic and anatomic analyses. J. Proteom. 2012, 75, 1099–1118. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Wei, D.; Meng, S.; Allan, A.C.; Zeng, L. Identification of regulatory genes implicated in continuous flowering of longan (Dimocarpus longan L.). PLoS ONE 2014, 9, e114568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.N.; Shi, S.Y.; Li, W.C.; Shu, B.; Liu, L.Q.; Xie, J.H.; Wei, Y.Z. Transcriptome analysis of ‘sijihua’ longan (Dimocarpus longan L.) based on next-generation sequencing technology. J. Hortic. Sci. Biotechnol. 2016, 91, 180–188. [Google Scholar] [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Bluemel, M.; Dally, N.; Jung, C. Flowering time regulation in crops-what did we learn from arabidopsis? Curr. Opin. Biotechnol. 2015, 32, 121–129. [Google Scholar]
- Dally, N.; Xiao, K.; Holtgräwe, D.; Jung, C. The b2 flowering time locus of beet encodes a zinc finger transcription factor. Proc. Natl. Acad. Sci. USA 2014, 111, 10365–10370. [Google Scholar] [CrossRef] [PubMed]
- Andrés, F.; Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 2012, 13, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, C. Long-distance regulation of flowering time. J. Exp. Bot. 2011, 62, 4399–4413. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Smaczniak, C.; Immink, R.G.; Muiño, J.M.; Blanvillain, R.; Busscher, M.; Busscher-Lange, J.; Dinh, Q.P.; Liu, S.; Westphal, A.H.; Boeren, S. Characterization of mads-domain transcription factor complexes in arabidopsis flower development. Proc. Natl. Acad. Sci. USA 2012, 109, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Y.; Kim, Y.; Kim, S.Y.; Lee, J.S.; Ahn, J.H. Control of flowering time and cold response by a nac-domain protein in arabidopsis. PLoS ONE 2007, 2, e642. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.; Choi, G.; Yi, H.; Yang, S.; Cho, I.; Kim, J.; Lee, S.; Paek, N.C.; Kim, J.H.; Song, P.S. Atmyb21, a gene encoding a flower-specific transcription factor, is regulated by cop1. Plant J. 2002, 30, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Hong, B.; Yang, Y.; Li, Q.; Ma, N.; Ma, C.; Gao, J. Overexpression of two chrysanthemum dgdreb1 group genes causing delayed flowering or dwarfism in arabidopsis. Plant Mol. Biol. 2009, 71, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Hu, Y.; Vannozzi, A.; Wu, K.; Cai, H.; Qin, Y.; Mullis, A.; Lin, Z.; Zhang, L. The wrky transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 2017, 36, 311–335. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.; Wang, L.; Kim, S.G.; Seo, P.J.; Qiao, M.; Wang, N.; Li, S.; Cao, X.; Park, C.M. Wrky71 accelerates flowering via the direct activation of flowering locus t and leafy in arabidopsis thaliana. Plant J. 2015, 85, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Chen, X.; Xie, K.; Xing, Q.; Wu, Y.; Li, J.; Du, C.; Sun, Z.; Guo, Z. Dlf1, a wrky transcription factor, is involved in the control of flowering time and plant height in rice. PLoS ONE 2014, 9, e102529. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hu, R.; Wang, H.; Cao, Y.; He, G.; Fu, C.; Zhou, G. Mlwrky12, a novel miscanthus transcription factor, participates in pith secondary cell wall formation and promotes flowering. Plant Sci. 2013, 212, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, H.; Yu, D. The arabidopsis wrky transcription factors wrky12 and wrky13 oppositely regulate flowering under short-day conditions. Mol. Plant 2016, 9, 1492–1503. [Google Scholar] [CrossRef] [PubMed]
- Kiranmai, K.; Lokanadha Rao, G.; Pandurangaiah, M.; Nareshkumar, A.; Amaranatha Reddy, V.; Lokesh, U.; Venkatesh, B.; Anthony Johnson, A.M.; Sudhakar, C. A novel wrky transcription factor, muwrky3 (Macrotyloma uniflorum lam. Verdc.) enhances drought stress tolerance in transgenic groundnut (Arachis hypogaea L.) plants. Front. Plant Sci. 2018, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fu, Q.; Chen, L.; Huang, W.; Yu, D. Arabidopsis thaliana wrky25, wrky26, and wrky33 coordinate induction of plant thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Lai, Z.; Shi, J.; Yong, X.; Chen, Z.; Xu, X. Roles of arabidopsis wrky18, wrky40 and wrky60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biol. 2010, 10, 281. [Google Scholar]
- Kilian, J.; Whitehead, D.J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, C.; Bornberg-Bauer, E.; Kudla, J.; Harter, K. The atgenexpress global stress expression data set: Protocols, evaluation and model data analysis of uv-b light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T. Monitoring the expression profiles of 7000 arabidopsis genes under drought, cold and high-salinity stresses using a full-length cdna microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Jing, S.; Fu, J.; Li, L.; Yu, D. Cloning and analysis of expression profile of 13 wrky genes in rice. Sci. Bull. 2004, 49, 2159–2168. [Google Scholar] [CrossRef]
- Raineri, J.; Wang, S.; Peleg, Z.; Blumwald, E.; Chan, R.L. The rice transcription factor oswrky47 is a positive regulator of the response to water deficit stress. Plant Mol. Biol. 2015, 88, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Baranwal, V.K.; Negi, N.; Khurana, P. Genome-wide identification and structural, functional and evolutionary analysis of wrky components of mulberry. Sci. Rep. 2016, 6, 30794. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, J.; Wang, L.; Wang, S. Genome-wide investigation of wrky transcription factors involved in terminal drought stress response in common bean. Front. Plant Sci. 2017, 8, 380. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) wrky gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of wrky gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2018, 19, 490. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, Y.; Chi, Y.; Fan, B.; Chen, Z. Characterization of soybean wrky gene family and identification of soybean wrky genes that promote resistance to soybean cyst nematode. Sci. Rep. 2017, 7, 17804. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mu, S.; Cheng, Z.; Cheng, Y.; Zhang, Y.; Miao, Y.; Hou, C.; Li, X.; Gao, J. Characterization and expression analysis of the wrky gene family in moso bamboo. Sci. Rep. 2017, 7, 6675. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Mao, M.; Wan, D.; Yang, Q.; Yang, F.; Mandlaa; Li, G.; Wang, R. Identification of thewrkygene family and functional analysis of two genes in caragana intermedia. BMC Plant Biol. 2018, 18, 31. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, P.; Lin, J.Y.; Zhao, C.; Bi, Y.; Wang, X. Genome-wide identification and characterization ofwrkygene family in peanut. Front. Plant Sci. 2016, 7, 534. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Wang, M.; Liu, S.; Du, X.; Song, W.; Nie, X. Transcriptome-wide identification and expression profiles of the wrky transcription factor family in broomcorn millet (Panicum miliaceum L.). BMC Genom. 2016, 17, 343. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Park, Y.H.; Bae, H. Novel genomic and evolutionary insight of wrky transcription factors in plant lineage. Sci. Rep. 2016, 6, 37309. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Min, J.; Lai, R.; Wu, Z.; Chen, Y.; Yu, L.; Cheng, C.; Jin, Y.; Tian, Q.; Liu, Q. Genome-wide sequencing of longan (Dimocarpus longan lour.) provides insights into molecular basis of its polyphenol-rich characteristics. Gigascience 2017, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Mao, Z.; Yu, H.; Zhang, Y.; Jiang, W.; Ling, J.; Huang, S.; Xie, B. Genome-wide analysis of wrky gene family in cucumis sativus. BMC Genom. 2011, 12, 471. [Google Scholar]
- Yu, J.; Yang, H. A draft sequence of the rice genome (Oryza sativa L. ssp. Indica). Science 2002, 296, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, P.; Yu, J.; Wang, L.; Dossa, K.; Zhang, Y.; Zhou, R.; Wei, X.; Zhang, X. Genome-wide analysis of wrky gene family in the sesame genome and identification of the wrky genes involved in responses to abiotic stresses. BMC Plant Biol. 2017, 17, 152. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The wrky superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Shuai, L.; Luo, C.; Zhu, L.; Sha, R.; Qu, S.; Cai, B.; Wang, S. Identification and expression analysis of wrky transcription factor genes in response to fungal pathogen and hormone treatments in apple (Malus domestica). J. Plant Biol. 2017, 60, 215–230. [Google Scholar]
- Zhi, Z.; Yang, L.; Wang, D.; Huang, Q.; Mo, Y.; Xie, G. Gene structures, evolution and transcriptional profiling of the wrky gene family in castor bean (Ricinus communis L.). PLoS ONE 2016, 11, e0148243. [Google Scholar]
- Cheng, Y.; Jalalahammed, G.; Yu, J.; Yao, Z.; Ruan, M.; Ye, Q.; Li, Z.; Wang, R.; Feng, K.; Zhou, G. Putative wrkys associated with regulation of fruit ripening revealed by detailed expression analysis of the wrky gene family in pepper. Sci. Rep. 2016, 6, 39000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L. The wrky transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evolut. Biol. 2005, 5, 1. [Google Scholar]
- Rishmawi, L.; Hülskamp, M. Non-cell-autonomous regulation of root hair patterning genes by wrky75 in arabidopsis thaliana. Plant Physiol. 2014, 165, 186. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Palmqvist, S.; Olsson, H.; Borén, M.; Ahlandsberg, S.; Jansson, C. A novel wrky transcription factor, susiba2, participates in sugar signaling in barley by binding to the sugar-responsive elements of the iso1 promoter. Plant Cell 2003, 15, 2076–2092. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. Miniseed3 (mini3), a wrky family gene, and haiku2 (iku2), a leucine-rich repeat (lrr) kinase gene, are regulators of seed size in arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, H.; Yang, Y.; Wang, Y.; Mo, Y.; Zhang, R.; Zhang, Y.; Ma, J.; Wei, C.; Zhang, X. Identification and expression analyses ofwrkygenes reveal their involvement in growth and abiotic stress response in watermelon (Citrullus lanatus). PLoS ONE 2018, 13, e0191308. [Google Scholar]
- Iwata, H.; Gaston, A.; Remay, A.; Thouroude, T.; Jeauffre, J.; Kawamura, K.; Oyant, L.H.S.; Araki, T.; Denoyes, B.; Foucher, F. The tfl1 homologue ksn is a regulator of continuous flowering in rose and strawberry. Plant J. Cell Mol. Biol. 2012, 69, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C. Pep1 regulates perennial flowering in Arabis alpina. Nature 2009, 459, 423–427. [Google Scholar]
- Koskela, E.A.; Hytönen, T. Mutation in terminal flower1 reverses the photoperiodic requirement for flowering in the wild strawberry fragaria vesca. Plant Physiol. 2012, 159, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Perrotte, J.; Gaston, A.; Potier, A.; Petit, A.; Rothan, C.; Denoyes, B. Narrowing down the single homoeologous fapfru locus controlling flowering in cultivated octoploid strawberry using a selective mapping strategy. Plant Biotechnol. J. 2016, 14, 2176–2189. [Google Scholar] [CrossRef] [PubMed]
- Gaston, A.; Perrotte, J.; Lerceteauköhler, E.; Rousseaugueutin, M.; Petit, A.; Hernould, M.; Rothan, C.; Denoyes, B. Pfru, a single dominant locus regulates the balance between sexual and asexual plant reproduction in cultivated strawberry. J. Exp. Bot. 2013, 64, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Besseau, S.; Li, J.; Palva, E.T. Wrky54 and wrky70 co-operate as negative regulators of leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 2667–2679. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Palva, E.T. The wrky70 transcription factor: A node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense. Plant Cell 2004, 16, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Deyholos, M.K. Functional characterization of arabidopsis nacl-inducible wrky25 and wrky33 transcription factors in abiotic stresses. Plant Mol. Biol. 2009, 69, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Tate, J.; Coggill, P.; Heger, A.; Pollington, J.E.; Gavin, O.L.; Gunasekaran, P.; Ceric, G.; Forslund, K. The pfam protein families database. Nucleic Acids Res. 2010, 38, D211–D222. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. Smart 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evolut. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. Plantcare, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. Expasy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C. Gsds: A gene structure display server. Hereditas 2007, 29, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. Meme suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Xie, J.; Liu, C.; Shi, S. The ubiquitin-conjugating enzyme gene family in longan (Dimocarpus longan lour.): Genome-wide identification and gene expression during flower induction and abiotic stress responses. Molecules 2018, 23, 662. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M. Tm4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Locus ID | Location | ORF (bp) | Size (aa) | PI | MW (KDa) | Intron | Full Length |
|---|---|---|---|---|---|---|---|---|
| DlWRKY1 | Dlo_000299.1 | scaffold1:3145979:3147233 | 1071 | 356 | 9.63 | 38.76 | 2 | 1255 |
| DlWRKY2 | Dlo_026119.1 | scaffold6:875263:878308 | 894 | 297 | 6.26 | 32.31 | 2 | 3046 |
| DlWRKY3 | Dlo_026149.1 | scaffold6:1127159:1130416 | 1596 | 532 | 7.26 | 57.64 | 3 | 3258 |
| DlWRKY4 | Dlo_026267.1 | scaffold6:2195842:2200859 | 1815 | 605 | 6.66 | 66.08 | 4 | 5018 |
| DlWRKY5 | Dlo_030713.1 | scaffold8:184175:186167 | 1059 | 353 | 5.63 | 39.46 | 2 | 1993 |
| DlWRKY6 | Dlo_002181.1 | scaffold11:1861336:1864296 | 1767 | 589 | 7.23 | 64.37 | 4 | 2961 |
| DlWRKY7 | Dlo_012455.1 | scaffold23:1107291:1111499 | 1914 | 638 | 6.75 | 69.01 | 5 | 4209 |
| DlWRKY8 | Dlo_013053.2 | scaffold24:1070557:1072933 | 1668 | 556 | 6.52 | 61.46 | 4 | 2377 |
| DlWRKY9 | Dlo_015501.2 | scaffold29:1782026:1783019 | 762 | 254 | 8.99 | 28.30 | 4 | 994 |
| DlWRKY10 | dlo_037126.1 | scaffold29:1793158:1794294 | 684 | 228 | 9.02 | 25.58 | 2 | 1146 |
| DlWRKY11 | Dlo_016404.1 | scaffold31:1522675:1524675 | 1026 | 342 | 5.60 | 38.86 | 2 | 2001 |
| DlWRKY12 | Dlo_019125.1 | scaffold38:1882835:1883726 | 480 | 160 | 5.16 | 18.38 | 2 | 892 |
| DlWRKY13 | Dlo_023965.1 | scaffold53:1206068:1207919 | 1035 | 345 | 9.77 | 38.51 | 2 | 1852 |
| DlWRKY14 | Dlo_028963.1 | scaffold71:878665:881164 | 1613 | 471 | 8.87 | 51.80 | 3 | 2500 |
| DlWRKY15 | Dlo_031097.1 | scaffold81:147303:148636 | 972 | 324 | 6.33 | 35.30 | 2 | 1334 |
| DlWRKY16 | Dlo_033905.1 | scaffold98:272537:275137 | 1419 | 473 | 5.82 | 51.20 | 2 | 2601 |
| DlWRKY17 | Dlo_001368.1 | scaffold105:274029:278833 | 1425 | 475 | 6.10 | 52.14 | 4 | 4805 |
| DlWRKY18 | Dlo_003898.1 | scaffold124:605265:607367 | 1053 | 351 | 9.04 | 39.34 | 1 | 2103 |
| DlWRKY19 | Dlo_003928.1 | scaffold124:1058067:1061659 | 633 | 211 | 6.37 | 23.26 | 2 | 3593 |
| DlWRKY20 | Dlo_004435.1 | scaffold129:429868:432959 | 1644 | 548 | 7.41 | 59.78 | 5 | 3092 |
| DlWRKY21 | Dlo_008095.1 | scaffold167:682922:683969 | 714 | 238 | 5.14 | 26.59 | 2 | 1048 |
| DlWRKY22 | Dlo_008126.1 | scaffold168:307774:310141 | 1245 | 415 | 4.62 | 44.95 | 1 | 2368 |
| DlWRKY23 | Dlo_008610.1 | scaffold176:75022:76492 | 1023 | 341 | 8.62 | 38.08 | 4 | 1471 |
| DlWRKY24 | Dlo_009865.1 | scaffold192:233555:234849 | 1071 | 357 | 5.50 | 39.25 | 2 | 1295 |
| DlWRKY25 | Dlo_011410.1 | scaffold213:248908:250725 | 1038 | 346 | 5.93 | 38.65 | 2 | 1818 |
| DlWRKY26 | Dlo_011411.1 | scaffold213:253855:257080 | 1122 | 374 | 6.00 | 40.22 | 2 | 3226 |
| DlWRKY27 | Dlo_012276.1 | scaffold229:13116:15182 | 1005 | 335 | 7.16 | 37.09 | 2 | 2067 |
| DlWRKY28 | Dlo_012878.1 | scaffold238:352167:354143 | 1527 | 509 | 5.89 | 55.49 | 3 | 1977 |
| DlWRKY29 | Dlo_013340.1 | scaffold245:258019:261130 | 696 | 232 | 8.95 | 26.57 | 2 | 3112 |
| DlWRKY30 | Dlo_013413.1 | scaffold247:267246:270528 | 2238 | 746 | 5.59 | 80.60 | 4 | 3283 |
| DlWRKY31 | Dlo_014324.1 | scaffold266:341214:343700 | 663 | 221 | 7.71 | 25.38 | 3 | 2487 |
| DlWRKY32 | Dlo_015139.1 | scaffold286:162902:164294 | 1059 | 353 | 6.32 | 38.46 | 2 | 1393 |
| DlWRKY33 | Dlo_015144.1 | scaffold286:195837:198196 | 615 | 205 | 9.03 | 23.13 | 1 | 2360 |
| DlWRKY34 | Dlo_015224.1 | scaffold287:217068:218497 | 480 | 160 | 9.54 | 18.10 | 1 | 1430 |
| DlWRKY35 | Dlo_016828.1 | scaffold322:63655:67562 | 1326 | 442 | 9.62 | 48.27 | 4 | 3908 |
| DlWRKY36 | Dlo_022548.1 | scaffold487:170363:175747 | 3813 | 1271 | 5.15 | 143.77 | 5 | 5385 |
| DlWRKY37 | Dlo_023098.1 | scaffold502:191885:193351 | 1056 | 352 | 9.46 | 38.46 | 2 | 1467 |
| DlWRKY38 | Dlo_023764.1 | scaffold524:170088:173717 | 1533 | 511 | 8.66 | 55.75 | 3 | 3630 |
| DlWRKY39 | Dlo_025188.1 | scaffold568:191129:193577 | 1530 | 510 | 8.26 | 55.75 | 5 | 2449 |
| DlWRKY40 | Dlo_025974.1 | scaffold597:89062:90386 | 1110 | 370 | 5.07 | 40.99 | 2 | 1325 |
| DlWRKY41 | Dlo_026484.1 | scaffold607:21585:23785 | 1218 | 406 | 6.06 | 45.38 | 4 | 2201 |
| DlWRKY42 | Dlo_027244.2 | scaffold640:85638:89083 | 2298 | 766 | 5.15 | 83.59 | 4 | 3446 |
| DlWRKY43 | Dlo_027361.1 | scaffold648:191661:193182 | 969 | 323 | 9.14 | 36.57 | 2 | 1522 |
| DlWRKY44 | Dlo_027614.1 | scaffold657:107511:111179 | 1521 | 507 | 5.55 | 54.88 | 4 | 3669 |
| DlWRKY45 | Dlo_029034.1 | scaffold711:179562:181398 | 1539 | 513 | 8.27 | 55.15 | 2 | 1837 |
| DlWRKY46 | Dlo_029939.1 | scaffold757:33093:37889 | 1710 | 570 | 6.38 | 61.42 | 5 | 4797 |
| DlWRKY47 | Dlo_031466.1 | scaffold829:42224:45497 | 1023 | 341 | 7.20 | 7.71 | 4 | 3274 |
| DlWRKY48 | Dlo_031469.1 | scaffold829:58277:59797 | 990 | 330 | 9.06 | 36.21 | 3 | 1521 |
| DlWRKY49 | Dlo_031936.1 | scaffold858:266912:269300 | 588 | 196 | 9.46 | 22.05 | 1 | 2389 |
| DlWRKY50 | Dlo_032595.1 | scaffold896:87649:89238 | 1185 | 395 | 6.67 | 43.04 | 2 | 1590 |
| DlWRKY51 | Dlo_033966.1 | scaffold980:88739:90132 | 933 | 311 | 5.14 | 34.86 | 2 | 1394 |
| DlWRKY52 | Dlo_001658.1 | scaffold1077:66972:68290 | 918 | 306 | 6.26 | 33.96 | 4 | 1319 |
| DlWRKY53 | Dlo_002663.1 | scaffold1135:95286:97669 | 1929 | 643 | 5.73 | 70.07 | 4 | 2384 |
| DlWRKY54 | Dlo_004749.1 | scaffold1314:73982:75144 | 795 | 265 | 5.24 | 30.27 | 2 | 1163 |
| DlWRKY55 | Dlo_010873.1 | scaffold2042:2013:3910 | 1023 | 341 | 9.42 | 37.97 | 3 | 1898 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Liu, C.; Xie, J.; Shi, S. Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 2169. https://doi.org/10.3390/ijms19082169
Jue D, Sang X, Liu L, Shu B, Wang Y, Liu C, Xie J, Shi S. Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses. International Journal of Molecular Sciences. 2018; 19(8):2169. https://doi.org/10.3390/ijms19082169
Chicago/Turabian StyleJue, Dengwei, Xuelian Sang, Liqin Liu, Bo Shu, Yicheng Wang, Chengming Liu, Jianghui Xie, and Shengyou Shi. 2018. "Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses" International Journal of Molecular Sciences 19, no. 8: 2169. https://doi.org/10.3390/ijms19082169
APA StyleJue, D., Sang, X., Liu, L., Shu, B., Wang, Y., Liu, C., Xie, J., & Shi, S. (2018). Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses. International Journal of Molecular Sciences, 19(8), 2169. https://doi.org/10.3390/ijms19082169