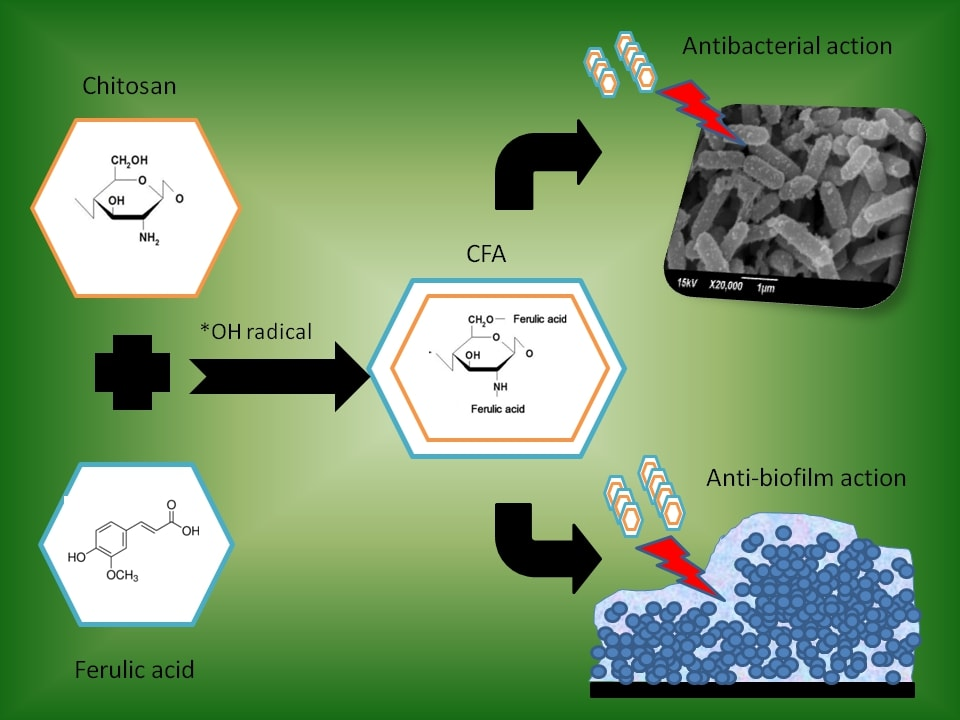
Antibacterial and Biofilm Modulating Potential of Ferulic Acid-Grafted Chitosan against Human Pathogenic Bacteria



Abstract
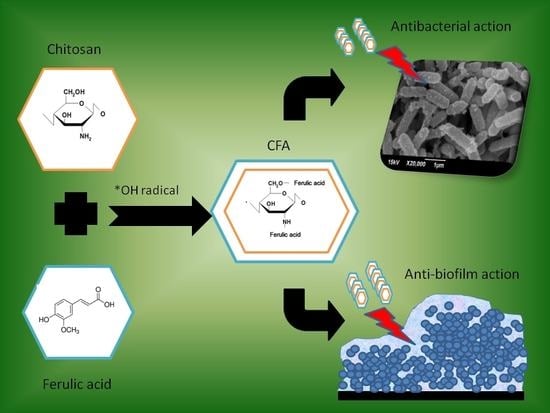
1. Introduction
2. Results
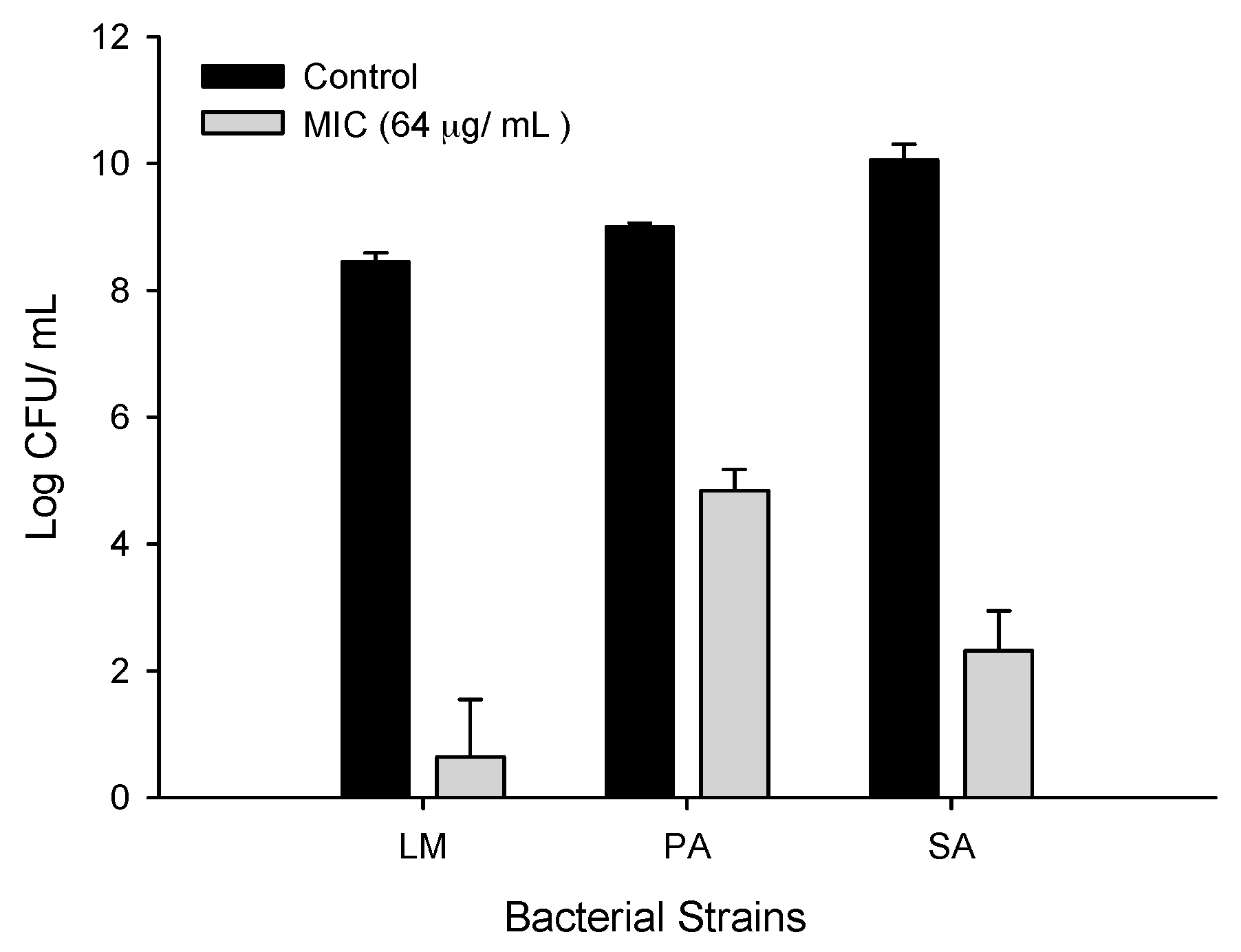
2.1. Determination of Minimum Inhibitory Concentration (MIC) of Ferulic Acid-Grafted Chitosan (CFA)
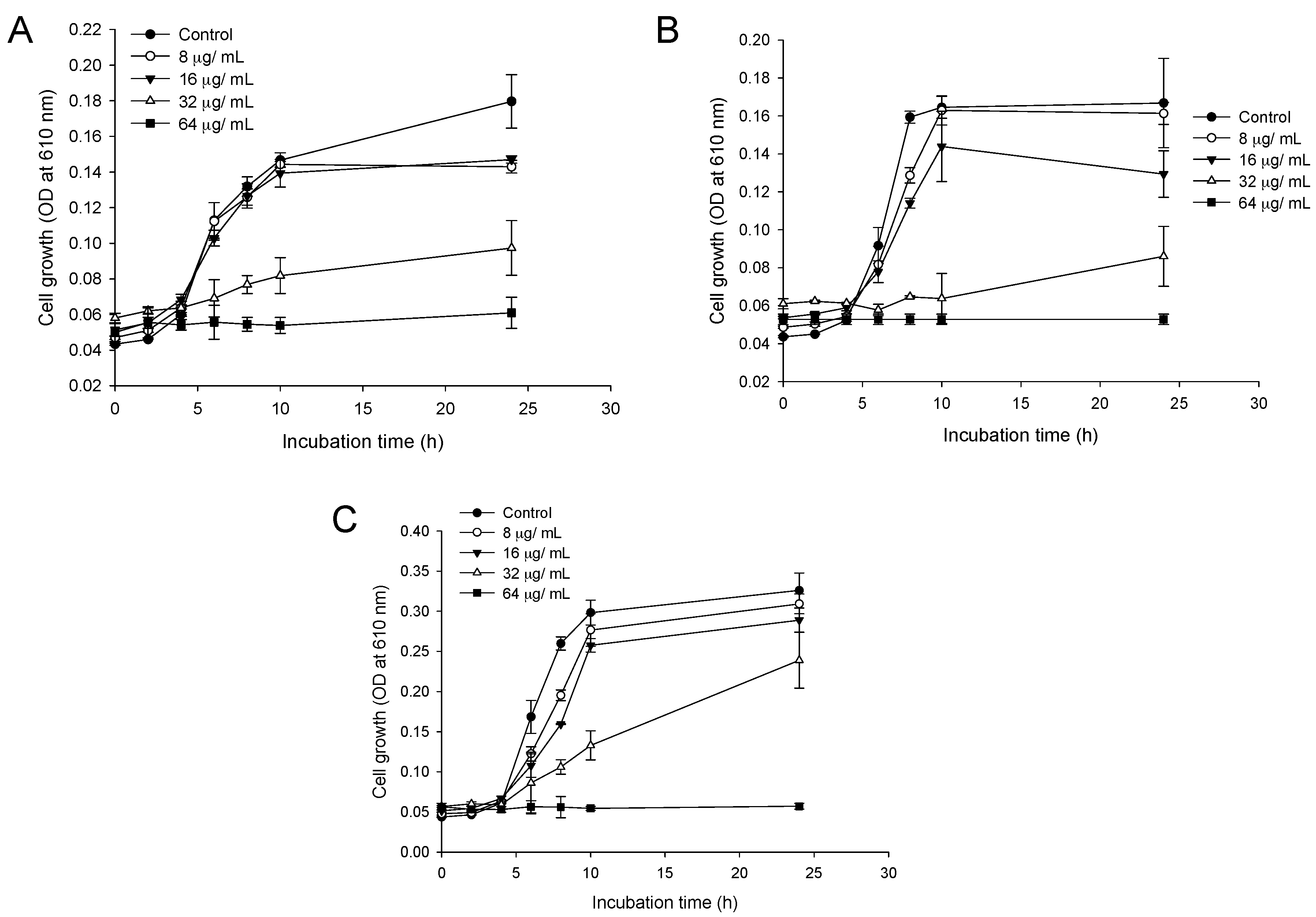
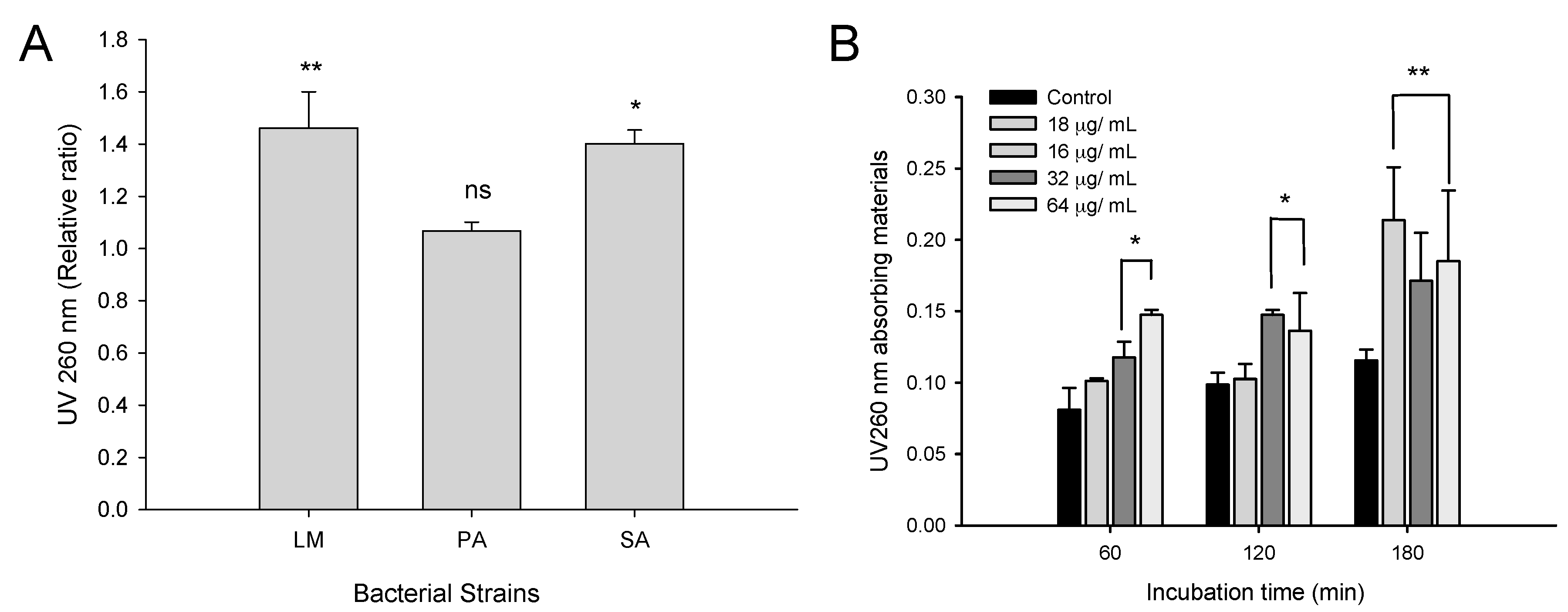
2.2. Time-Dependent Growth Inhibition Studies
2.3. Antibacterial Activity of CFA
2.3.1. Antibacterial Activity of CFA against Gram-Positive Bacteria
2.3.2. Effect of CFA on Bacterial Membrane Integrity
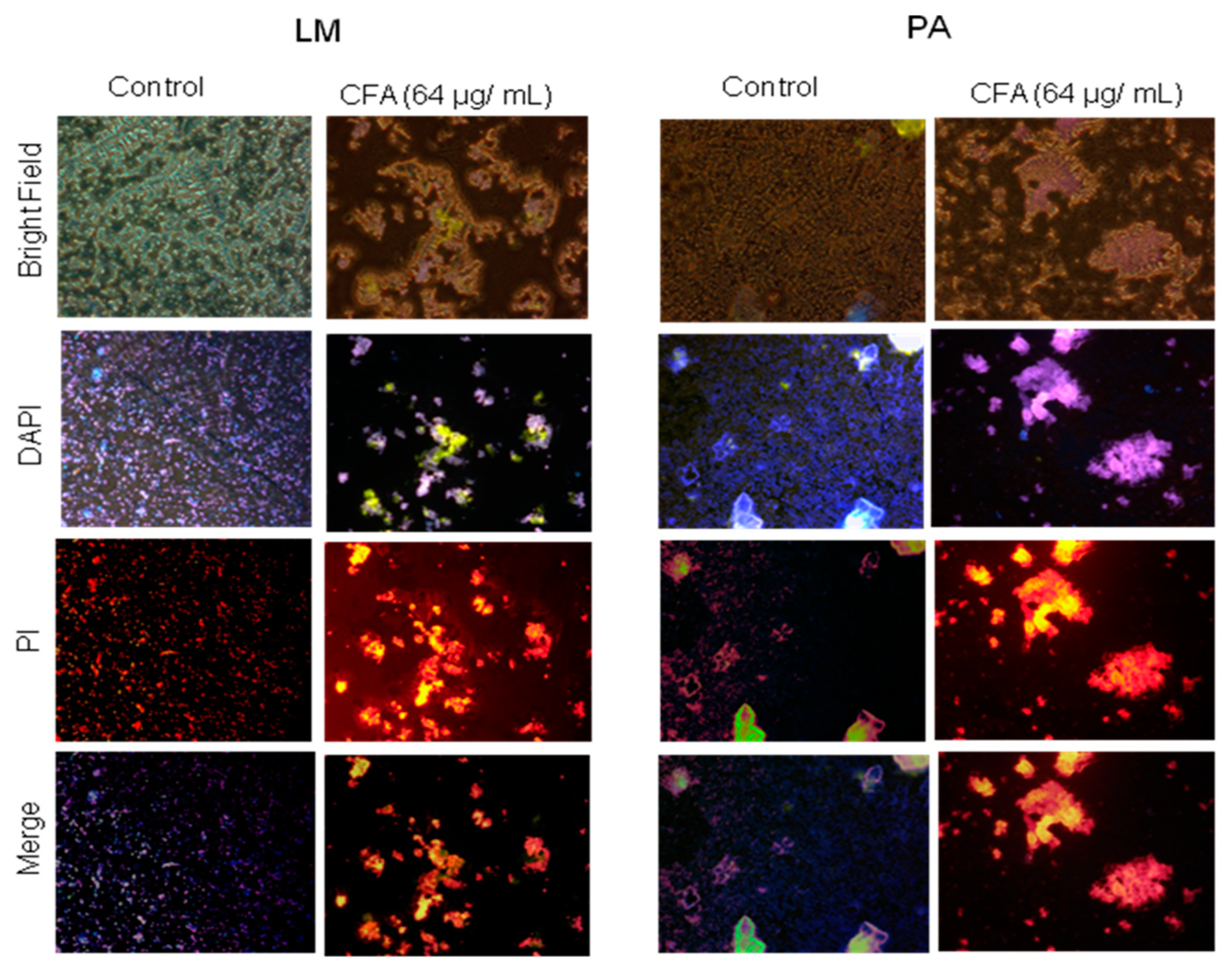
2.3.3. Effect of CFA on Membrane Permeability in Gram-Positive Bacteria
2.3.4. Effect of CFA on the Cellular Morphology
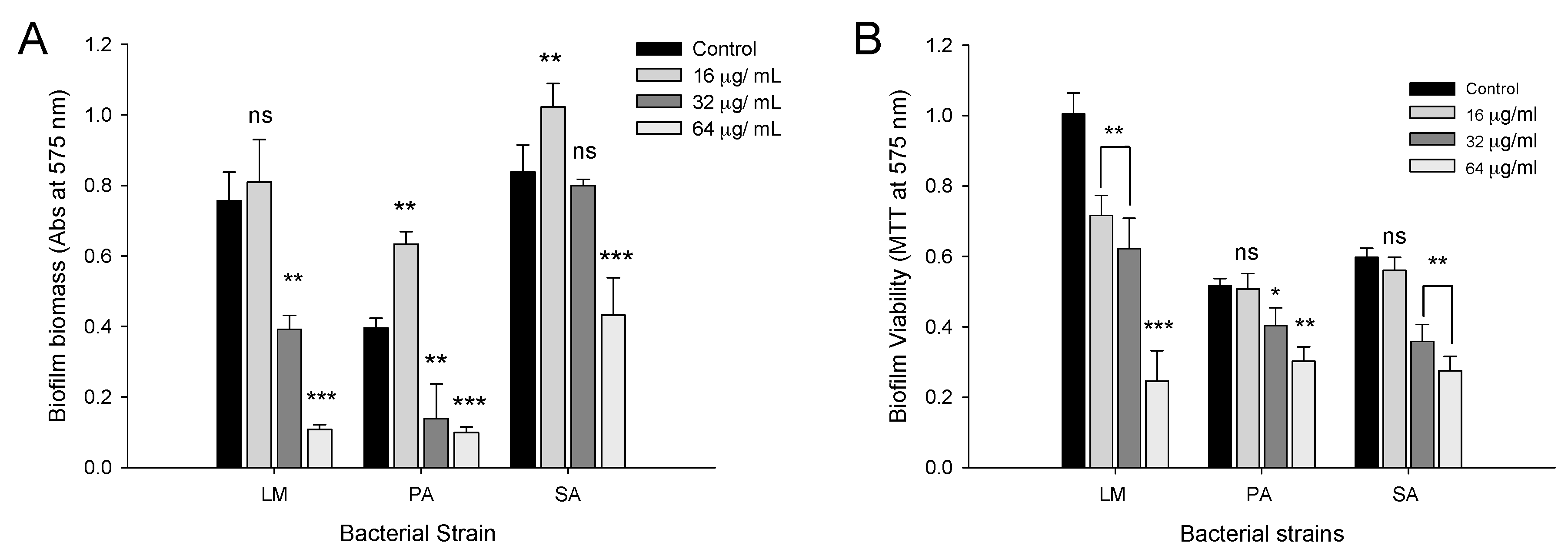
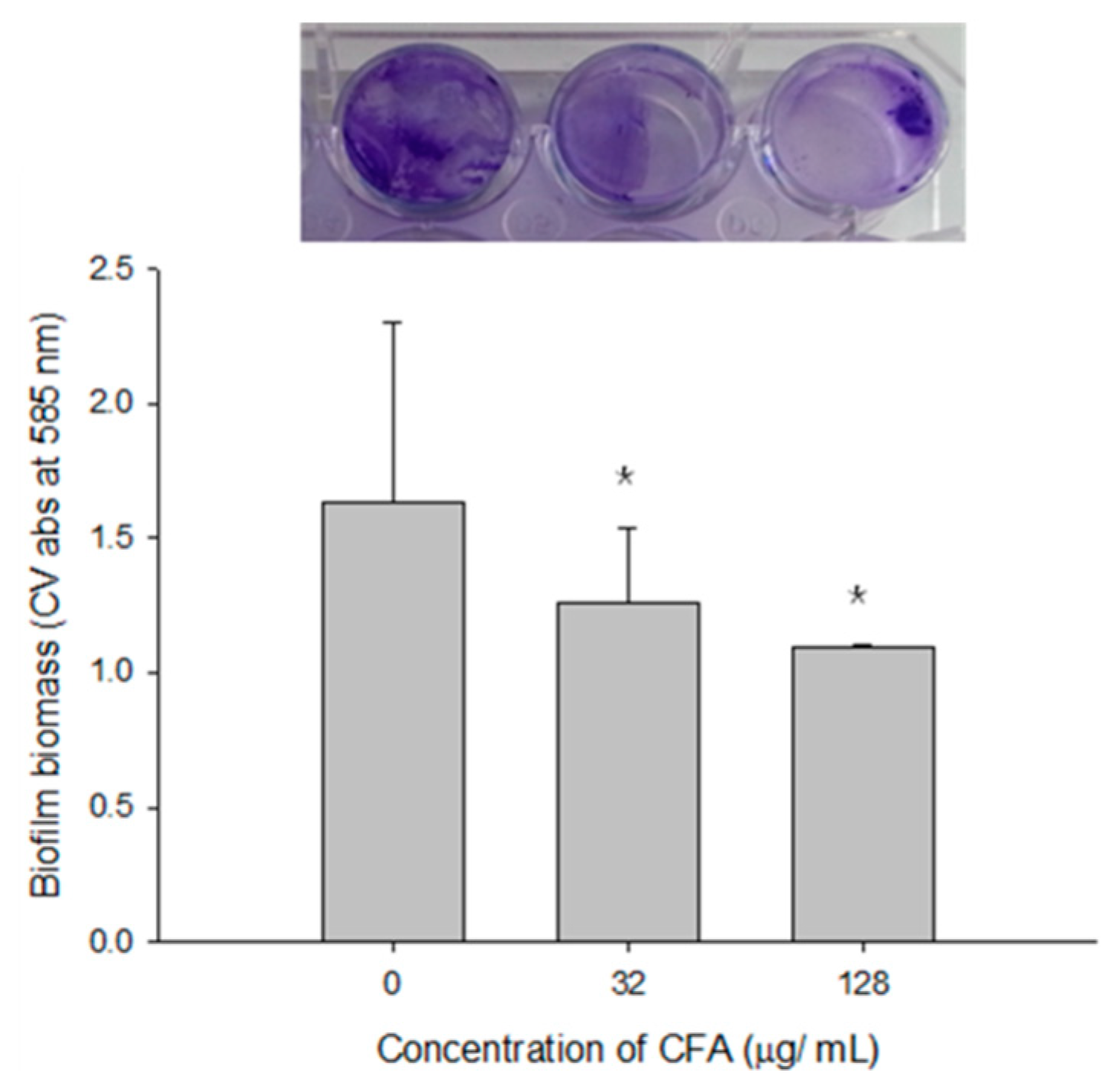
2.4. Biofilm Inhibition Properties of CFA
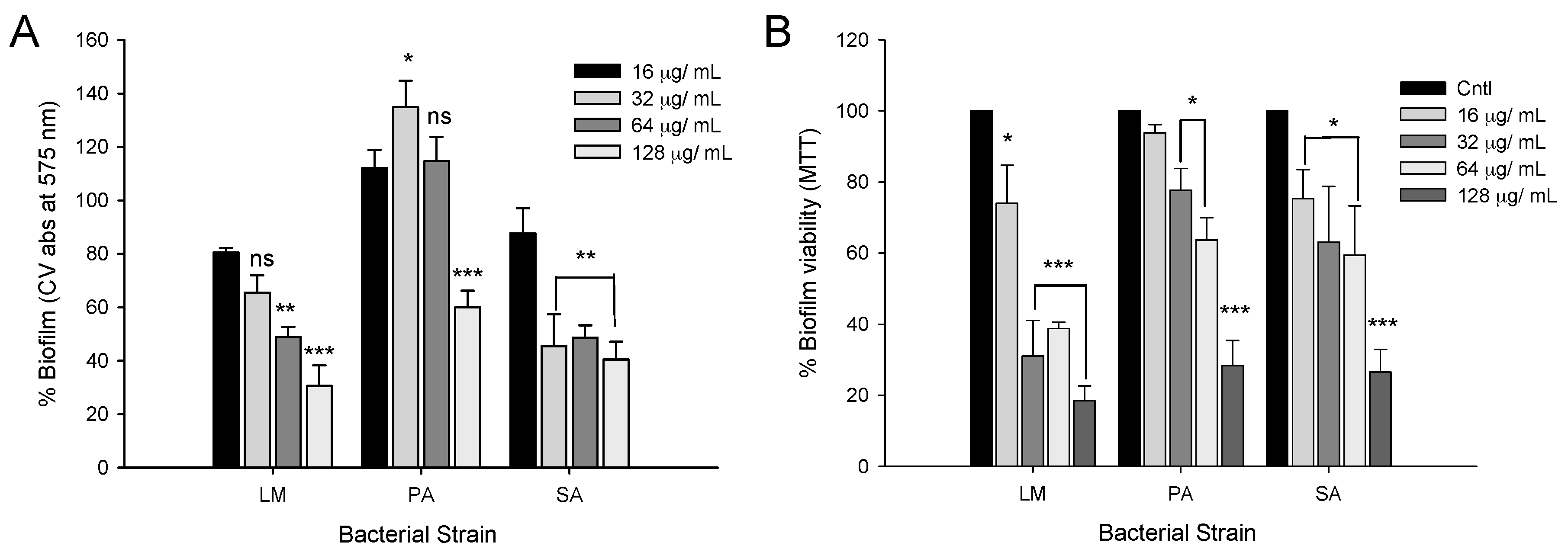
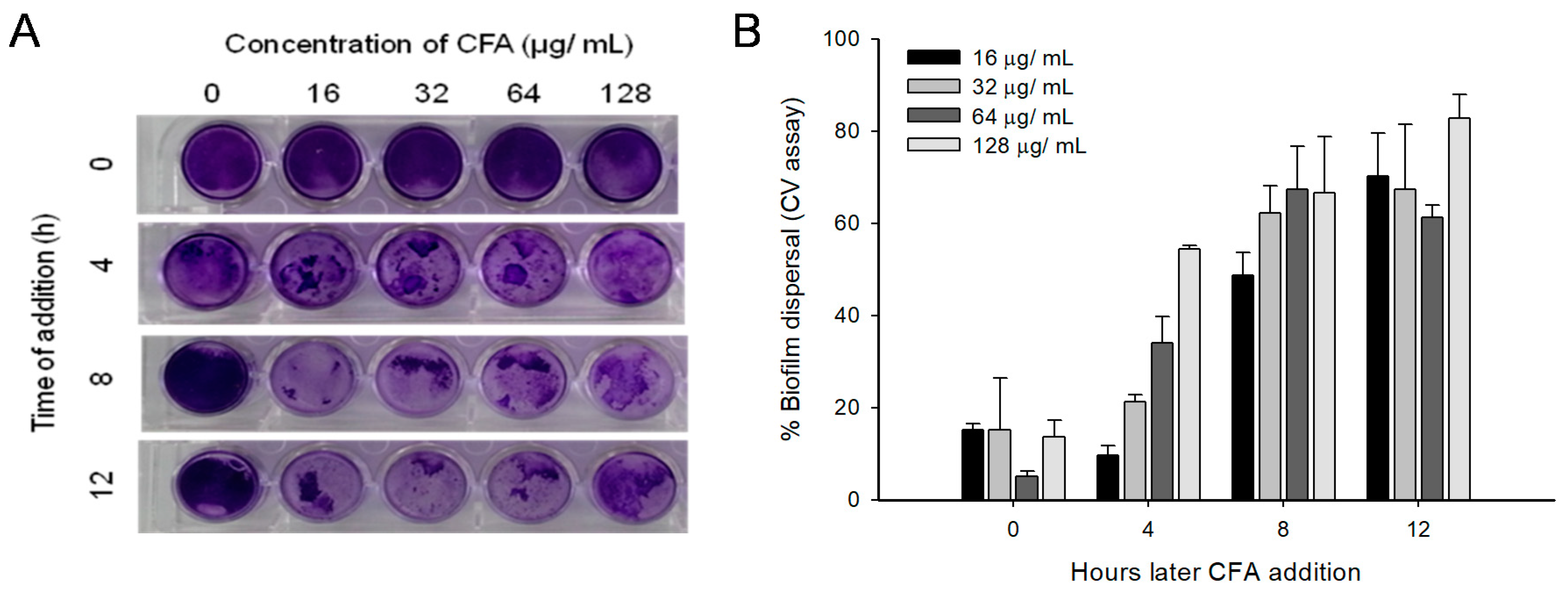
2.5. Disruption of Preformed Mature Biofilms by CFA
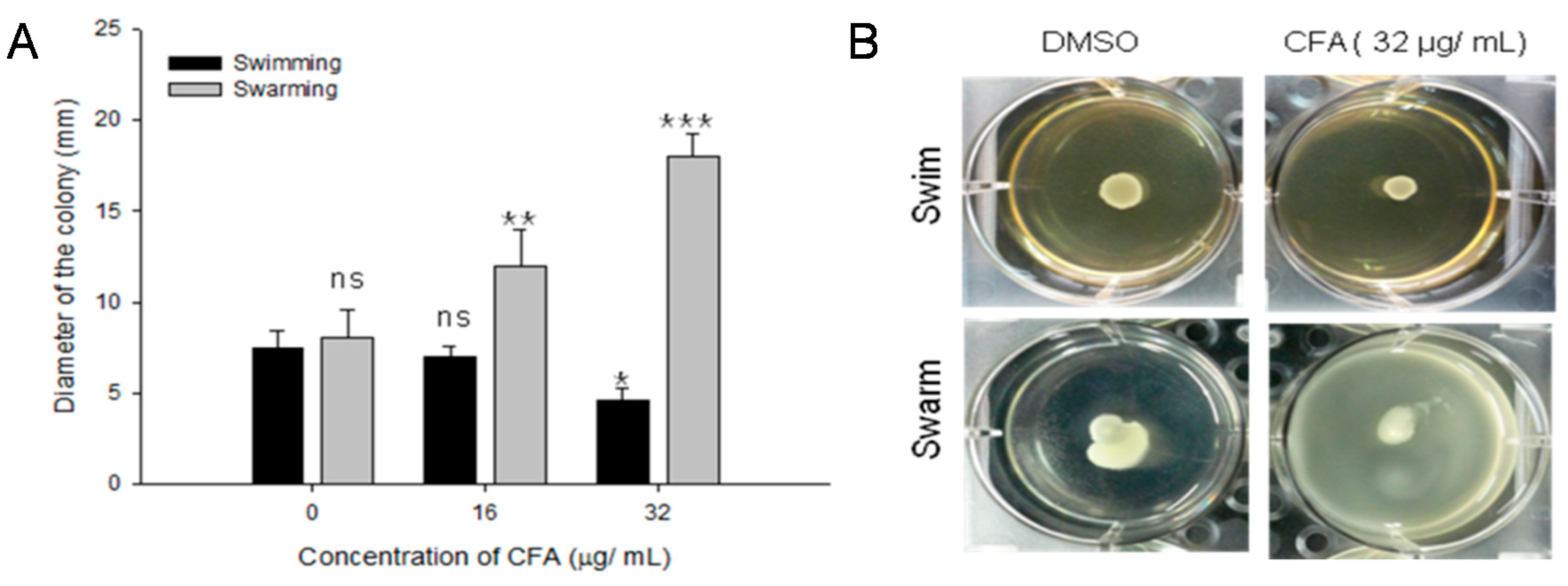
2.6. Effect of CFA on Bacterial Motility
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Bacterial Strains and Culture Conditions
4.3. Antibacterial Assay
4.4. Growth Curves and Viability Assays
4.5. Quantification of Cytoplasmic Materials Released from the Cell
4.6. Propidium Iodide (PI) and 4′,6-Diamidino-2-Phenylindole (DAPI) Uptake Assay
4.7. Scanning Electron Microscopy (SEM)
4.8. Biofilm Modulating Potential of CFA
4.8.1. Inhibition of Initial Attachment
4.8.2. Effect on Preformed Biofilms
4.8.3. Biofilm Estimation by Crystal Violet Staining Method
4.8.4. Biofilm Viability Determination by MTT Assay
4.8.5. Effect on Bacterial Motility
4.8.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LM | Listeria monocytogens |
| PA | Pseudomonas aeruginosa |
| SA | Staphylococcus aureus |
| CS | Chitosan |
| UMC | Unmodified Chitosan |
| CFA | Chitosan Conjugated Ferulic Acid |
| MBC | Minimum Bactericidal Concentration |
| MIC | Minimum Inhibitory Concentration |
| PI | Propidium Iodide |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| SEM | Scanning Electron Microscopy |
| DMSO | Dimethyl Sulfoxide |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide |
| PBS | Phosphate Buffered Saline |
| TSB | Tryptic Soy Broth |
| MHB | Muller-Hinton Broth |
| CLSM | Confocal Laser Scanning Microscopy |
References
- Begley, M.; Hill, C.; Gahan, C.G.M. Identification and disruption of btlA, a locus involved in bile tolerance and general stress resistance in Listeria monocytogenes. FEMS Microbiol. Lett. 2003, 218, 31–38. [Google Scholar] [CrossRef] [PubMed]
- De Noordhout, C.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Long, T.E. Repurposing thiram and disulfiram as antibacterial agents for multidrug-resistant Staphylococcus aureus infections. Antimicrob. Agents Chemother. 2017, 61, e00898. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H.; Chan, W.Y. Chitosan: An Update on Potential Biomedical and Pharmaceutical Applications. Mar. Drugs 2015, 13, 5156–5186. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Kim, S.K.; Je, J.Y. Chitosan gallate as potential antioxidant biomaterial. Bioorg. Med. Chem. Lett. 2011, 21, 3070–3073. [Google Scholar] [CrossRef] [PubMed]
- Senevirathne, M.; Jeon, Y.J.; Kim, Y.T.; Park, P.J.; Jung, W.K.; Ahn, C.B.; Je, J.Y. Prevention of oxidative stress in Chang liver cells by gallic acid-graf ted-chitosans. Carbohydr. Polym. 2012, 87, 876–880. [Google Scholar] [CrossRef]
- Cho, Y.S.; Kim, S.K.; Ahn, C.B.; Je, J.Y. Inhibition of acetylcholinesterase by gallic acid-grafted-chitosans. Carbohydr. Polym. 2011, 84, 690–693. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, D.; Eom, S.H.; Kim, S.H.; Oh, J.; Jung, W.K.; Kim, Y.M. Synergistic antibacterial effects of chitosan-caffeic acid conjugate against antibiotic-resistant acne-related bacteria. Mar. Drugs 2017, 15, 167. [Google Scholar] [CrossRef] [PubMed]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Saavedra, M.J.; Simoes, M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling 2012, 28, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Je, J.Y. Gallic acid grafted chitosan inhibits food borne pathogens by a membrane damage mechanism. J. Agric. Food Chem. 2013, 61, 6574–6579. [Google Scholar] [CrossRef] [PubMed]
- Champer, J.; Patel, J.; Fernando, N.; Salehi, E.; Wong, V.; Kim, J. Chitosan against cutaneous pathogens. AMB Express 2013, 3, 37. [Google Scholar] [CrossRef] [PubMed]
- Raafat, D.; Bargen, K.V.; Haas, A.; Sahl, H.G. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol. 2008, 74, 3764–3773. [Google Scholar] [CrossRef] [PubMed]
- Dola, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2003, 8, 881–890. [Google Scholar] [CrossRef]
- Machado, I.; Grac, J.; Sousa, A.M.; Lopes, S.P.; Pereira, M.O. Effect of antimicrobial residues on early adhesion and biofilm formation by wild-type and benzalkonium chloride-adapted Pseudomonas aeruginosa. Biofouling 2011, 27, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Asli, A.; Brouillette, E.; Ster, C.; Ghinet, M.G.; Brzezinski, R.; Lacasse, P.; Jacques, M.; Malouin, F. Antibiofilm and antibacterial effects of specific chitosan molecules on Staphylococcus aureus isolates associated with bovine mastitis. PLoS ONE 2017, 12, e0176988. [Google Scholar] [CrossRef] [PubMed]
- Thaya, R.; Vasheeharan, B.; Sivakamavalli, J.; Iswarya, A.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnam, S.; Alanbr, M.N.; Khaled, J.M.; Benneli, G. Synthesis of chitosan-alginate microspheres with high antimicrobial and antibiofilm activity against multidrug-resistant microbial pathogens. Microb. Pathog. 2018, 114, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Amato, A.; Migneco, L.M.; Martinelli, A.; Pietrelli, L.; Piozzi, A.; Francolini, I. Antimicrobial activity of catechol-functionalized-chitosan versus Staphylococcus epidermidis. Carbohydr. Polym. 2018, 179, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Lee, H.Y.; Ahn, J. Characteristics of biofilm formation by selected foodborne pathogens. J. Food Saf. 2011, 31, 91–97. [Google Scholar] [CrossRef]
- Orgaz, M.M.; Lobete, C.H.; Puga, C.; Jose, S. Effectiveness of chitosan against mature biofilms formed by food related bacteria. Int. J. Mol. Sci. 2011, 12, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Caiazza, N.C.; Merrriitt, J.H.; Brothers, K.M.; O’Toole, G.A. Inverse regulation of biofilm formation and swarming motility by Pseudomonas aeruginosa PA14. J. Bacteriol. 2007, 189, 3602–3612. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, J.; Kan, J.; Jin, C. Synthesis of chitosan-gallic acid conjugate: Structure characterization and in vitro anti-diabetic potential. Int. J. Biol. Macromol. 2013, 62, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, H.; Lu, J.; Wen, X.; Kan, J.; Jin, C. Preparation and characterization of novel phenolic acid (hydroxybenzoic and hydroxycinnamic acid derivatives) grafted chitosan microspheres with enhanced adsorption properties for Fe(II). Chem. Eng. J. 2015, 262, 803–812. [Google Scholar] [CrossRef]
- Hu, B.; Ma, F.; Yang, Y.; Xie, M.; Zhang, C.; Xu, Y.; Zeng, X. Antioxidant nanocomplexes for delivery of epigallocatechin-3-gallate. J. Agric. Food Chem. 2016, 64, 3422–3429. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.B.; Bozell, J.J.; Hayes, D.G.; Zivanovic, S. Introduction of primary antioxidant activity to chitosan for application as a multifunctional food packaging material. Food Hydrocoll. 2013, 33, 207–214. [Google Scholar] [CrossRef]
- Lee, S.J.; Kang, M.S.; Oh, J.S.; Na, H.S.; Lim, Y.J.; Jeong, Y.I.; Lee, H.C. Caffeic acid conjugated chitosan derivatives and their antitumor activity. Arch. Pharm. Res. 2013, 36, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory and Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 10th ed.; Approved Standards, CLSI document M07-A10; Clinical and Laboratory and Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Reddy, M.V.; Thota, N.; Sangwan, P.L.; Malhotra, P.; Ali, F.; Khan, I.A.; Chimni, S.S.; Koul, S. Novel bisstyryl derivatives of bukuchiol: Targeting oral cavity pathogens. Eur. J. Med. Chem. 2010, 45, 3125–3134. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.M.; Silva, S.; Veiga, M.; Vincente, S.; Tavaria, F.K.; Pintado, M.E. Investigation of chitosan’s antibacterial activity against vancomycin resistant microorganisms and their biofilms. Carbohydr. Polym. 2017, 74, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Sing, M.; Mukhopadhyay, K. C-terminal aminoacids of alpha melanocyte-stimulating hormone are required for its antimicrobial activity against Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 1920–1929. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E.A. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 2013, 29, 125–130. [Google Scholar] [CrossRef]
- Saising, J.; Dube, L.; Ziebandt, A.K.; Voravuthikunchai, S.P.; Nega, M.; Gotz, F. Activity of gallidermin on Staphylococcus aureus and Staphylococcus epidermidis biofilms. Antimicrob. Agents Chemother. 2012, 56, 5804–5810. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Gilbert, E.U. Colorimetric method for identifying plant essential oil components that affect biofilm formation and structure. Appl. Environ. Microb. 2004, 70, 6951–6956. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Tryptic Soy Broth (TSB) | Muller-Hinton Broth (MHB) | ||
|---|---|---|---|---|
| UMC | CFA | UMC | CFA | |
| Listeria monocytogenes KCTC 3569 | 64 | 64 | 64 | 64 |
| Staphylococcus aureus KCTC 1916 | 2048 | 512 | 128 | 64 |
| Pseudomonas aeruginosa KCCM 11321 | 1024 | 512 | 128 | 64 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dasagrandhi, C.; Park, S.; Jung, W.-K.; Kim, Y.-M. Antibacterial and Biofilm Modulating Potential of Ferulic Acid-Grafted Chitosan against Human Pathogenic Bacteria. Int. J. Mol. Sci. 2018, 19, 2157. https://doi.org/10.3390/ijms19082157
Dasagrandhi C, Park S, Jung W-K, Kim Y-M. Antibacterial and Biofilm Modulating Potential of Ferulic Acid-Grafted Chitosan against Human Pathogenic Bacteria. International Journal of Molecular Sciences. 2018; 19(8):2157. https://doi.org/10.3390/ijms19082157
Chicago/Turabian StyleDasagrandhi, Chakradhar, Seulki Park, Won-Kyo Jung, and Young-Mog Kim. 2018. "Antibacterial and Biofilm Modulating Potential of Ferulic Acid-Grafted Chitosan against Human Pathogenic Bacteria" International Journal of Molecular Sciences 19, no. 8: 2157. https://doi.org/10.3390/ijms19082157
APA StyleDasagrandhi, C., Park, S., Jung, W.-K., & Kim, Y.-M. (2018). Antibacterial and Biofilm Modulating Potential of Ferulic Acid-Grafted Chitosan against Human Pathogenic Bacteria. International Journal of Molecular Sciences, 19(8), 2157. https://doi.org/10.3390/ijms19082157