The Expression of CARK1 or RCAR11 Driven by Synthetic Promoters Increases Drought Tolerance in Arabidopsis thaliana

Abstract
:1. Introduction
2. Results
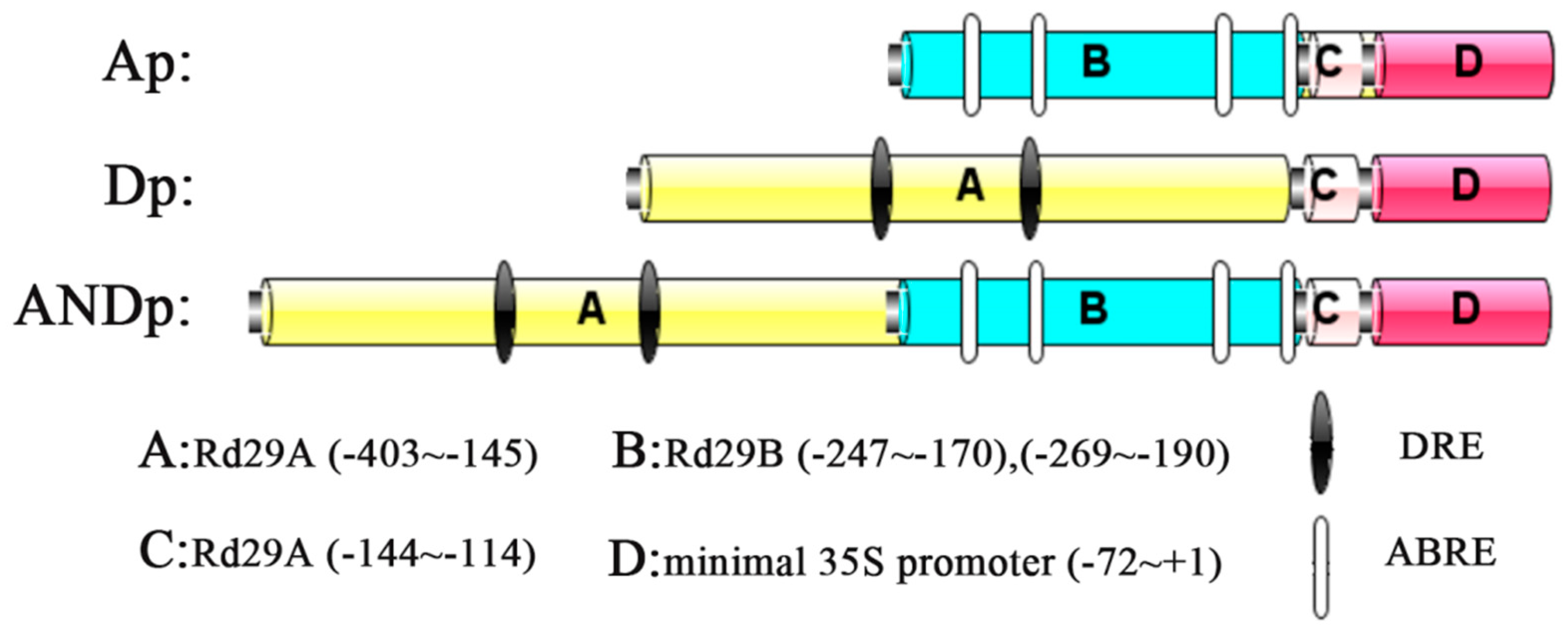
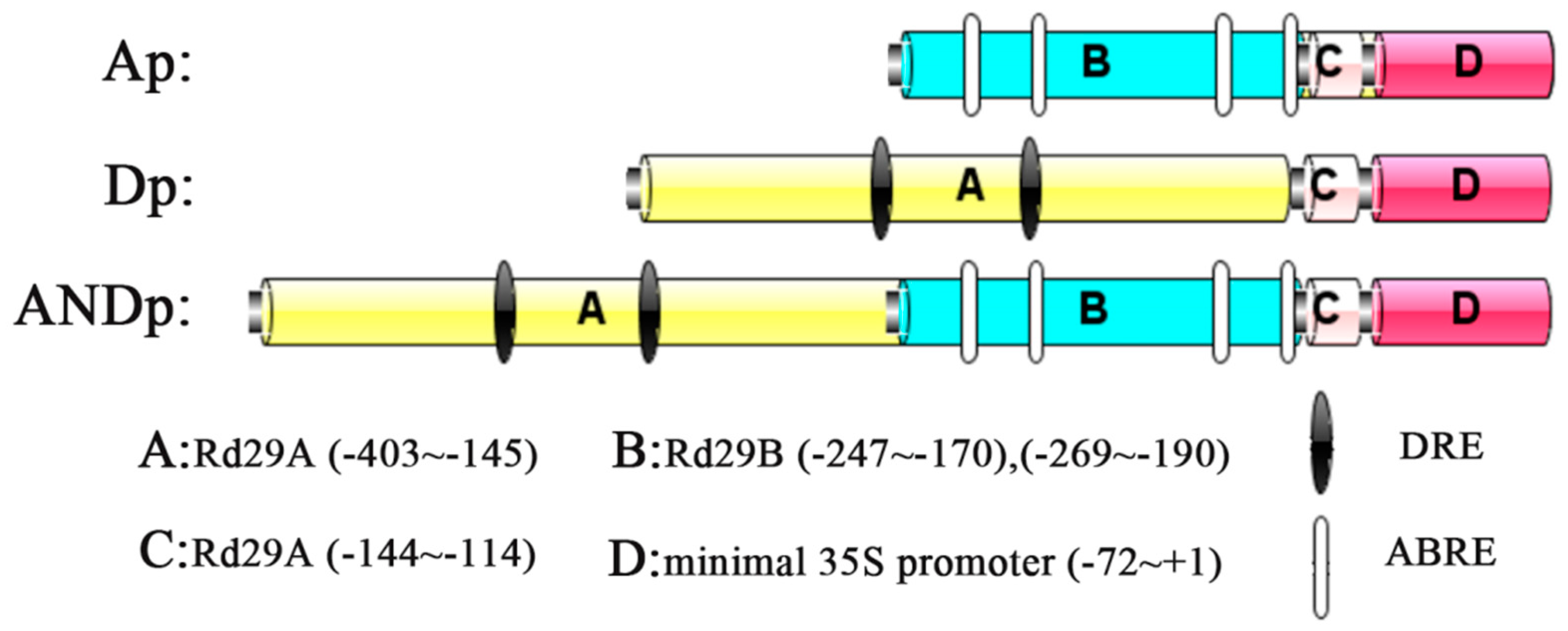
2.1. Selection of Promoter Fragments
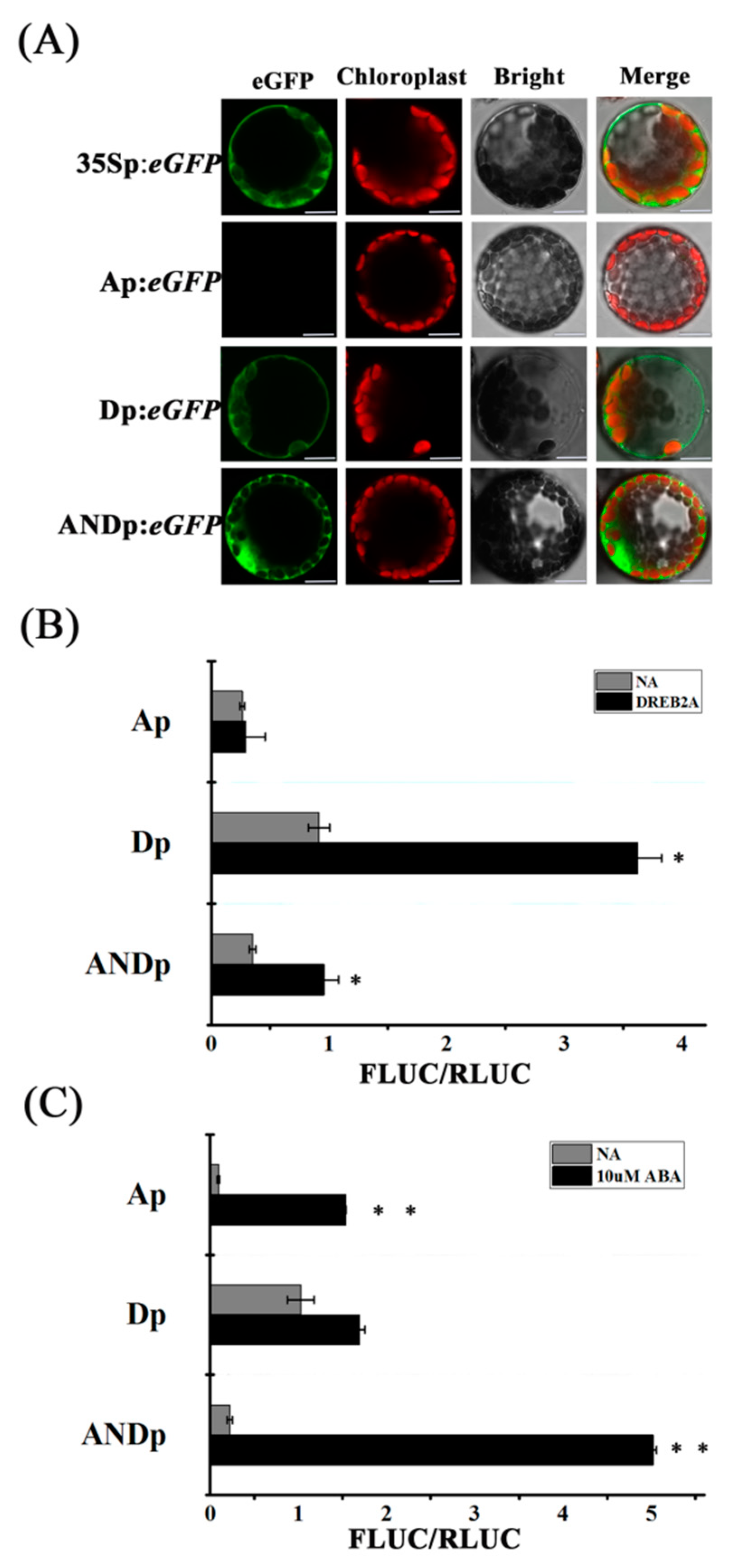
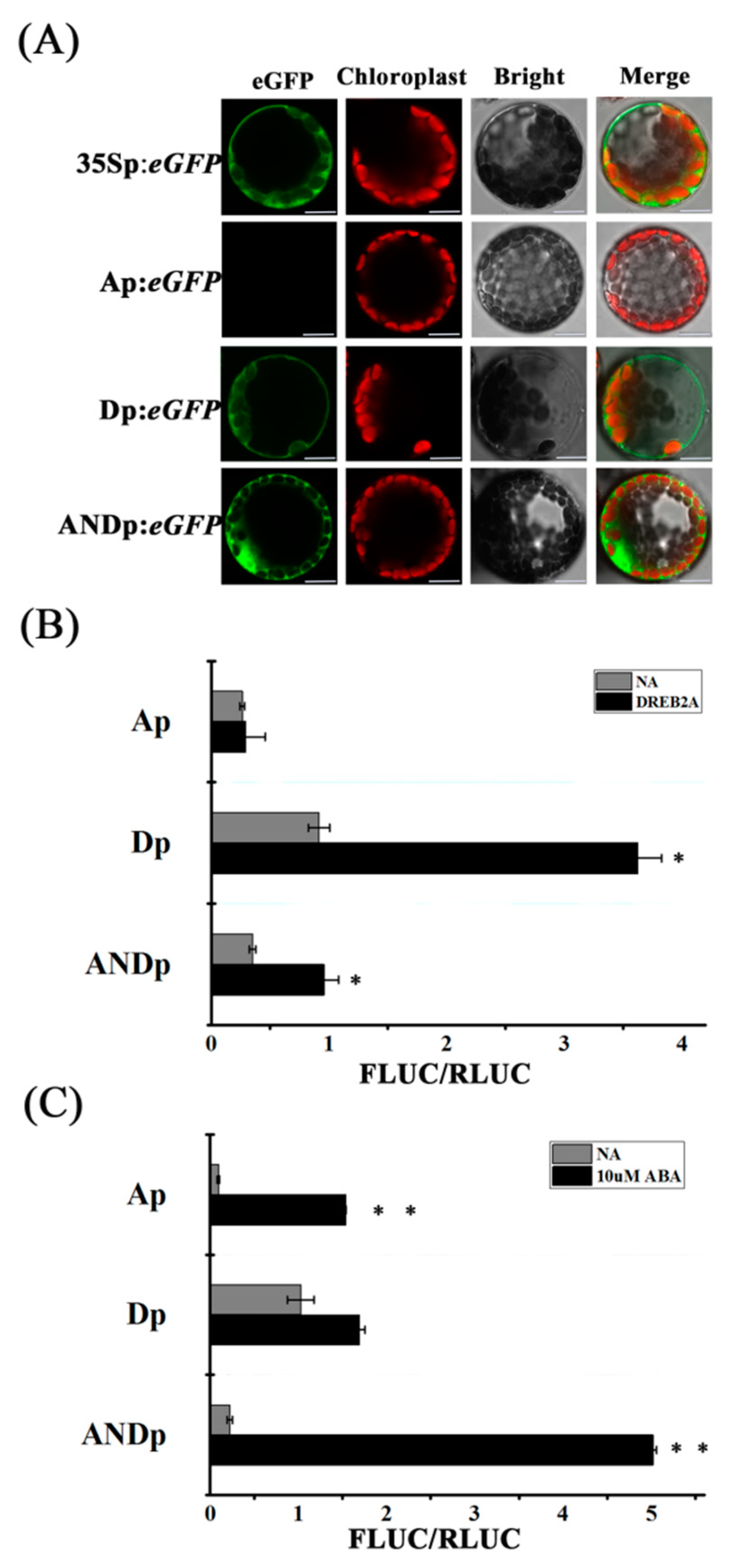
2.2. Confirmation of Transcriptional Activity
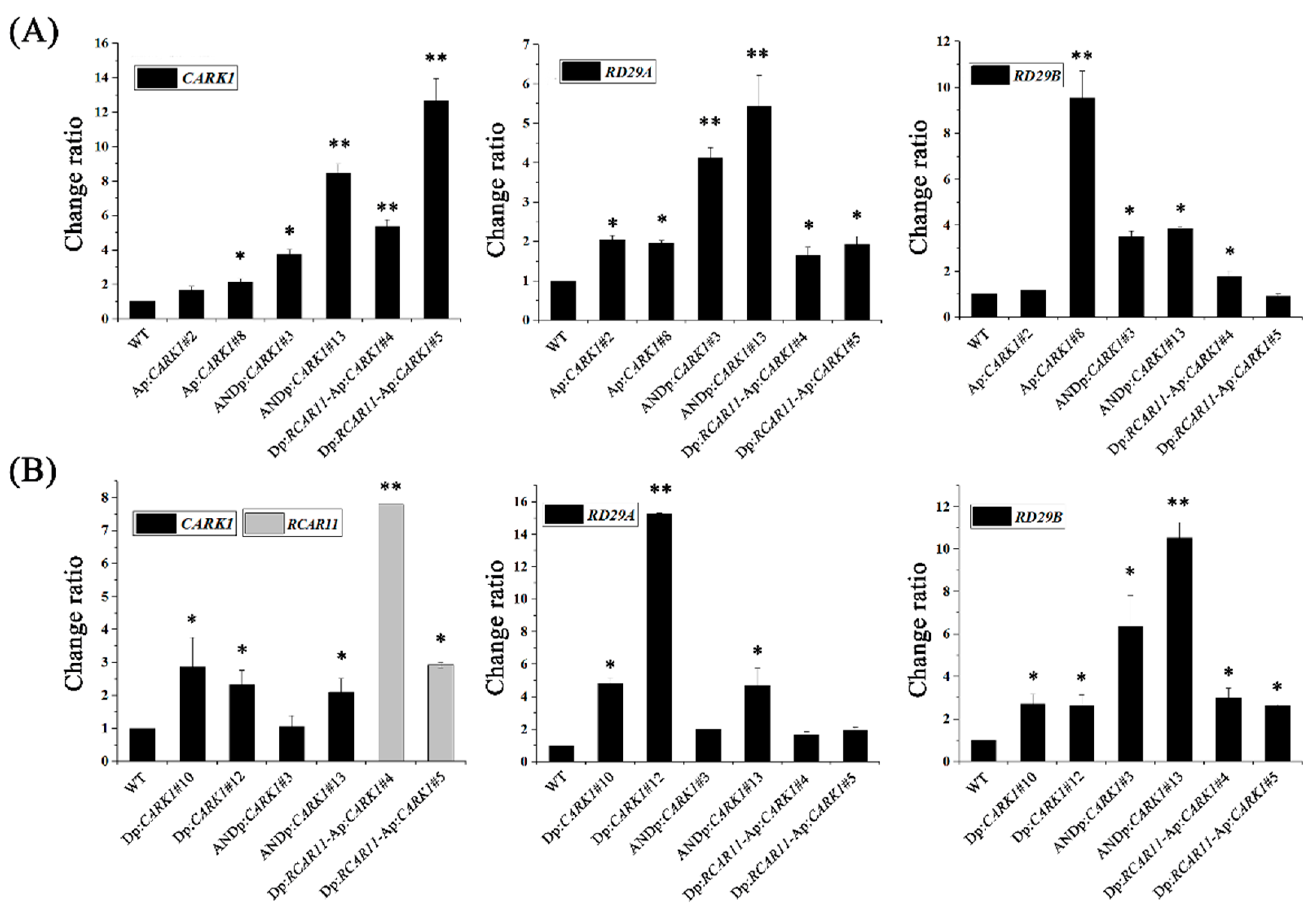
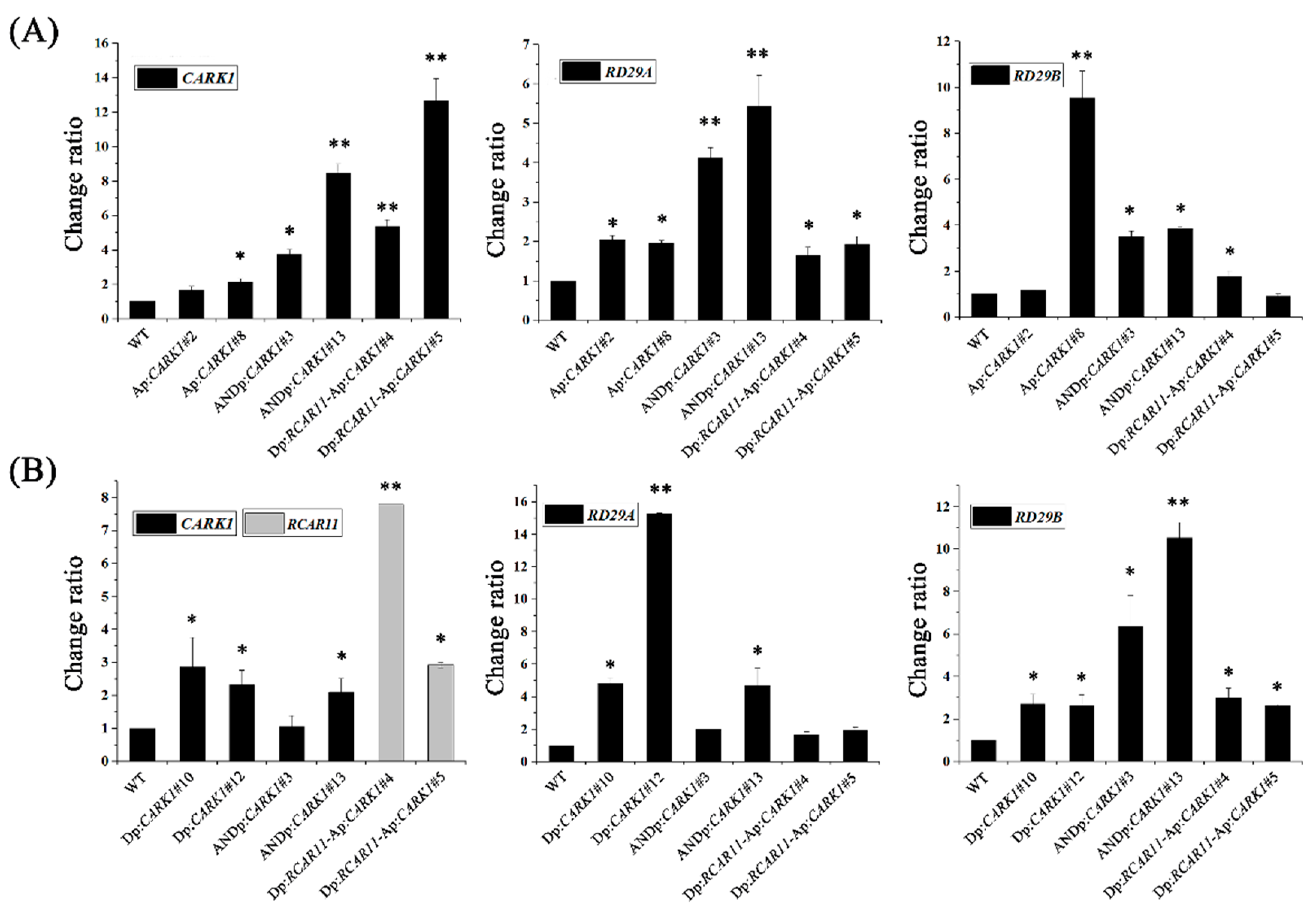
2.3. Synthetic Promoters Increase the Expression of Drought-Related Genes upon Abscisic Acid (ABA) or d-Mannitol Treatment
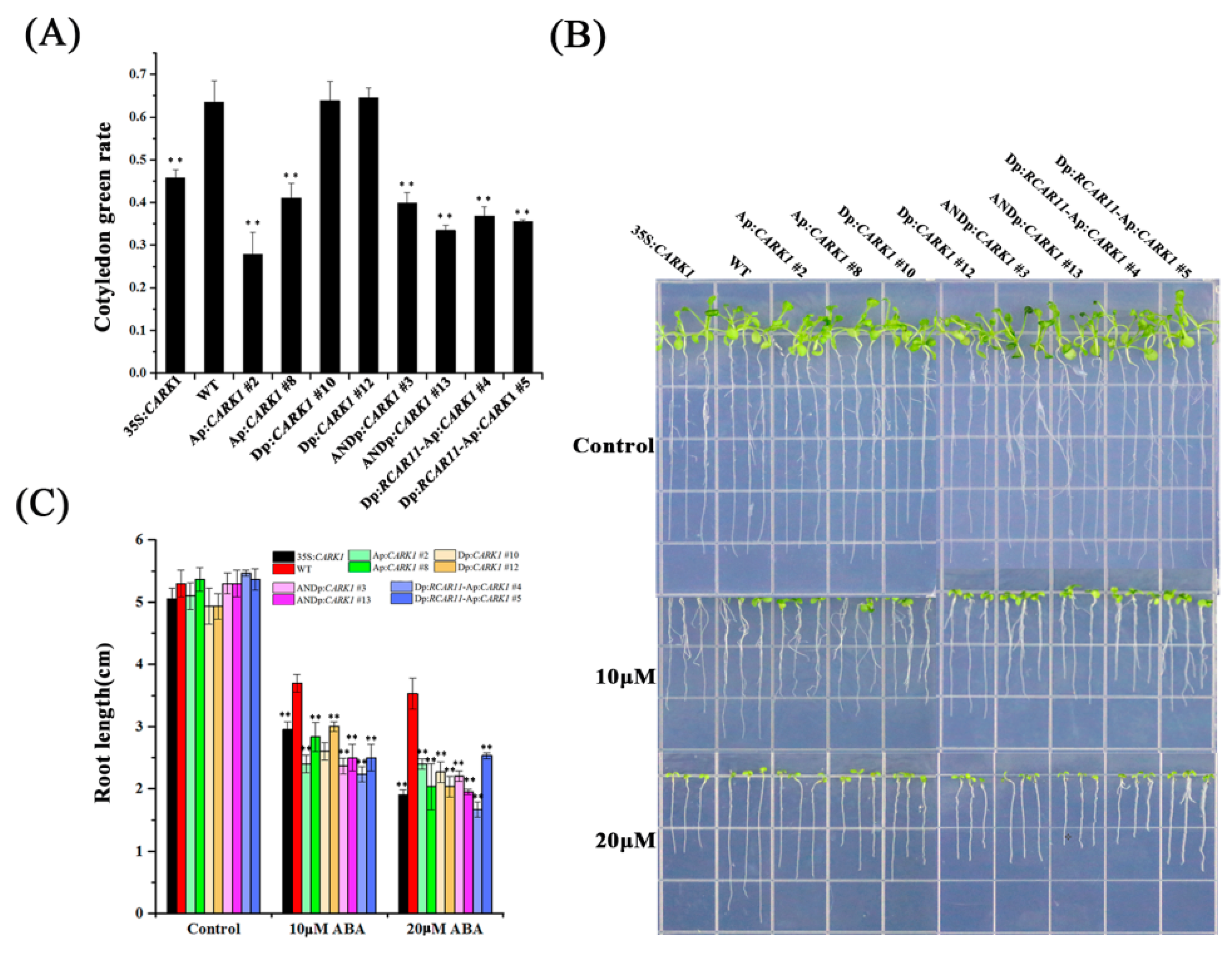
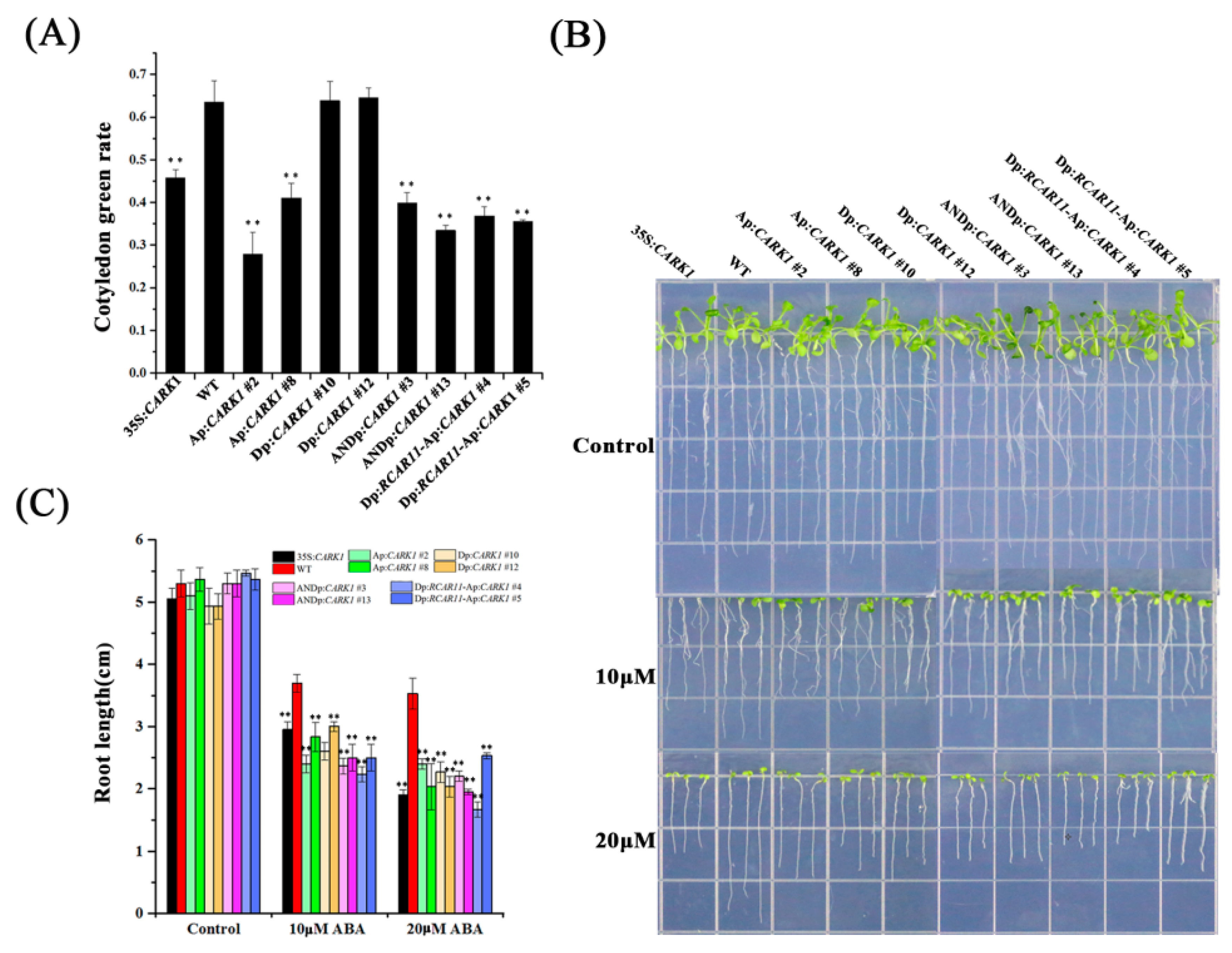
2.4. Synthetic Promoters Regulate Plant Growth in Response to ABA
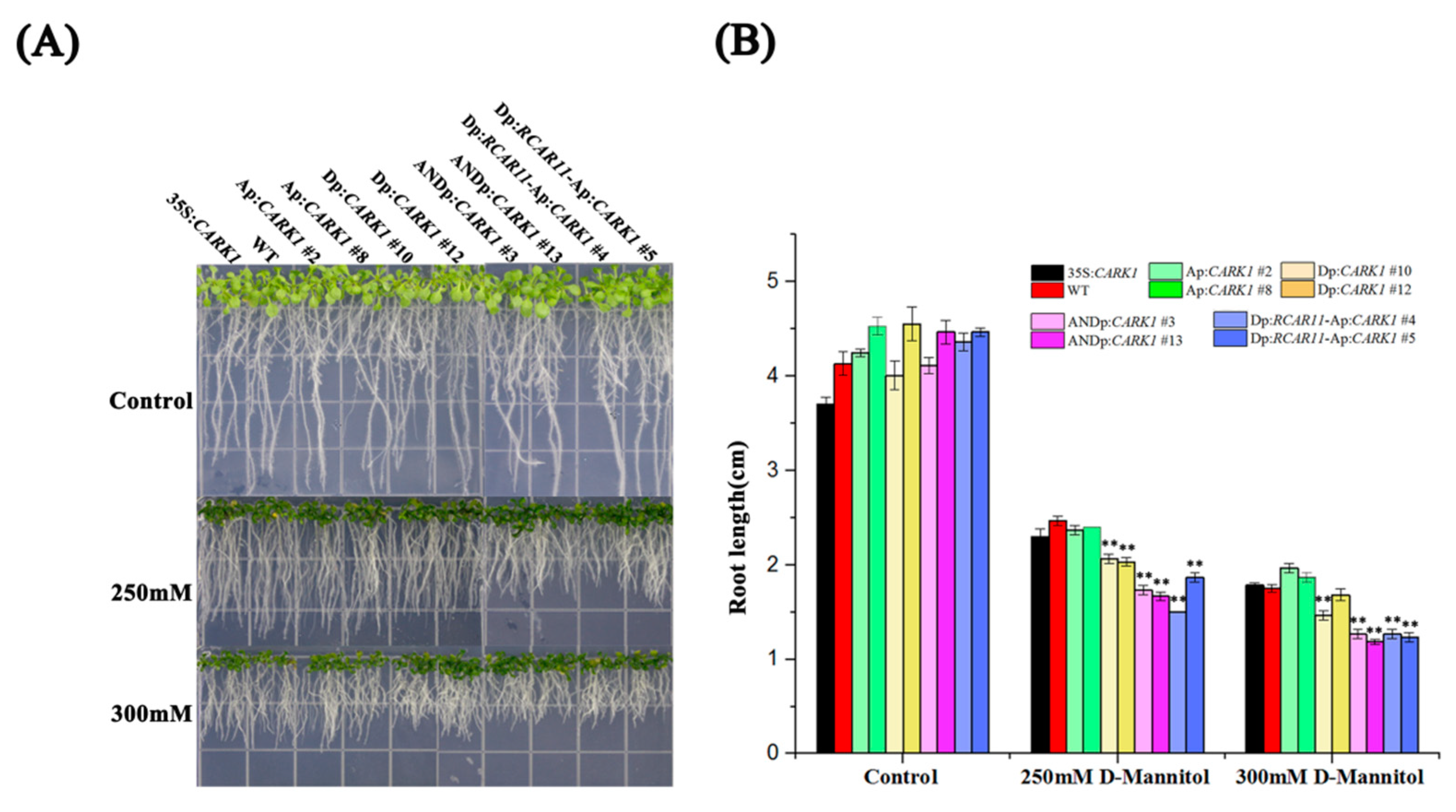
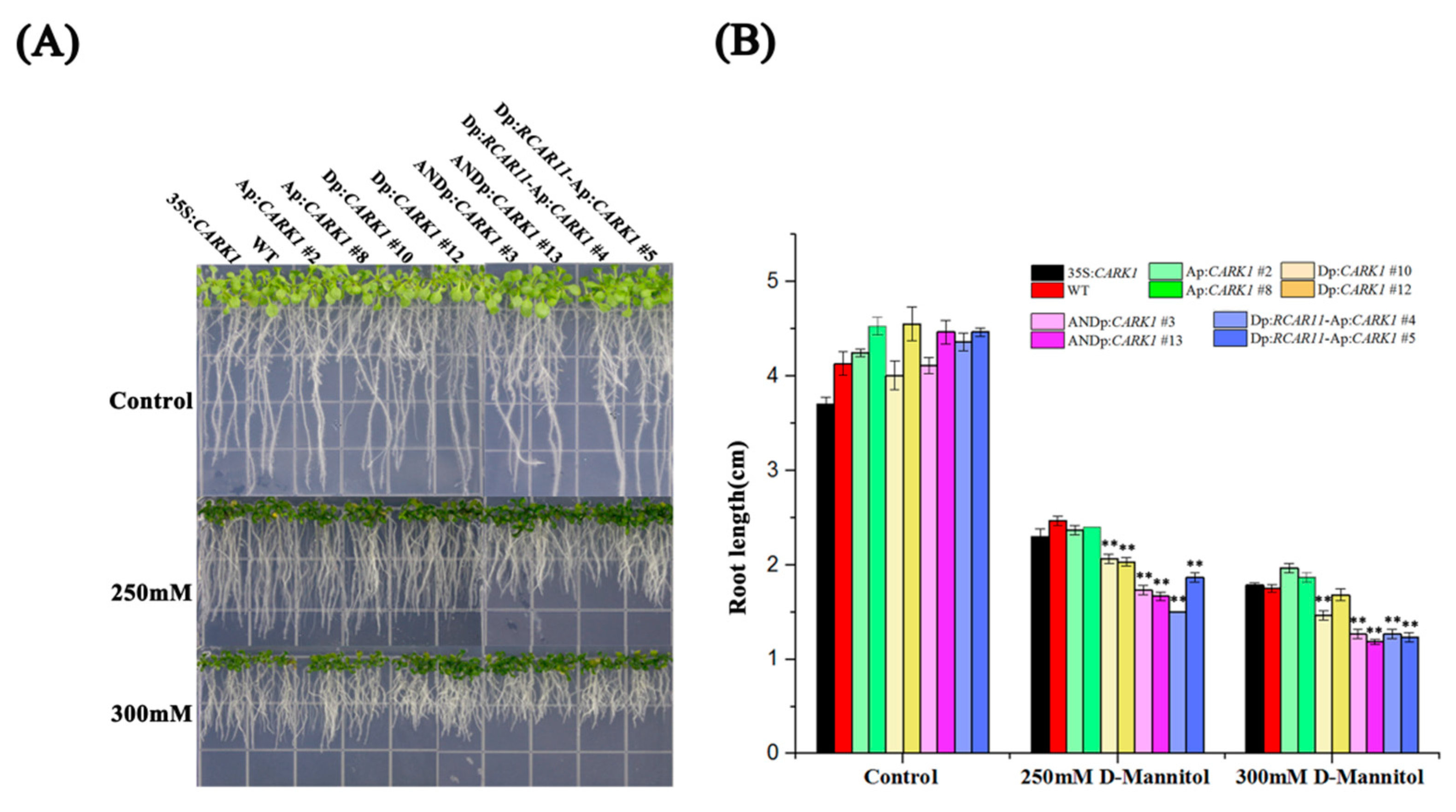
2.5. Synthetic Promoters Enhance Drought Tolerance
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Protoplast Isolation and Transient Activation Assay
4.3. Analysis of Quantitative Real Time-PCR (qRT-PCR)
4.4. Phenotype Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [PubMed]
- Kurek, I.; Chang, T.K.; Bertain, S.M.; Madrigal, A.; Liu, L.; Lassner, M.W.; Zhu, G. Enhanced Thermostability of Arabidopsis Rubisco activase improves photosynthesis and growth rates under moderate heat stress. Plant Cell 2007, 19, 3230–3241. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tao, Y.; Zhu, J.; Miao, J.; Liu, J.; Liu, Y.; Yi, C.; Yang, Z.; Gong, Z.; Liang, G. GNS4, a novel allele of DWARF11, regulates grain number and grain size in a high-yield rice variety. Rice 2017, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, L.; Chang, W.; Li, Z.; Miao, M.; Li, Y.; Yang, J.; Liu, Z.; Tan, J. Overexpression of the maize E3 ubiquitin ligase gene ZmAIRP4 enhances drought stress tolerance in Arabidopsis. Plant Physiol. Biochem. 2017, 123, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhao, Y.; Ran, X.; Guo, L.; Zhao, D.G. Overexpression of a New Chitinase Gene EuCHIT2 EnhancesResistance to Erysiphe cichoracearum dc in Tobacco Plants. Int. J. Mol. Sci. 2017, 18, 2361. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Zhang, F.; Sun, F.; Luo, Q.; Wang, R.; Hu, R.; Chen, M.; Chang, J.; Yang, G.; He, G. A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants. Plant Sci. 2017, 265, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Zhai, N.; Jia, H.; Liu, D.; Liu, S.; Ma, M.; Guo, X.; Li, H. GhMAP3K65, a Cotton Raf-Like MAP3K Gene, Enhances Susceptibility to Pathogen Infection and Heat Stress by Negatively Modulating Growth and Development in Transgenic Nicotiana benthamiana. Int. J. Mol. Sci. 2017, 18, 2462. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, M.; Liu, Q.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Improving Plant Drought, Salt and Freezing Tolerance by Gene Transfer of a Single Stress-Inducible Transcription Factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Su, S.; Ma, L.; Wen, Y.; Wang, X. Molecular cloning and functional characterization of CoFT1, a homolog of FLOWERING LOCUS T (FT) from Camellia oleifera. Gene 2017, 626, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Mundy, J.; Yamaguchi-Shinozaki, K.; Chua, N.H. Nuclear proteins bind conserved elements in the abscisic acid-responsive promoter of a rice rab gene. Proc. Natl. Acad. Sci. USA 1990, 87, 1406–1410. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Izawa, T.; Chua, N.H.; Shimamoto, K. The rab16B Promoter of Rice Contains Two Distinct Abscisic Acid-Responsive Elements. Plant Physiol. 1996, 112, 483–491. [Google Scholar] [CrossRef] [PubMed]
- RoyChoudhury, A.; Roy, C.; Sengupta, D.N. Transgenic tobacco plants overexpressing the heterologous lea gene Rab16A from rice during high salt and water deficit display enhanced tolerance to salinity stress. Plant Cell Rep. 2007, 26, 1839–1859. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.S.; Wilhelm, K.S.; Thomashow, M.F. The 5’-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Mol. Biol. Rep. 1994, 24, 701–713. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Cao, Y.; Liu, X.; Lin, Y.; Ou, Y.; Zhang, H.; Liu, J. Strength comparison between cold-inducible promoters of Arabidopsis cor15a and cor15b genes in potato and tobacco. Plant Physiol. Biochem. 2013, 71, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Behnam, B.; Kikuchi, A.; Celebi-Toprak, F.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Watanabe, K.N. Arabidopsis rd29A:DREB1A enhances freezing tolerance in transgenic potato. Plant Cell Rep. 2007, 26, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Rerksiri, W.; Zhang, X.; Xiong, H.; Chen, X. Expression and promoter analysis of six heat stress-inducible genes in rice. Sci. World J. 2013, 2013, 397401. [Google Scholar] [CrossRef] [PubMed]
- Msanne, J.; Lin, J.; Stone, J.M.; Awada, T. Characterization of abiotic stress-responsive Arabidopsis thaliana RD29A and RD29B genes and evaluation of transgenes. Planta 2011, 234, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Venter, M. Synthetic promoters: Genetic control through cis engineering. Trends Plant Sci. 2007, 12, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Shokouhifar, F.; Zamani, M.R.; Motallebi, M.; Mousavi, A.; Malboobi, M.A. Construction and functional analysis of pathogen-inducible synthetic promoters in Brassica napus. Biol. Plant. 2011, 55, 689–695. [Google Scholar] [CrossRef]
- Liu, W.; Stewart, C.N., Jr. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Scranton, M.A.; Ostrand, J.T.; Georgianna, D.R.; Lofgren, S.M.; Li, D.; Ellis, R.C.; Carruthers, D.N.; Dräger, A.; Masica, D.L.; Mayfield, S.P. Synthetic promoters capable of driving robust nuclear gene expression in the green alga Chlamydomonas reinhardtii. Algal Res. 2016, 15, 135–142. [Google Scholar] [CrossRef]
- Ganguly, M.; Roychoudhury, A.; Sarkar, S.N.; Sengupta, D.N.; Datta, S.K.; Datta, K. Inducibility of three salinity/abscisic acid-regulated promoters in transgenic rice with gusA reporter gene. Plant Cell Rep. 2011, 30, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Lou, Q.; Zhang, X.; Jiao, S.; Liu, Y.; Wang, Y. Construction of Flower-specific Chimeric Promoters and Analysis of Their Activities in Transgenic Torenia. Plant Mol. Biol. Rep. 2013, 32, 234–245. [Google Scholar] [CrossRef]
- Zhu, Z.; Gao, J.; Yang, J.X.; Wang, X.Y.; Ren, G.D.; Ding, Y.L.; Kuai, B.K. Synthetic promoters consisting of defined cis-acting elements link multiple signaling pathways to probenazole-inducible system. J. Zhejiang Univ. Sci. B 2015, 16, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhu, M.; Ye, R.; Liu, Z.; Zhou, F.; Chen, H.; Lin, Y. Novel green tissue-specific synthetic promoters and cis-regulatory elements in rice. Sci. Rep. 2015, 5, 18256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Song, B.; Zhang, C.; Ou, Y.; Xie, C.; Liu, J. Construction and functional characteristics of tuber-specific and cold-inducible chimeric promoters in potato. Plant Cell Rep. 2015, 27, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Fujita, Y.; Katsura, K.; Maruyama, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulation of ABI3- and ABA-responsive genes including RD29B and RD29A in seeds, germinating embryos, and seedlings of Arabidopsis. Plant Mol. Biol. 2006, 60, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F. Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, N.; Sarkeshik, A.; Nito, K.; Park, S.Y.; Wang, A.; Carvalho, P.C.; Lee, S.; Caddell, D.F.; Cutler, S.R.; Chory, J. PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis. Plant J. 2010, 61, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Dorosh, L.; Kharenko, O.A.; Rajagopalan, N.; Loewen, M.C. Molecular Mechanisms in the Activation ofAbscisic Acid Receptor PYR1. PLoS Comput. Biol. 2013, 9, e1003114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, X.Y.; Li, D.K.; Sun, Y.N.; Li, Y.; Luo, Q.; Liu, Z.B.; Wang, J.M.; Li, X.F.; Zhang, H.; et al. CARK1 mediates ABA signaling by phosphorylation of ABA receptors. Cell Discov. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Guzman, M.; Pizzio, G.A.; Antoni, R.; Vera-Sirera, F.; Merilo, E.; Bassel, G.W.; Fernandez, M.A.; Holdsworth, M.J.; Perez-Amador, M.A.; Kollist, H. Arabidopsis PYR/PYL/RCAR Receptors Play a Major Role in Quantitative Regulation of Stomatal Aperture and Transcriptional Response to Abscisic Acid. Plant Cell 2012, 24, 2483–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.; Kim, T.H. Production of ABA responses requires both the nuclear and cytoplasmic functional involvement of PYR1. Biochem. Biophys. Res. Commun. 2017, 484, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Bihmidine, S.; Lin, J.; Stone, J.M.; Awada, T.; Specht, J.E.; Clemente, T.E. Activity of the Arabidopsis RD29A and RD29B promoter elements in soybean under water stress. Planta 2013, 237, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Jorna, M.L.; Brinkhorst-van der Swan, D.L.C.; Karssen, C.M. The isolation of abscisic acid (ABA) deficient mutants by selection of induced revertants in non-germinating gibberellin sensitive lines of Arabidopsis thaliana (L.) heynh. Theoretical Appl. Genet. 1982, 61, 385–393. [Google Scholar]
- Virlouvet, L.; Ding, Y.; Fujii, H.; Avramova, Z.; Fromm, M. ABA signaling is necessary but not sufficient for RD29B transcriptional memory during successive dehydration stresses in Arabidopsis thaliana. Plant J. 2014, 79, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 2006, 18, 1292–1309. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Mizoi, J.; Yoshida, T.; Fujita, Y.; Nakajima, J.; Ohori, T.; Todaka, D.; Nakashima, K.; Hirayama, T.; Shinozaki, K. An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis. Plant Cell Physiol. 2011, 52, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.Y.; Hu, Y.F.; Li, J.J.; Yao, X.; Liu, K.D. BnaABF2, a bZIP transcription factor from rapeseed (Brassica napus L.), enhances drought and salt tolerance in transgenic Arabidopsis. Bot Stud. 2016, 57, 12. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, M.J.; Finch-Savage, W.E.; Grappin, P.; Job, D. Post-genomics dissection of seed dormancy and germination. Trends Plant Sci. 2008, 13, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, Y.; Yan, S.; Li, H.; He, S.; Huang, M.; Li, L. ABA-mediated inhibition of seed germination is associated with ribosomal DNA chromatin condensation, decreased transcription, and ribosomal RNA gene hypoacetylation. Plant Mol. Biol. 2012, 79, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, Z.; Gao, J.; Gong, Z. Abscisic acid inhibits root growth in Arabidopsis through ethylene biosynthesis. Plant J. 2014, 79, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep. 2006, 25, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiologia Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant. Methods 2005, 1, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | 35S:CARK1 | Ap:CARK1 | Dp:CARK1 | ANDp:CARK1 | Dp:RCAR11-Ap:CARK1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| #2 | #8 | #10 | #12 | #3 | #13 | #4 | #5 | |||
| 1 | 0 | 6% | 2% | 0 | 0 | 0 | 80% | 95% | 100% | 90% |
| 2 | 4% | 0 | 11% | 7.5% | 0 | 80% | 9% | 90% | 100% | 55.5% |
| 3 | 0 | 16.7% | 100% | 10% | 0 | 0 | 20% | 85% | 90% | 100% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, H.; Li, X.; Chen, S.; Zhang, M.; Liu, Z.; Wang, J.; Li, X.; Yang, Y. The Expression of CARK1 or RCAR11 Driven by Synthetic Promoters Increases Drought Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2018, 19, 1945. https://doi.org/10.3390/ijms19071945
Ge H, Li X, Chen S, Zhang M, Liu Z, Wang J, Li X, Yang Y. The Expression of CARK1 or RCAR11 Driven by Synthetic Promoters Increases Drought Tolerance in Arabidopsis thaliana. International Journal of Molecular Sciences. 2018; 19(7):1945. https://doi.org/10.3390/ijms19071945
Chicago/Turabian StyleGe, Hu, Xiaoyi Li, Shisi Chen, Mengru Zhang, Zhibin Liu, Jianmei Wang, Xufeng Li, and Yi Yang. 2018. "The Expression of CARK1 or RCAR11 Driven by Synthetic Promoters Increases Drought Tolerance in Arabidopsis thaliana" International Journal of Molecular Sciences 19, no. 7: 1945. https://doi.org/10.3390/ijms19071945
APA StyleGe, H., Li, X., Chen, S., Zhang, M., Liu, Z., Wang, J., Li, X., & Yang, Y. (2018). The Expression of CARK1 or RCAR11 Driven by Synthetic Promoters Increases Drought Tolerance in Arabidopsis thaliana. International Journal of Molecular Sciences, 19(7), 1945. https://doi.org/10.3390/ijms19071945