Dietary Conjugated Linoleic Acid-Enriched Cheeses Influence the Levels of Circulating n-3 Highly Unsaturated Fatty Acids in Humans

 , ,
, ,  ,
, 
Abstract
:1. Introduction
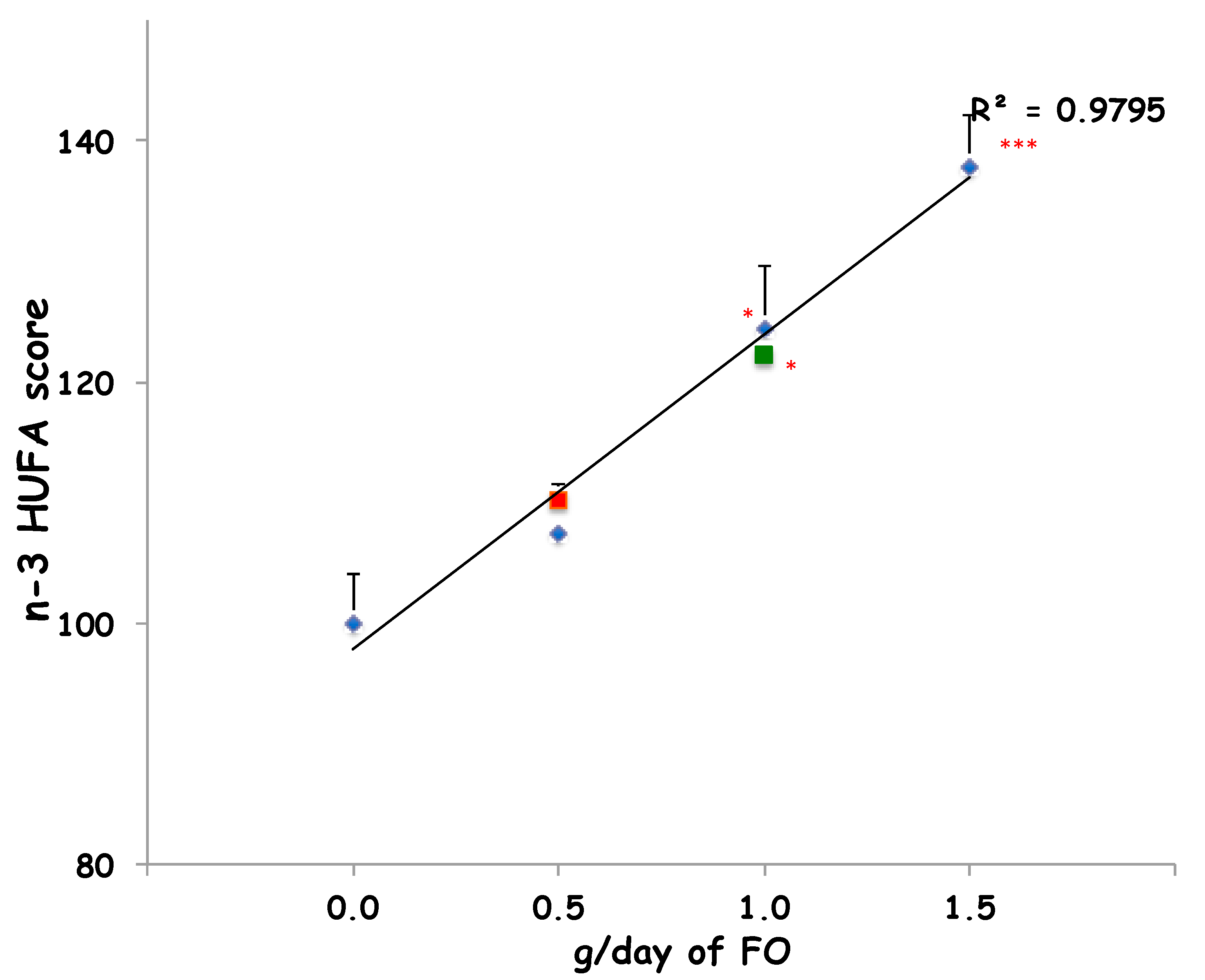
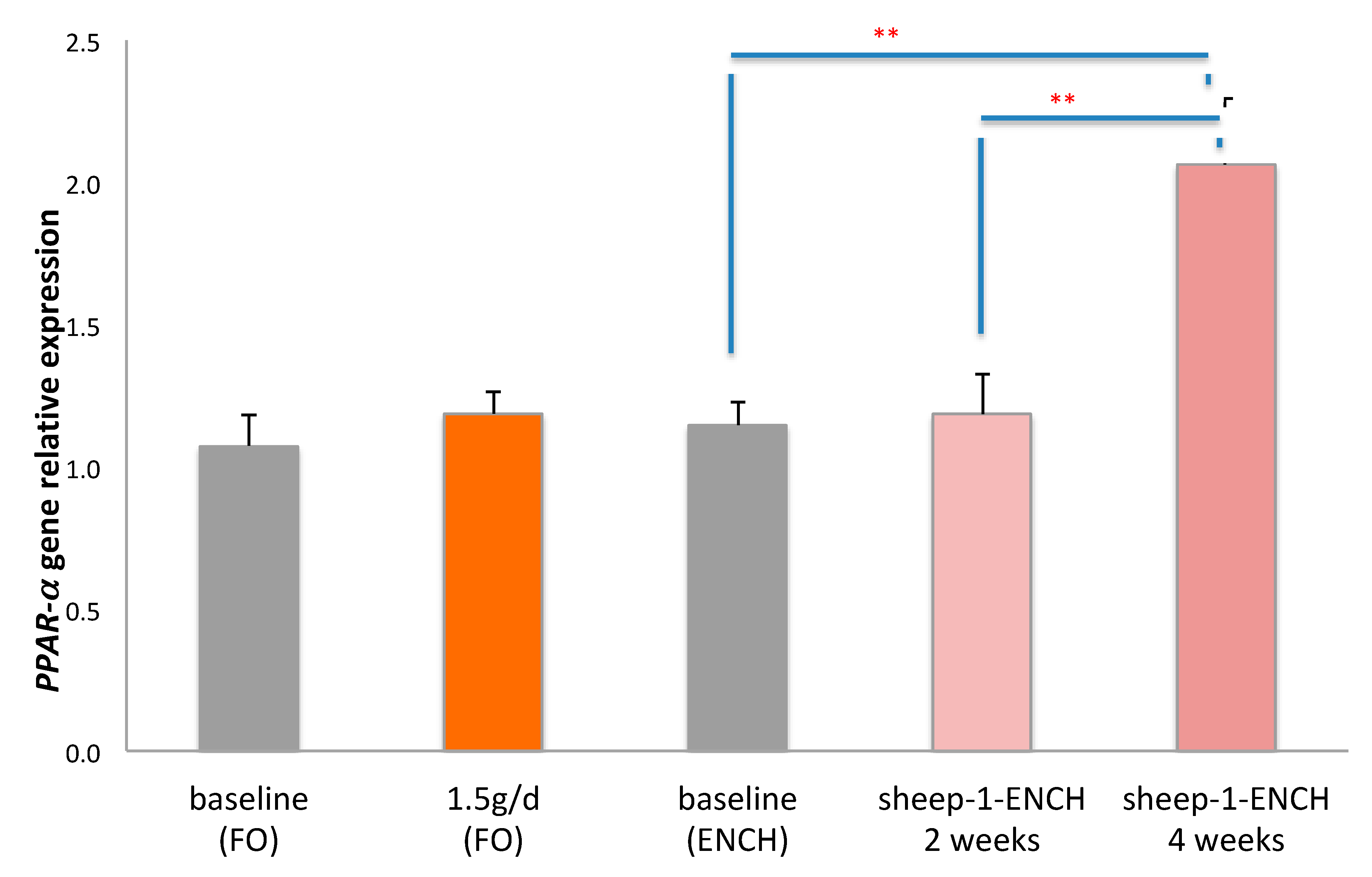
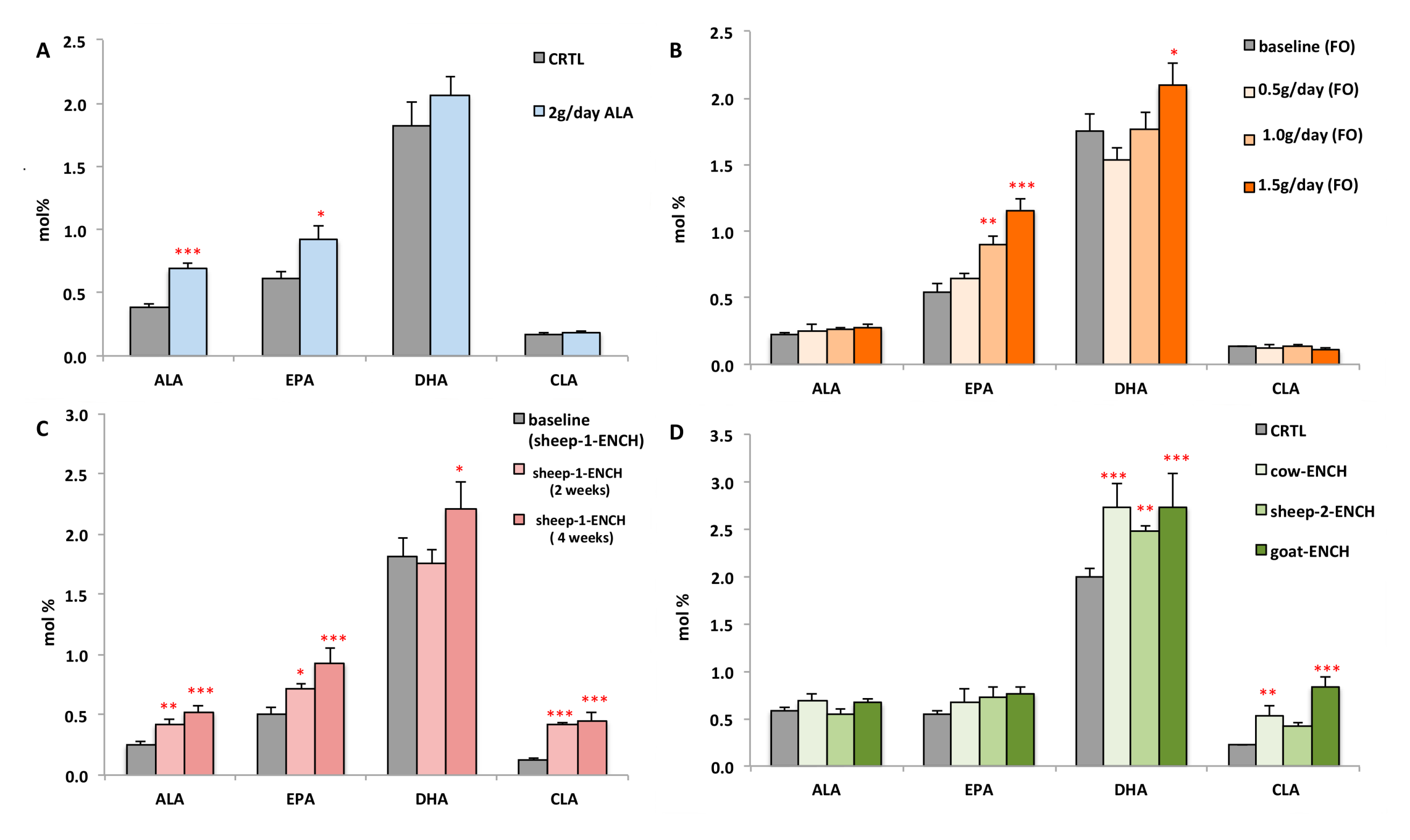
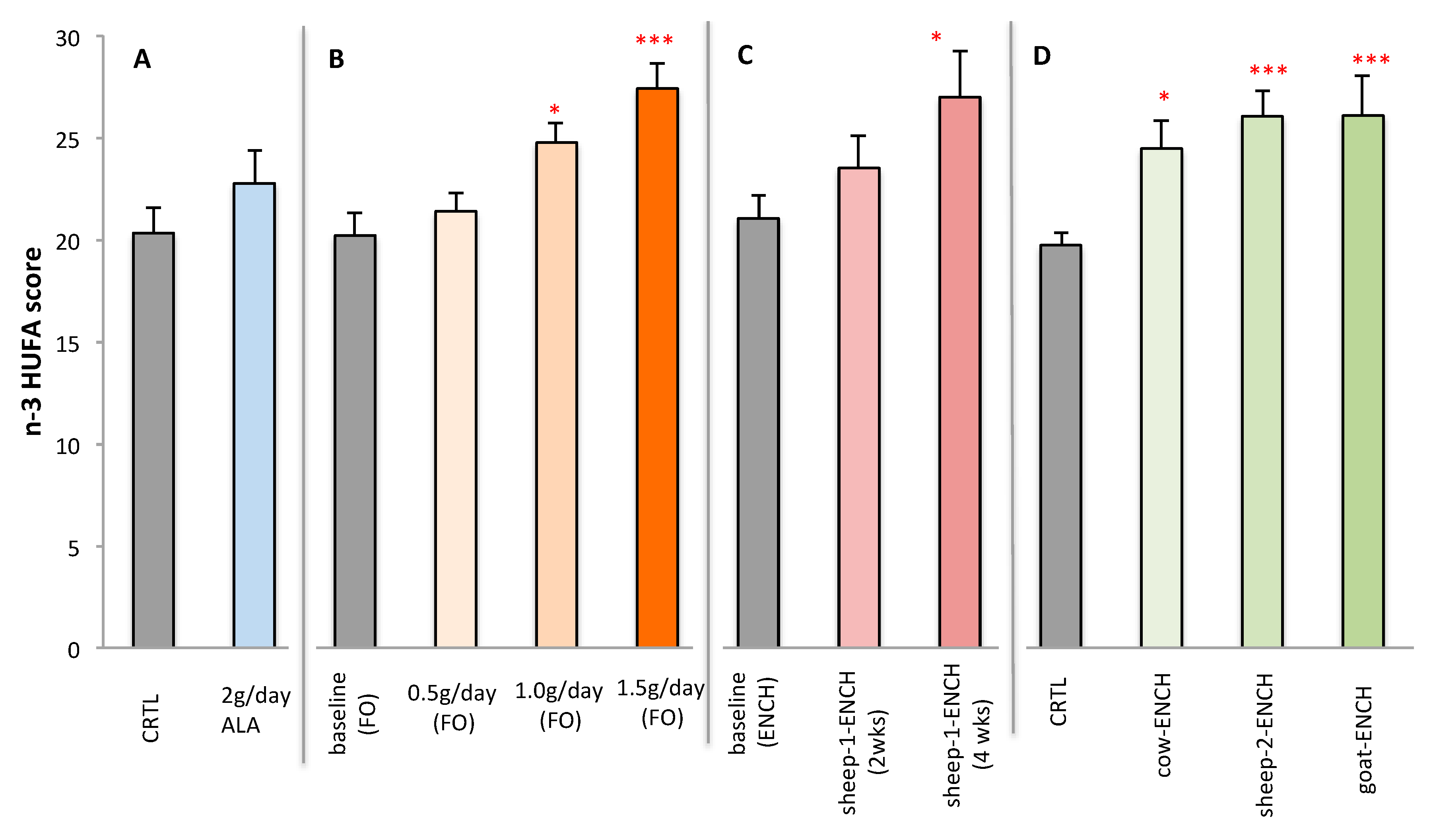
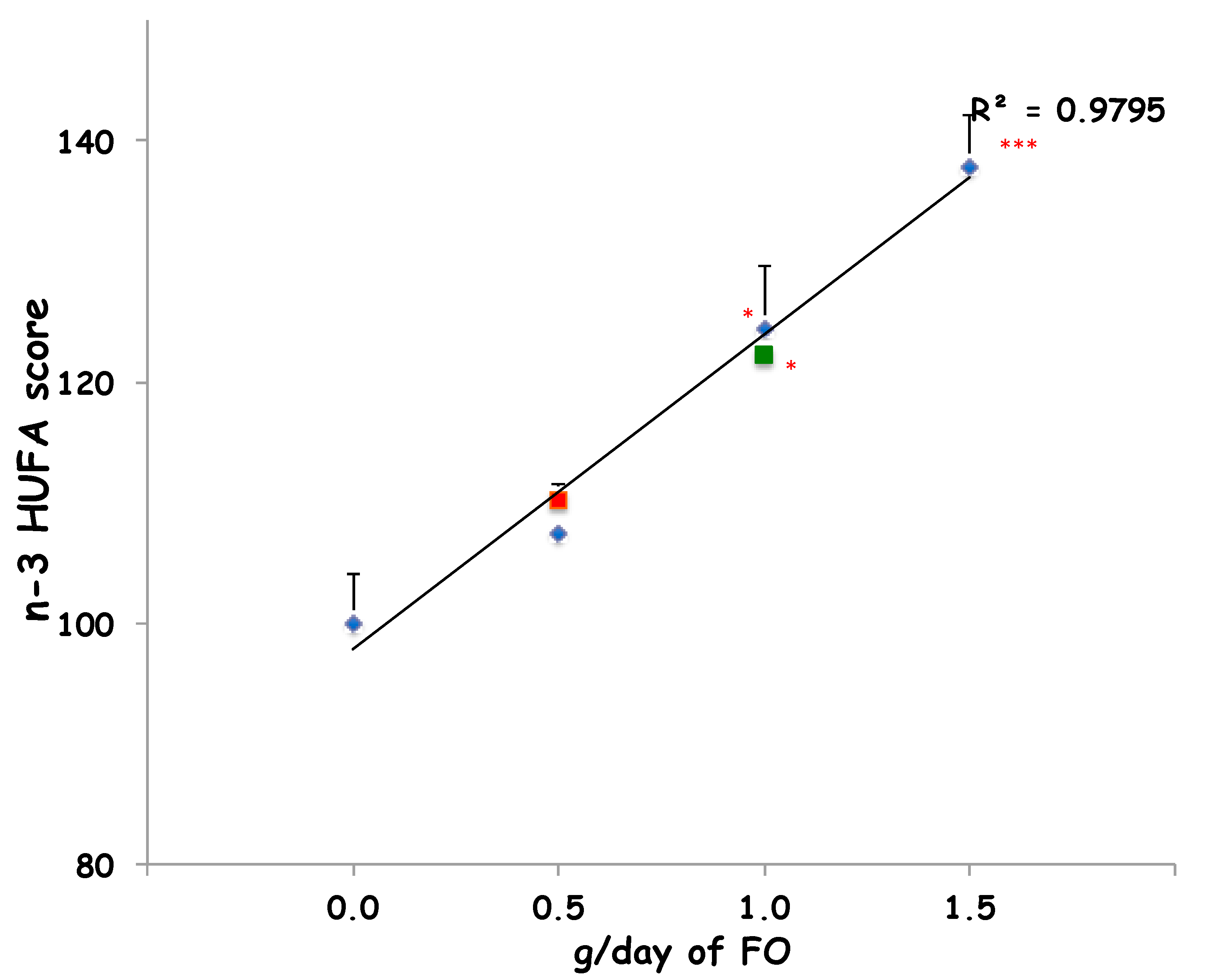
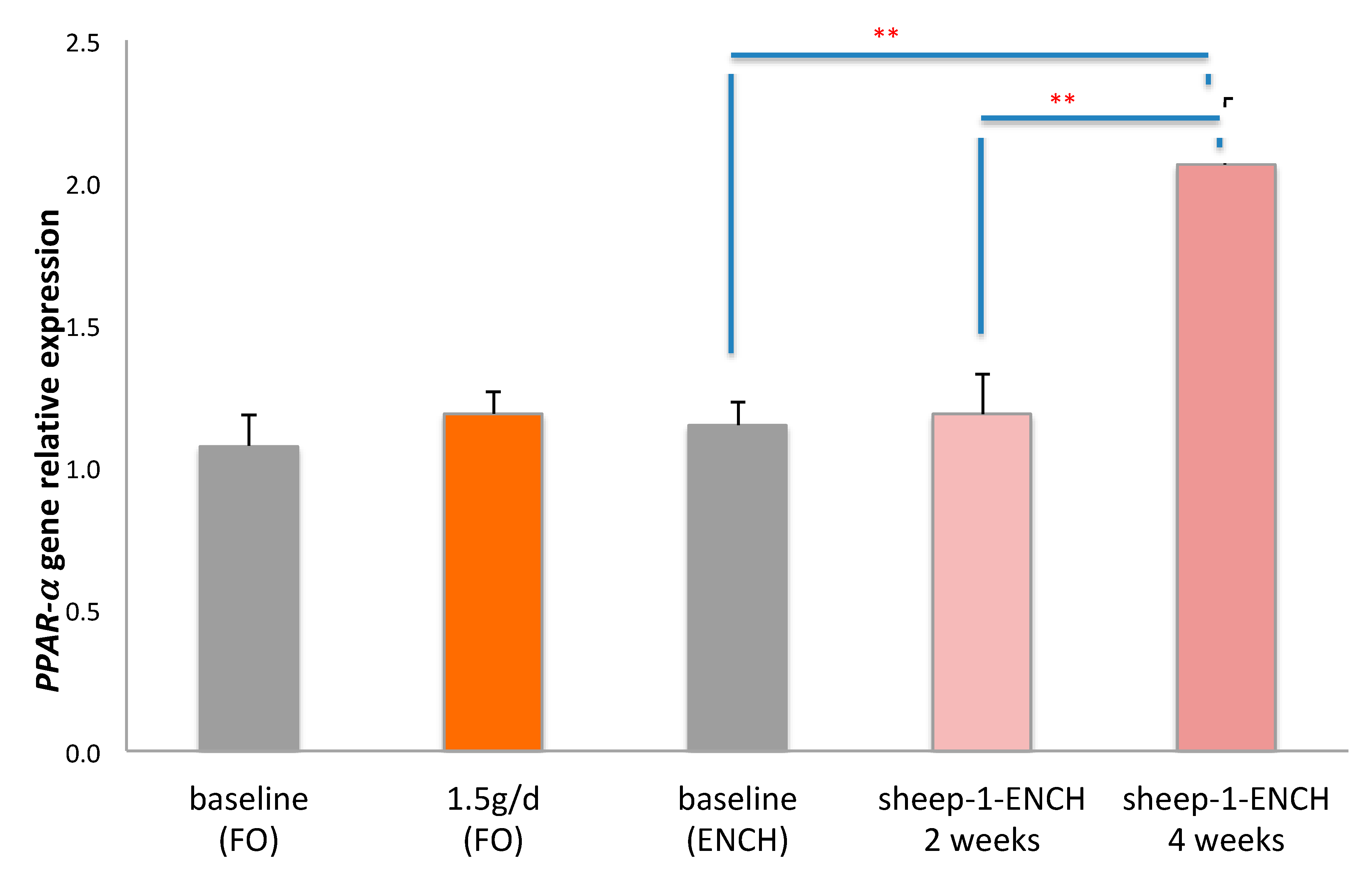
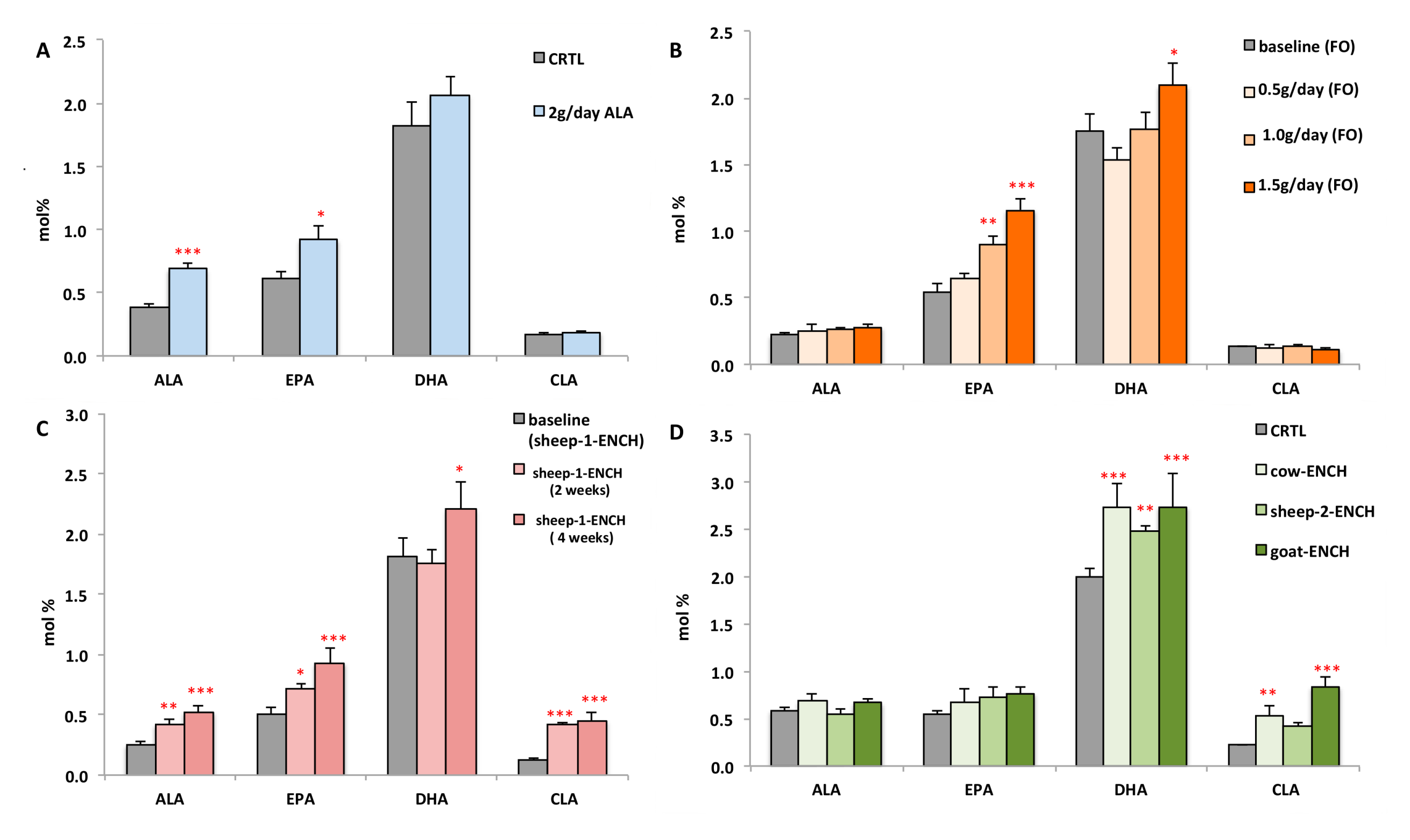
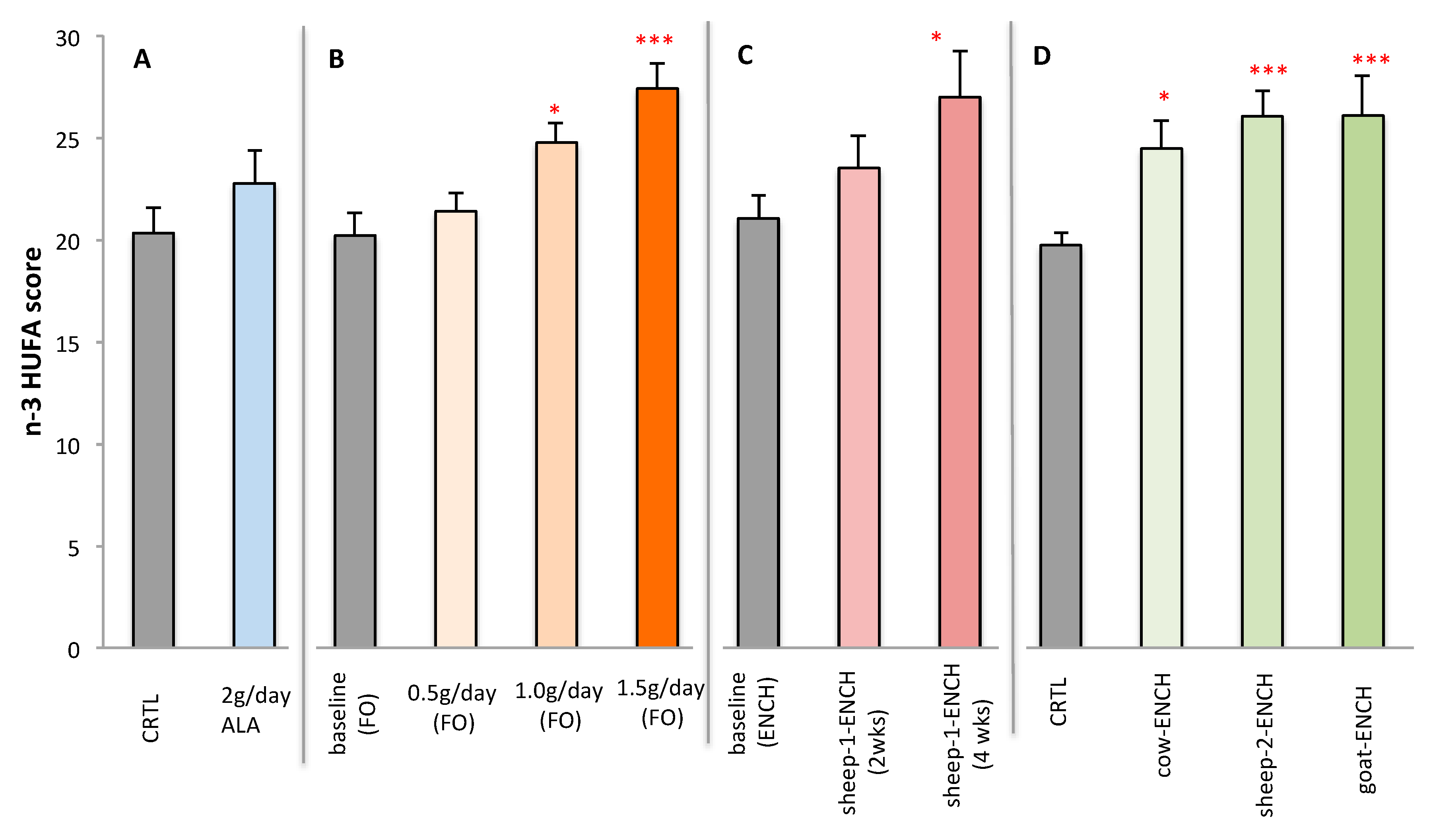
2. Results
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Blood Sampling
4.3. Study Design
- Cow’s milk (cow-ENCH); supplied by Teagasc, Moorepark, Fermoy, Co. Cork, Ireland. CLA was increased in the milk by supplementation with linseed and sunflower oils (unpublished data).
- Sheep’s milk (sheep-2-ENCH); Pecorino Sardo PDO cheese supplied by AGRIS, Bonassai, Olmedo, Italy, which was enriched in CLA by using a sheep feeding regimen characterized by extensive grazing of grass–legume pastures and low levels of supplements [67].
- Goat’s milk (goat-ENCH); supplied by INRA, Clermont-Ferrand-Theix, France. The CLA-enriched goat spread-type cheese was manufactured by Actalia (Surgères, France). The milk was enriched in CLA as a result of an alfalfa hay-based diet supplemented with sunflower oil (130 g/day) and Vitamin E (50 g/day) [68].
4.4. Lipid Analyses
4.5. Quantitative Analysis of PPAR-α mRNA
5. Statistical Analyses
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Eslick, G.D.; Howe, P.R.; Smith, C.; Priest, R.; Bensoussan, A. Benefits of fish oil supplementation in hyperlipidemia: A systematic review and meta-analysis. Int. J. Cardiol. 2009, 136, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M.; Sprecher, H.; Hachey, D.; Edmond, J.; Anderson, R.E. Neonatal polyunsaturated fatty acid metabolism. Lipids 1999, 34, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The omega-6/omega-3 fatty acid ratio, genetic variation, and cardiovascular disease. Asia Pac. J. Clin. Nutr. 2008, 17, 131–134. [Google Scholar] [PubMed]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.A.; Muhlhausler, B.; Makrides, M. Conversion of linoleic acid and α-linolenic acid to long-chain polyunsaturated fatty acids (LCPUFAs), with a focus on pregnancy, lactation and the first 2 years of life. Matern. Child Nutr. 2011, 7, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Mest, H.J.; Beitz, J.; Heinroth, I.; Block, H.U.; Forster, W. The influence of linseed oil diet on fatty acid pattern in phospholipids and thromboxane formation in platelets in man. Klin. Wochenschr. 1983, 61, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, O.; Takahashi, M.; Shigematsu, T.; Shimamura, K.; Kimura, J.; Ezaki, H.; Gotoh, T. Long-term effects of dietary α-linolenic acid from perilla oil on serum fatty acids composition and on the risk factors of coronary heart disease in japanese elderly subjects. J. Nutr. Sci. Vitaminol. 1999, 45, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Gerster, H. Can adults adequately convert α-linolenic acid (18:3n-3) to eicosapentaenoic acid (20:5n-3) and docosahexaenoic acid (22:6n-3)? Int. J. Vitam. Nutr. Res. 1998, 68, 159–173. [Google Scholar] [PubMed]
- Li, D.; Sinclair, A.; Wilson, A.; Nakkote, S.; Kelly, F.; Abedin, L.; Mann, N.; Turner, A. The effect of dietary α-linolenic acid on thrombotic risk factors in vegetarian men. Am. J. Clin. Nutr. 1999, 69, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Mantzioris, E.; James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary substitution with an α-linolenic acid-rich vegetable oil increases eicosapentaenoic acid concentrations in tissues. Am. J. Clin. Nutr. 1994, 59, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plourde, M.; Cunnane, S.C. Extremely limited synthesis of long chain polyunsaturates in adults: Implications for their dietary essentiality and use as supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 619–634. [Google Scholar] [CrossRef] [PubMed]
- Pawlosky, R.J.; Hibbeln, J.R.; Novotny, J.A.; Salem, N., Jr. Physiological compartmental analysis of α-linolenic acid metabolism in adult humans. J. Lipid Res. 2001, 42, 1257–1265. [Google Scholar] [PubMed]
- Ferdinandusse, S.; Denis, S.; Dacremont, G.; Wanders, R.J. Studies on the metabolic fate of n-3 polyunsaturated fatty acids. J. Lipid Res. 2003, 44, 1992–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murru, E.; Banni, S.; Carta, G. Nutritional properties of dietary omega-3-enriched phospholipids. BioMed Res. Int. 2013, 2013, 965417. [Google Scholar] [CrossRef] [PubMed]
- Chapkin, R.S.; Wang, N.; Fan, Y.Y.; Lupton, J.R.; Prior, I.A. Docosahexaenoic acid alters the size and distribution of cell surface microdomains. Biochim. Biophys. Acta 2008, 1778, 466–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- Jump, D.B. The biochemistry of n-3 polyunsaturated fatty acids. J. Biol. Chem. 2002, 277, 8755–8758. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Nettleton, J.A.; Folsom, A.R.; Investigators, A.S. Plasma fatty acid composition and incident heart failure in middle-aged adults: The atherosclerosis risk in communities (ARIC) study. Am. Heart J. 2008, 156, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, E.; Freund-Levi, Y. Immunomodulation of microglia by docosahexaenoic acid and eicosapentaenoic acid. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Dietary omega 3 fatty acids and the developing brain. Brain Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Periz, A.; Horrillo, R.; Ferre, N.; Gronert, K.; Dong, B.; Moran-Salvador, E.; Titos, E.; Martinez-Clemente, M.; Lopez-Parra, M.; Arroyo, V.; et al. Obesity-induced insulin resistance and hepatic steatosis are alleviated by omega-3 fatty acids: A role for resolvins and protectins. FASEB J. 2009, 23, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Importance of the ratio of omega-6/omega-3 essential fatty acids: Evolutionary aspects. World Rev. Nutr. Diet. 2003, 92, 1–22. [Google Scholar] [PubMed]
- Simopoulos, A.P. Evolutionary aspects of diet: The omega-6/omega-3 ratio and the brain. Mol. Neurobiol. 2011, 44, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; American Heart Association. Nutrition Committee. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Grieger, J.A.; Etherton, T.D. Dietary reference intakes for DHA and EPA. Prostaglandins Leukot. Essent. Fatty Acids 2009, 81, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D. The percentage of n-3 highly unsaturated fatty acids in total hufa as a biomarker for omega-3 fatty acid status in tissues. Lipids 2008, 43, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Mele, M.; Contarini, G.; Cercaci, L.; Serra, A.; Buccioni, A.; Povolo, M.; Conte, G.; Funaro, A.; Banni, S.; Lercker, G.; et al. Enrichment of pecorino cheese with conjugated linoleic acid by feeding dairy ewes with extruded linseed: Effect on fatty acid and triglycerides composition and on oxidative stability. Int. Dairy J. 2011, 21, 365–372. [Google Scholar] [CrossRef]
- Vasta, V.; Mele, M.; Serra, A.; Scerra, M.; Luciano, G.; Lanza, M.; Priolo, A. Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins. J. Anim. Sci. 2009, 87, 2674–2684. [Google Scholar] [CrossRef] [PubMed]
- Cabiddu, A.; Addis, M.; Fiori, M.; Spada, S.; Decandia, M.; Molle, G. Pros and cons of the supplementation with oilseed enrichedconcentrates on milk fatty acid profile of dairy sheep grazing mediterranean pastures. Small Rumin. Res. 2017, 146, 63–72. [Google Scholar] [CrossRef]
- Ferlay, A.; Bernard, L.; Meynadier, A.; Malpuech-Brugère, C. Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: A review. Biochimie 2017, 141, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Banni, S.; Martin, J.C. Conjugated linoleic acid and metabolites. In Trans Fatty Acid in Human Nutrition; Sebedio, J.L., Christie, W.W., Eds.; Oily Press: Dundee, UK, 1998; pp. 261–302. [Google Scholar]
- Kepler, C.R.; Hirons, K.P.; McNeill, J.J.; Tove, S.B. Intermediates and products of the biohydrogenation of linoleic acid by Butyrinvibrio fibrisolvens. J. Biol. Chem. 1966, 241, 1350–1354. [Google Scholar] [PubMed]
- Kepler, C.R.; Tucker, W.P.; Tove, S.B. Biohydrogenation of unsaturated fatty acids. IV. Substrate specificity and inhibition of linoleate delta-12-cis, delta-11-trans-isomerase from Butyrivibrio fibrisolvens. J. Biol. Chem. 1970, 245, 3612–3620. [Google Scholar] [PubMed]
- Adlof, R.O.; Duval, S.; Emken, E.A. Biosynthesis of conjugated linoleic acid in humans. Lipids 2000, 35, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Kuhnt, K.; Kraft, J.; Moeckel, P.; Jahreis, G. Trans-11-18:1 is effectively delta9-desaturated compared with trans-12-18:1 in humans. Br. J. Nutr. 2006, 95, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Banni, S. Conjugated linoleic acid metabolism. Curr. Opin. Lipidol. 2002, 13, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Pintus, S.; Murru, E.; Carta, G.; Cordeddu, L.; Batetta, B.; Accossu, S.; Pistis, D.; Uda, S.; Elena Ghiani, M.; Mele, M.; et al. Sheep cheese naturally enriched in α-linolenic, conjugated linoleic and vaccenic acids improves the lipid profile and reduces anandamide in the plasma of hypercholesterolaemic subjects. Br. J. Nutr. 2013, 109, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Ruuska, S.E.; Shaw, N.S.; Dong, D.; Noy, N. Ligand selectivity of the peroxisome proliferator-activated receptor α. Biochemistry 1999, 38, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Sette, S.; Le Donne, C.; Piccinelli, R.; Arcella, D.; Turrini, A.; Leclercq, C.; Group, I.-S.S. The third Italian National Food Consumption Survey, INRAN-SCAI 2005-06—Part 1: Nutrient intakes in Italy. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, T.L.; Southgate, D.A. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Von Schacky, C. The omega-3 index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Carta, G.; Murru, E.; Lopes, P.A.; Martins, S.V.; Prates, J.A.; Banni, S. Effects of dietary CLA on n-3 hufa score and n-acylethanolamides biosynthesis in the liver of obese zucker rats. Prostaglandins Leukot. Essent. Fatty Acids 2015, 98, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Moya-Camarena, S.Y.; Vanden Heuvel, J.P.; Blanchard, S.G.; Leesnitzer, L.A.; Belury, M.A. Conjugated linoleic acid is a potent naturally occurring ligand and activator of pparα. J. Lipid Res. 1999, 40, 1426–1433. [Google Scholar] [PubMed]
- Belury, M.A.; Moya-Camarena, S.Y.; Liu, K.L.; Vanden Heuvel, J.P. Dietary conjugated linoleic acid induces peroxisone-specific enzyme accumulation and ornithine decarboxylase activity in mouse liver. J. Nutr. Biochem. 1997, 8, 579–584. [Google Scholar] [CrossRef]
- Attar-Bashi, N.M.; Weisinger, R.S.; Begg, D.P.; Li, D.; Sinclair, A.J. Failure of conjugated linoleic acid supplementation to enhance biosynthesis of docosahexaenoic acid from α-linolenic acid in healthy human volunteers. Prostaglandins Leukot. Essent. Fatty Acids 2007, 76, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Vigano, D.; Guidali, C.; Petrosino, S.; Realini, N.; Rubino, T.; di Marzo, V.; Parolaro, D. Involvement of the endocannabinoid system in phencyclidine-induced cognitive deficits modelling schizophrenia. Int. J. Neuropsychopharmacol. 2009, 12, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T. Efficiency of conversion of α-linolenic acid to long chain n-3 fatty acids in man. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C. Metabolism of α-linolenic acid in humans. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Zamberletti, E.; Piscitelli, F.; de Castro, V.; Murru, E.; Gabaglio, M.; Colucci, P.; Fanali, C.; Prini, P.; Bisogno, T.; Maccarrone, M.; et al. Lifelong imbalanced la/ala intake impairs emotional and cognitive behavior via changes in brain endocannabinoid system. J. Lipid Res. 2017, 58, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, M.U.; O’Reilly, E.J.; Heitmann, B.L.; Pereira, M.A.; Balter, K.; Fraser, G.E.; Goldbourt, U.; Hallmans, G.; Knekt, P.; Liu, S.; et al. Major types of dietary fat and risk of coronary heart disease: A pooled analysis of 11 cohort studies. Am. J. Clin. Nutr. 2009, 89, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic acid: Physiological role, metabolism and nutritional implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
- Biolchini, M.; Murru, E.; Anfora, G.; Loy, F.; Banni, S.; Crnjar, R.; Sollai, G. Fat storage in drosophila suzukii is influenced by different dietary sugars in relation to their palatability. PLoS ONE 2017, 12, e0183173. [Google Scholar] [CrossRef] [PubMed]
- Souza-Mello, V. Peroxisome proliferator-activated receptors as targets to treat non-alcoholic fatty liver disease. World J. Hepatol. 2015, 7, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Cappa, M.; Bizzarri, C.; Petroni, A.; Carta, G.; Cordeddu, L.; Valeriani, M.; Vollono, C.; De Pasquale, L.; Blasevich, M.; Banni, S. A mixture of oleic, erucic and conjugated linoleic acids modulates cerebrospinal fluid inflammatory markers and improve somatosensorial evoked potential in x-linked adrenoleukodystrophy female carriers. J. Inherit. Metab. Dis. 2012, 35, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Saba, F.; Sirigu, A.; Pillai, R.; Caria, P.; Cordeddu, L.; Carta, G.; Murru, E.; Sogos, V.; Banni, S. Downregulation of inflammatory markers by conjugated linoleic acid isomers in human cultured astrocytes. Nutr. Neurosci. 2017, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandusse, S.; Rusch, H.; van Lint, A.E.; Dacremont, G.; Wanders, R.J.; Vreken, P. Stereochemistry of the peroxisomal branched-chain fatty acid α- and β-oxidation systems in patients suffering from different peroxisomal disorders. J. Lipid Res. 2002, 43, 438–444. [Google Scholar] [PubMed]
- Leung, L.K.; Glauert, H.P. Reduction of the concentrations of prostaglandins E2 and F2α, and thromboxane B2 in cultured rat hepatocytes treated with the peroxisome proliferator ciprofibrate. Toxicol. Lett. 1996, 85, 143–149. [Google Scholar] [CrossRef]
- Benjamin, S.; Prakasan, P.; Sreedharan, S.; Wright, A.D.; Spener, F. Pros and cons of CLA consumption: An insight from clinical evidences. Nutr. Metab. 2015, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.L.; Grimm, N.K.; Pariza, M.W. Anticarcinogens from fried ground beef: Heat-altered derivatives of linoleic acid. Carcinogenesis 1987, 8, 1881–1887. [Google Scholar] [CrossRef] [PubMed]
- Whigham, L.; Cook, M.; Atkinson, R. Conjugated linoleic acid: Implications for human health. Pharmacol. Res. 2000, 42, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Pariza, M.; Park, Y.; Cook, M. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res. 2001, 40, 283–298. [Google Scholar] [CrossRef]
- McGuire, M.; McGuire, M. Conjugated linoleic acid (CLA): A ruminant fatty acid with beneficial effects on human health. J. Anim. Sci. 2000, 77, 1–8. [Google Scholar] [CrossRef]
- Addis, M.; Cabiddu, A.; Pinna, G.; Decandia, M.; Piredda, G.; Pirisi, A.; Molle, G. Milk and cheese fatty acid composition of sheep fed different mediterranean forages with particular reference to cla cis-9, trans-11. J. Dairy Sci. 2005, 88, 3443–3454. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat: A review. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Chiang, S.; Gessert, C.; Lowry, O. Colorimetric determination of extracted lipids. An adaptation for microgram amounts of lipids obtained from cerumen. Curr. List Med. Lit. Res. Rep. 1957, 33, 56–113. [Google Scholar]
- Piscitelli, F.; Carta, G.; Bisogno, T.; Murru, E.; Cordeddu, L.; Berge, K.; Tandy, S.; Cohn, J.S.; Griinari, M.; Banni, S.; et al. Effect of dietary krill oil supplementation on the endocannabinoidome of metabolically relevant tissues from high-fat-fed mice. Nutr. Metab. 2011, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banni, S.; Carta, G.; Contini, M.S.; Angioni, E.; Deiana, M.; Dessi, M.A.; Melis, M.P.; Corongiu, F.P. Characterization of conjugated diene fatty acids in milk, dairy products, and lamb tissues. J. Nutr. Biochem. 1996, 7, 150–155. [Google Scholar] [CrossRef]
- Melis, M.P.; Angioni, E.; Carta, G.; Murru, E.; Scanu, P.; Spada, S.; Banni, S. Characterization of conjugated linoleic acid and its metabolities by rp-hplc with diode array detector. Eur. J. Lipid Sci. Technol. 2001, 103, 617–621. [Google Scholar] [CrossRef]
- Batetta, B.; Griinari, M.; Carta, G.; Murru, E.; Ligresti, A.; Cordeddu, L.; Giordano, E.; Sanna, F.; Bisogno, T.; Uda, S.; et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese zucker rats. J. Nutr. 2009, 139, 1495–1501. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA mol % in Plasma | Baseline (Time 0) | Sheep-1-ENCH (2 Weeks) | Sheep-1-ENCH (4 Weeks) | |||
|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | |
| 14:0 | 1.25 | 0.24 | 1.16 | 0.16 | 1.36 | 0.26 |
| 16:0 (PA) | 30.28 | 0.54 | 29.52 | 0.34 | 24.72 | 0.63 *** |
| 17:0 | 7.27 | 0.22 | 7.49 | 0.12 | 6.02 | 0.21 *** |
| 18:0 | 1.03 | 0.06 | 1.16 | 0.06 | 1.03 | 0.08 |
| 16:1 | 2.54 | 0.15 | 2.53 | 0.22 | 2.32 | 0.30 |
| 18:1n-9 (OA) | 18.20 | 0.60 | 18.62 | 1.53 | 19.95 | 1.03 |
| 18:3n-3 (ALA) | 0.25 | 0.03 | 0.42 | 0.04 ** | 0.52 | 0.05 *** |
| 18:4n-3 | 0.03 | 0.01 | 0.03 | 0.00 | 0.03 | 0.00 |
| 20:5n-3 (EPA) | 0.51 | 0.05 | 0.71 | 0.05 * | 0.93 | 0.13 *** |
| 22:6n-3 (DHA) | 1.81 | 0.15 | 1.75 | 0.11 | 2.51 | 0.15 * |
| 18:2n-6 (LA) | 27.05 | 0.85 | 29.68 | 1.59 | 31.63 | 0.62 |
| 18:3n-6 | 0.54 | 0.05 | 0.49 | 0.03 | 0.49 | 0.04 |
| 20:3n-6 | 1.52 | 0.12 | 1.53 | 0.19 | 1.64 | 0.13 |
| 20:4n-6 (ARA) | 6.79 | 0.37 | 6.46 | 0.43 | 7.11 | 0.38 |
| 22:4n-6 | 0.14 | 0.02 | 0.11 | 0.01 | 0.11 | 0.00 |
| 18:2c9t11 (CLA) | 0.13 | 0.10 | 0.42 | 0.23 *** | 0.45 | 0.07 *** |
| FA mol % in Plasma | CRTL | Cow-ENCH | Sheep-2-ENCH | Goat-ENCH | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| 14:0 | 0.64 | 0.02 | 1.31 | 0.12 *** | 0.66 | 0.06 | 0.58 | 0.07 |
| 15:0 | 0.22 | 0.00 | 0.42 | 0.03 *** | 0.17 | 0.01 * | 0.22 | 0.02 |
| 16:0 (PA) | 11.53 | 0.25 | 11.26 | 2.27 | 11.13 | 0.30 | 11.93 | 0.85 |
| 17:0 | 0.50 | 0.02 | 0.38 | 0.04 | 0.34 | 0.02 ** | 0.50 | 0.05 |
| 18:0 | 9.03 | 0.29 | 6.90 | 0.42 | 6.95 | 0.21 * | 7.89 | 0.78 |
| 14:1 | 0.09 | 0.01 | 0.07 | 0.01 | 0.05 | 0.00 | 0.07 | 0.01 |
| 16:1 | 1.82 | 0.12 | 2.07 | 0.24 | 1.77 | 0.06 | 1.57 | 0.11 |
| 18:1n-9 (OA) | 26.31 | 1.02 | 29.96 | 1.90 | 27.48 | 1.30 | 26.70 | 1.19 |
| 18:3n-3 (ALA) | 0.58 | 0.03 | 0.69 | 0.04 | 0.54 | 0.05 | 0.67 | 0.08 |
| 18:4n-3 | 0.02 | 0.00 | 0.02 | 0.01 | 0.02 | 0.00 | 0.02 | 0.00 |
| 20:5n-3 (EPA) | 0.54 | 0.03 | 0.67 | 0.07 | 0.73 | 0.11 | 0.76 | 0.14 |
| 22:6n-3 (DHA) | 1.99 | 0.09 | 2.73 | 0.33 *** | 2.47 | 0.08 ** | 2.74 | 0.28 *** |
| 18:2n-6 (LA) | 35.58 | 0.76 | 40.73 | 2.47 * | 38.67 | 1.56 | 34.32 | 1.24 |
| 18:3n-6 | 0.41 | 0.03 | 0.50 | 0.06 | 0.39 | 0.03 | 0.37 | 0.05 |
| 20:3n-6 | 1.89 | 0.08 | 2.32 | 0.11 * | 1.58 | 0.09 | 1.96 | 0.16 |
| 20:4n-6 (ARA) | 8.06 | 0.35 | 7.79 | 0.85 | 7.20 | 0.31 | 7.52 | 0.33 |
| 22:4n-6 | 0.27 | 0.01 | 0.31 | 0.01 | 0.23 | 0.02 | 0.27 | 0.03 |
| 18:1t11 (VA) | 0.36 | 0.02 | 0.69 | 0.07 ** | 0.57 | 0.06 * | 1.03 | 0.15 *** |
| 18:2c9t11 (CLA) | 0.22 | 0.01 | 0.53 | 0.11 ** | 0.41 | 0.04 | 0.85 | 0.11 *** |
| Blood Parameters | Mean | SEM |
|---|---|---|
| Age (years) | 30–40 | |
| BMI (kg/m2) | 20–26 | |
| Glycemia (mg/dL) | 85.5 | 1.5 |
| Triglyceride (mg/dL) | 80.3 | 6.3 |
| Total cholesterol (C) (mg/dL) | 190.5 | 6.7 |
| HDL-C (mg/dL) | 60.1 | 1.9 |
| LDL-C (mg/dL) | 113.8 | 6.1 |
| Total C/HDL-C ratio | 3.3 | 0.1 |
| g FA/50 g of Cheese | Mean of CRTL Cheeses | Cow-ENCH | Sheep-2-ENCH | Goat-ENCH |
|---|---|---|---|---|
| short chain (c4–c10) | 2.30 | 1.55 | 2.06 | 2.15 |
| 12:0 | 0.63 | 0.38 | 0.50 | 0.43 |
| 14:0 | 1.33 | 0.79 | 1.06 | 1.11 |
| 15:0 | 0.14 | 0.14 | 0.13 | 0.13 |
| 16:0 (PA) | 2.98 | 2.30 | 2.16 | 2.27 |
| 17:0 | 0.09 | 0.05 | 0.08 | 0.07 |
| 18:0 | 1.02 | 1.35 | 1.20 | 1.08 |
| 14:1 | 0.05 | 0.04 | 0.02 | 0.01 |
| 16:1 | 0.10 | 0.11 | 0.07 | 0.05 |
| 18:1n-9 (OA) | 1.85 | 2.59 | 1.99 | 1.98 |
| 18:2n-6 (LA) | 0.24 | 0.23 | 0.20 | 0.41 |
| 18:3n-3 (ALA) | 0.09 | 0.15 | 0.13 | 0.04 |
| 18:1t11 (VA) | 0.19 | 0.86 | 0.61 | 1.06 |
| 18:2c9t11 (CLA) | 0.07 | 0.30 | 0.31 | 0.48 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murru, E.; Carta, G.; Cordeddu, L.; Melis, M.P.; Desogus, E.; Ansar, H.; Chilliard, Y.; Ferlay, A.; Stanton, C.; Coakley, M.; et al. Dietary Conjugated Linoleic Acid-Enriched Cheeses Influence the Levels of Circulating n-3 Highly Unsaturated Fatty Acids in Humans. Int. J. Mol. Sci. 2018, 19, 1730. https://doi.org/10.3390/ijms19061730
Murru E, Carta G, Cordeddu L, Melis MP, Desogus E, Ansar H, Chilliard Y, Ferlay A, Stanton C, Coakley M, et al. Dietary Conjugated Linoleic Acid-Enriched Cheeses Influence the Levels of Circulating n-3 Highly Unsaturated Fatty Acids in Humans. International Journal of Molecular Sciences. 2018; 19(6):1730. https://doi.org/10.3390/ijms19061730
Chicago/Turabian StyleMurru, Elisabetta, Gianfranca Carta, Lina Cordeddu, Maria Paola Melis, Erika Desogus, Hastimansooreh Ansar, Yves Chilliard, Anne Ferlay, Catherine Stanton, Mairéad Coakley, and et al. 2018. "Dietary Conjugated Linoleic Acid-Enriched Cheeses Influence the Levels of Circulating n-3 Highly Unsaturated Fatty Acids in Humans" International Journal of Molecular Sciences 19, no. 6: 1730. https://doi.org/10.3390/ijms19061730
APA StyleMurru, E., Carta, G., Cordeddu, L., Melis, M. P., Desogus, E., Ansar, H., Chilliard, Y., Ferlay, A., Stanton, C., Coakley, M., Ross, R. P., Piredda, G., Addis, M., Mele, M. C., Cannelli, G., Banni, S., & Manca, C. (2018). Dietary Conjugated Linoleic Acid-Enriched Cheeses Influence the Levels of Circulating n-3 Highly Unsaturated Fatty Acids in Humans. International Journal of Molecular Sciences, 19(6), 1730. https://doi.org/10.3390/ijms19061730