Is There a Role for the Non-Helicobacter pylori Bacteria in the Risk of Developing Gastric Cancer?

{kind=link}
Abstract
:1. Introduction
2. Is H. pylori a Risk Factor or a True Carcinogen?
3. Gastric Microbiota and its Influence on Gastric Carcinogenesis
Author Contributions
Conflicts of Interest
References
- International Agency for Research on Cancer. IARC Monographs on the evaluation of carcinogenic risk to humans. In Schistosomaotes, Liver Flukes and Helicobacter pylori; IARC: Lyon, France, 1994; Volume 61, pp. 12–41. [Google Scholar]
- European Helicobacter pylori study group. Current European concepts in the management of Helicobacter pylori infection—The Maastricht consensus report. Gut 1997, 41, 81–83. [Google Scholar]
- Hooi, J.K.Y.; Lai, W.Y.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 4204–4229. [Google Scholar] [CrossRef] [PubMed]
- Peek, R.M.; Crabtree, J.E. Helicobacter pylori and gastric neoplasia. J. Pathol. 2006, 208, 2332–2348. [Google Scholar] [CrossRef] [PubMed]
- Yakirevich, E.; Resnick, M.B. Pathology of gastric cancer and its precursor lesions. Gastroenterol. Clin. N. Am. 2013, 42, 2612–2684. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.Y.; Tung, S.Y.; Pan, H.Y.; Yen, C.W.; Xu, H.W.; Lin, Y.J.; Deng, Y.F.; Hsu, W.T.; Wu, C.S.; Li, C. Increased abundance of Clostridium and Fusobacterium in gastric microbiota of patients with gastric cancer in Taiwan. Sci. Rep. 2018, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A. Time trends of mortality from gastric cancer in Europe. Dig. Dis. Sci. 2011, 56, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A. Temporal trends and geographical variations of peptic ulcer disease. Aliment. Pharmacol. Ther. 1995, 9 (Suppl. 2), 3–12. [Google Scholar] [PubMed]
- Khalifa, M.M.; Sharaf, R.R.; Aziz, R.K. Helicobacter pylori: A poor man’s gut pathogen? Gut Pathog. 2010, 2, 2. [Google Scholar] [CrossRef] [PubMed]
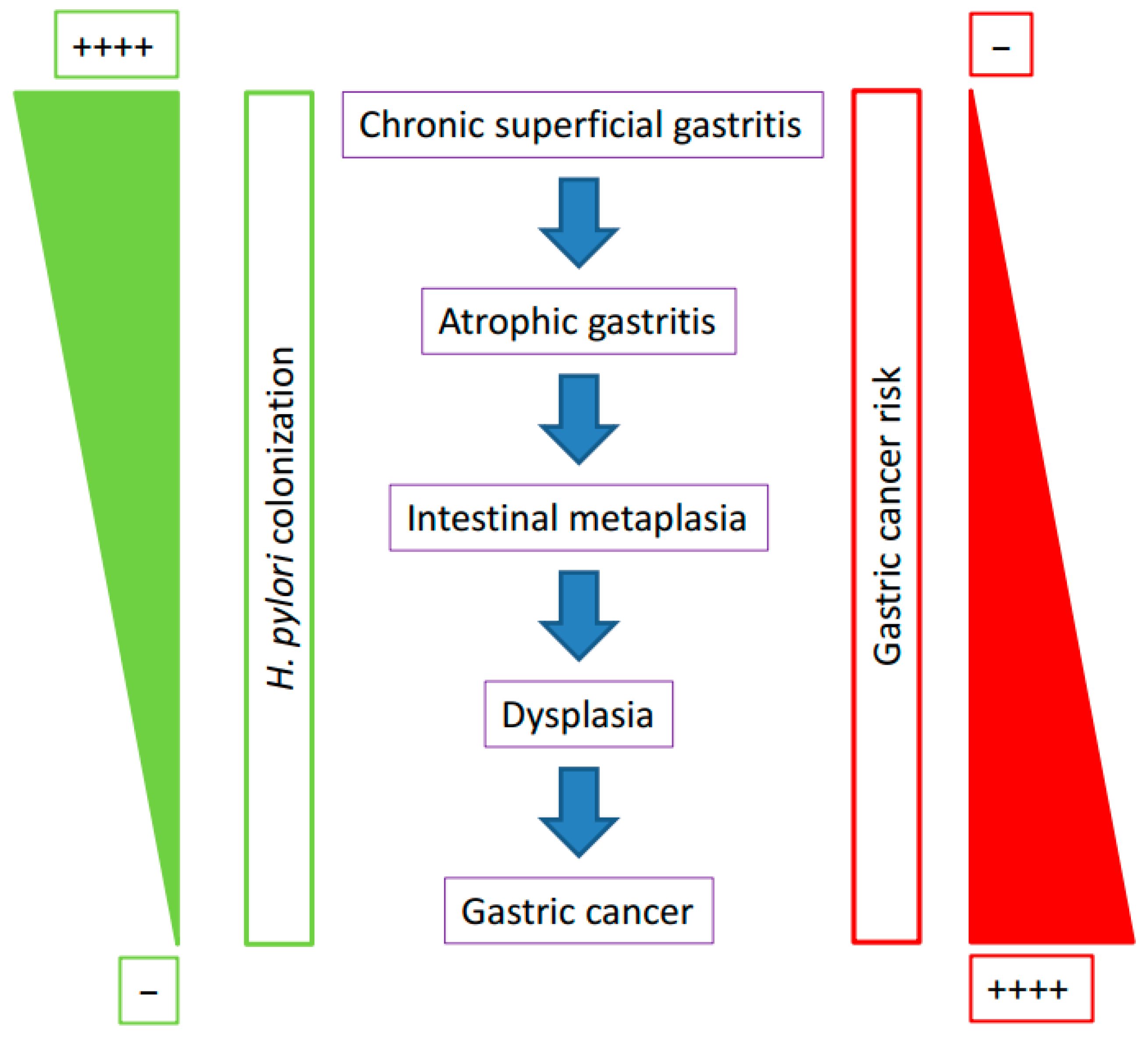
- Correa, P.; Cuello, C.; Duque, E.; Burbano, L.C.; Garcia, F.T.; Bolanos, O.; Brown, C.; Haenszel, W. Gastric cancer in Colombia. III Natural history of precursor lesions. J. Natl. Cancer Inst. 1976, 57, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process-First American Cancer Society Award lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar] [PubMed]
- Correa, P. Helicobacter pylori and gastric carcinogenesis. Am. J. Surg. Pathol. 1995, 19 (Suppl. 1), S37–S43. [Google Scholar] [PubMed]
- Relman, D.A. Microbiology: Learning about who we are. Nature 2012, 486, 1941–1995. [Google Scholar] [CrossRef] [PubMed]
- Llorca, L.; Perez Perez, G.; Urruzuno, P.; Martinez, M.J.; Iizumi, T.; Gao, Z.; Shon, J.; Chung, J.; Cox, L.; Simon-Soro, A.; et al. Characterization of the gastric microbiota in a pediatric population according to Helicobacter pylori status. Pediatr. Infect. Dis. 2017, 36, 1731–1778. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.; Newell, D.G.; Fullerton, F.; Yarnell, J.W.; Stacey, A.R.; Wald, N.; Sitas, F. Association between infection of Helicobacter pylori and risk of gastric cancer: Evidence from a prospective investigation. BMJ 1991, 302, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Nomura, A.; Stemmermann, G.N.; Chyou, P.H.; Kato, I.; Perez Perez, G.I.; Blaser, M.J. Helicobacter pylori infection and gastric carcinoma among Japanese American in Hawaii. N. Engl. J. Med. 1991, 325, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Hunt, R.H. The evolving epidemiology of Helicobacter pylori infection and gastric cancer. Can. J. Gastroenterol. 2003, 17 (Suppl. B), 18B–20B. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T. development of gastric cancer associated with Helicobacter pylori infection. Cancer Chemother. Pharmacol. 2004, 54 (Suppl. 1), S12–S20. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, F.; Takagi, S.; Kusuhara, H.; Iwao, E.; Yokoyama, Y.; Ikeda, Y. Induction of gastric ulcer and intestinal metaplasia in Mongolian gerbils infected with Helicobacter pylori. J. Gastroenterol. 1996, 31, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, F.; Takagi, S.; Iwao, E.; Yokoyama, Y.; Haga, K.; Hanada, S. Development of poor differentiated adenocarcinoma and carcinoid due to long term Helicobacter pylori colonization in Mongolian gerbils. J. Gastroenterol. 1999, 34, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Poh, A.R.; O’Donoghue, R.J.J.; Ernst, M.; Putoczki, T.L. Mouse models for gastric cancer: Matching models to biological questions. J. Gastroenterol. Hepatol. 2016, 31, 1257–1272. [Google Scholar] [CrossRef] [PubMed]
- Covacci, A.; Telford, J.L.; Del Giudice, G.; Parsonnet, J.; Rappuoli, R. Helicobacter pylori virulence and genetic geography. Science 1999, 284, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Higashi, H.; Tsutsumi, R.; Muto, S.; Sugiyama, T.; Azuma, T.; Asaka, M.; Hatekayama, M. SHP-2 tyrosine phosphatase as an intercellular target of Helicobacter pylori CagA protein. Science 2002, 295, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Powell, S.E.; Betel, D.; Shah, M.A. The gastric microbiome and its influence on gastric carcinogenesis. Current knowledge and ongoing research. Hematol. Oncol. Clin. N. Am. 2017, 31, 389–408. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, N.; Yuasa, H.; Tanaka, S.; Sawa, H.; Miura, M.; Matsui, A.; Higashi, H.; Musashi, M.; Iwabuchi, K.; Suzuki, M.; et al. Transgenic expression of Helicobacter pylori CagA induces gastrointestinal and hematopoietic neoplasms in mouse. Proc. Natl. Acad. Sci. USA 2008, 105, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Asaka, M. Helicobacter pylori infection and gastric cancer. Med. Electron Microsc. 2004, 37, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.; Valenzuela Valderrama, M.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and gastric cancer: Adaptive cellular mechanisms involved in diasese progression. Front. Microbiol. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Esch, T.; Stefano, G.B. Proinflammation: A common denominator or initiator of different pathophysiological disease processes. Med. Sci. Monit. 2002, 8, HY1–HY9. [Google Scholar] [PubMed]
- El-Omar, E.M.; Carrington, M.; Chow, W.; McColl, K.E.; Bream, J.H.; Young, H.A.; Herrera, J.; Lissowska, J.; Yuan, C.C.; Rothman, N.; et al. Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature 2000, 404, 398–402. [Google Scholar] [CrossRef] [PubMed]
- El-Omar, E.M. The importance of interleukin 1β in Helicobacter pylori associated disease. Gut 2001, 48, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.; Machado, J.C.; Pharoah, P.; Seruca, R.; Sousa, S.; Carvalho, R.; Capelinha, A.F.; Quint, W.; Caldas, C.; van Doorn, L.J.; et al. Helicobacter pylori and interleukin 1 genotyping: An opportunity to identify high-risk individuals for gastric carcinoma. J. Natl. Cancer Inst. 2002, 94, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Blaser, M.J. The human microbiome at the interface of health and disease. Nat. Rev. Genet. 2012, 3, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Matsumoto, A.; Tanaka, H.; Matsumura, I. Gastric microbiota: An emerging player in Helicobacter pylori-induced gastric malignancies. Cancer Lett. 2018, 414, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, N.; Jo, H.J.; Park, J.H.; Nam, R.H.; Seok, Y.J.; Kim, Y.R.; Kim, J.S.; Kim, J.M.; Kim, J.M.; et al. An appropriate cutoff value for determining the colonization of Helicobacter pylori by pyrosequencing method: Comparison with conventional methods. Helicobacter 2015, 20, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Parson, B.N.; Ijaz, U.; D’Amore, R.; Burkitt, M.D.; Eccles, R.; Lenzi, L.; Duckworth, C.A.; Moore, A.R.; Tiszlavicz, L.; Varro, A.; et al. Comparison of the human gastric microbiota in hypochlorhydria states arising as results of Helicobacter pylori-induced atrophic gastritis, autoimmune atrophic gastritis and proton pump inhibitor use. PLoS Pathog. 2017. [Google Scholar] [CrossRef] [PubMed]
- Thorell, K.; Bengtsson-Palme, J.; Liu, O.H.; Palacios Gonzales, R.V.; Nookaew, I.; Rabeneck, L.; Paszat, L.; Graham, D.Y.; Nielsen, J.; Lundin, S.B.; et al. In vivo analysis of the viable microbiota and Helicobacter pylori transcriptome in gastric infection and early stages of carcinogenesis. Infect. Immun. 2017, 85, e00031-17. [Google Scholar] [CrossRef] [PubMed]
- Shanks, A.M.; El-Omar, E.M. Helicobacter pylori infection, host genetics and gastric cancer. J. Dig. Dis. 2009, 10, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Shon, S.H.; Kim, N.; Jo, H.J.; Kim, J.; Park, J.H.; Nam, R.H.; Seok, Y.-J.; Kim, Y.-R.; Lee, D.H. Analysis of gastric body microbiota by pyrosequencing : Possible role of bacteria other than Helicobacter pylori in the gastric carcinogenesis. J. Cancer Prev. 2017, 22, 115–125. [Google Scholar] [CrossRef]
- Lofgren, J.L.; Whary, M.T.; Ge, Z.; Muthupalani, S.; Taylor, N.S.; Mobley, M.; Potter, A.; Varro, A.; Elbach, D.; Suerbaum, S.; et al. Lack of commensal flora in Helicobacter pylori-infected INS-GAS mice reduces gastritis and delays intraepithelial neoplasia. Gastroenterology 2011, 140, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Klymiuk, I.; Bilgilier, C.; Stadlmann, A.; Thannesberger, J.; Kastner, M.T.; Hogenauer, C.; Puspok, A.; Biowski-Frotz, S.; Schrutka-Kolbl, C.; Thallinger, G.G.; et al. The human gastric microbiome is predicated upon infection with Helicobacter pylori. Front. Microbiol. 2017, 8, 2508. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribero, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal Microbiome dysbiosis in gastric carcinogenesis. Gut 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Shi, J.; Fang, C.; Zou, X.; Huang, O. Gastric carcinoma in young (younger than 40 years) Chinese patients: Clinicopathology, family history, and postresection survival. Medicine 2016, 95, e2873. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.F.; Camargo, M.C.; Fraumeni, J.F.; Correa, P.; Rosenberg, P.S.; Rabkin, C.S. Age-specific trends in incidence of non cardi- gastric cáncer in the US adults. JAMA 2010, 303, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.F.; Rabkin, C.S.; Turner, N.; Fraumeni, J.F.; Rosenberg, P.S.; Camargo, M.C. The changing face of noncardia gastric cancer incidence among US non-Hispanic Whites. J. Natl. Cancer Inst. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Zhou, F.; Wang, J.; Zi, L. Risk factors for lymphonode metastasis of early gastric cancer in patients younger than 40. Medicine 2017, 96, 37. [Google Scholar] [CrossRef] [PubMed]
- Braga-Neto, M.B.; Gomes carneiro, J.; de Castro Barbosa, A.M.; Silva, I.S.; Maia, D.C.; Maciel, F.S.; Alves de Alcantara, R.J.; Vasconcelos, P.R.L.; Braga, L.L.B.C. Clinical characteristics of distal gastric cáncer in Young adults from Northeastern Brazil. BMC Cancer 2018, 18, 131. [Google Scholar] [CrossRef] [PubMed]
- Balakrish, M.; George, R.; Sharma, A.; Graham, D.Y.; Malaty, H.M. An investigation into recent increase in gastric cancer in the USA. Dig. Dis. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Carmel, R.; Johnson, C.S. Racial patterns in pernicious anemia. Early age of onset and increased frequency of intrinsic factor antibody in black women. N. Engl. J. Med. 1978, 298, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Neumann, W.L.; Coss, E.; Rugge, M.; Genta, R.M. Autoimmune atrophic gastritis-pathogenesis, pathology and management. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Yamashita, S.; Shimazu, T.; Lida, N.; Takeshima, H.; Nakajima, T.; Oda, I.; Nanjo, S.; Kusano, C.; Mori, A.; et al. Novel epigenetic markers for gastric cáncer risk stratification in individuals after Helicobacter pylori eradication. Gastric Cancer 2018. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.K.; Wong, I.O.L.; Cheung, K.S.; Yeung, K.F.; Chan, E.W.; Wong, A.Y.S.; Chen, L.; Wong, I.C.K.; Graham, D.Y. Effect of Helicobacter pylori treatment on incidence of gastric cancer in older individuals. Gastroenterology 2018. [Google Scholar] [CrossRef] [PubMed]
- Doorakkers, E.; Lagergren, J.; Engstrand, L.; Brusselaers, N. Helicobacter pylori eradication treatment and the risk of gastric adenocarcinoma in a western population. Gut 2018. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Perez Perez, G.I. Is There a Role for the Non-Helicobacter pylori Bacteria in the Risk of Developing Gastric Cancer? Int. J. Mol. Sci. 2018, 19, 1353. https://doi.org/10.3390/ijms19051353
Li J, Perez Perez GI. Is There a Role for the Non-Helicobacter pylori Bacteria in the Risk of Developing Gastric Cancer? International Journal of Molecular Sciences. 2018; 19(5):1353. https://doi.org/10.3390/ijms19051353
Chicago/Turabian StyleLi, Jackie, and Guillermo I. Perez Perez. 2018. "Is There a Role for the Non-Helicobacter pylori Bacteria in the Risk of Developing Gastric Cancer?" International Journal of Molecular Sciences 19, no. 5: 1353. https://doi.org/10.3390/ijms19051353
APA StyleLi, J., & Perez Perez, G. I. (2018). Is There a Role for the Non-Helicobacter pylori Bacteria in the Risk of Developing Gastric Cancer? International Journal of Molecular Sciences, 19(5), 1353. https://doi.org/10.3390/ijms19051353