Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage


Abstract
:1. Introduction
2. Results
2.1. Identification of TCP TFs in Chinese Cabbage
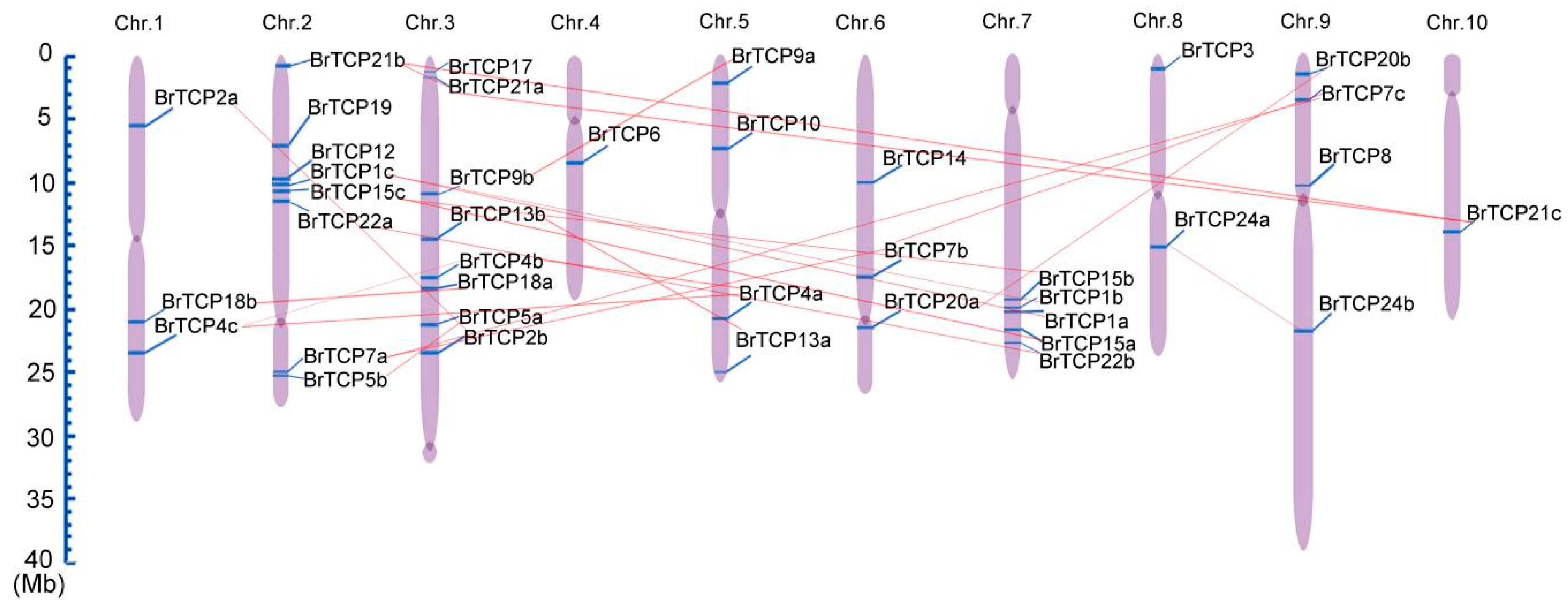
2.2. Chromosomal Location and Distribution of TCP Genes in Chinese Cabbage
2.3. Phylogeny and Gene Structure of the BrTCP Family
2.4. Conserved Domains and Motif Analysis of Chinese Cabbage
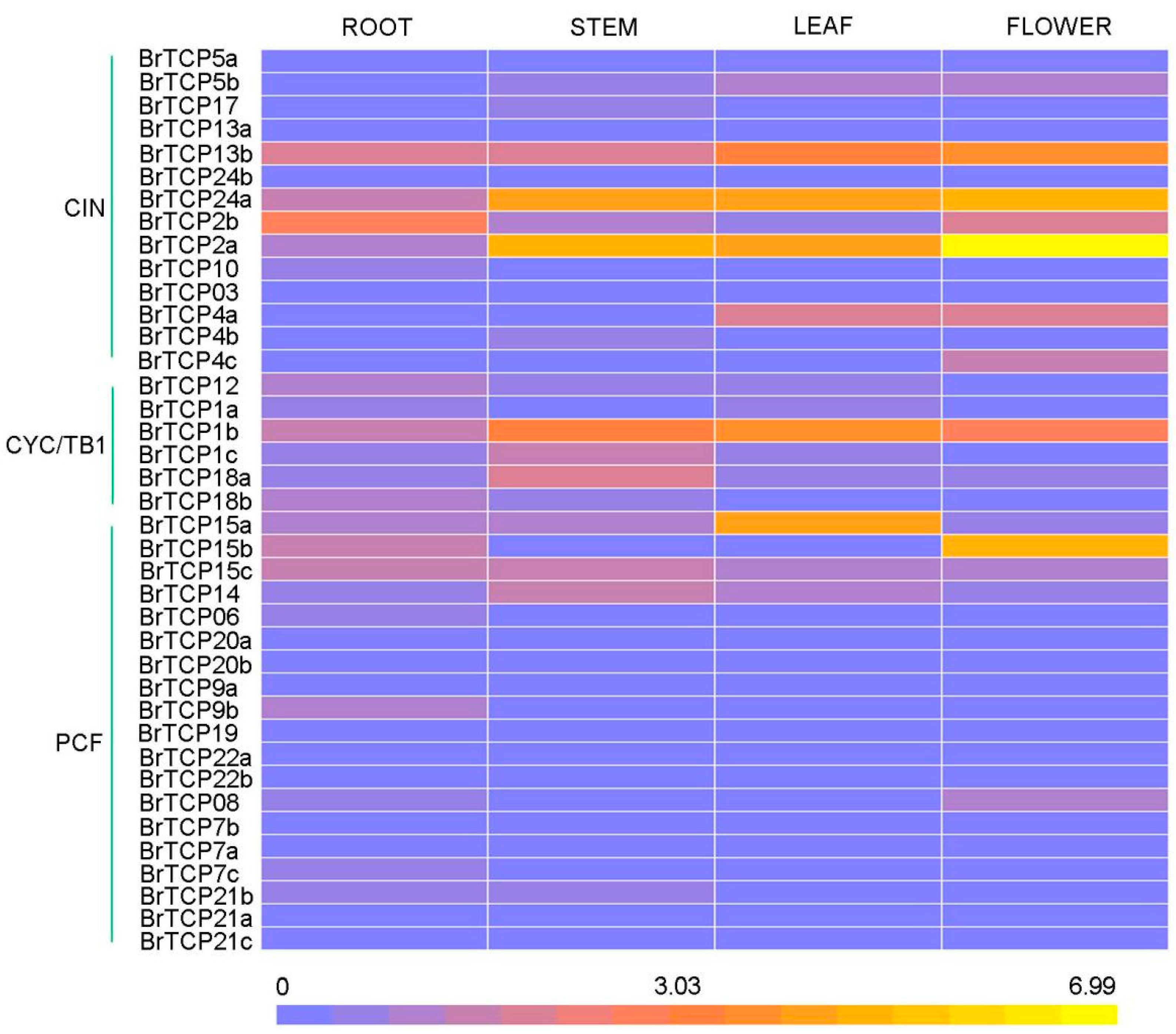
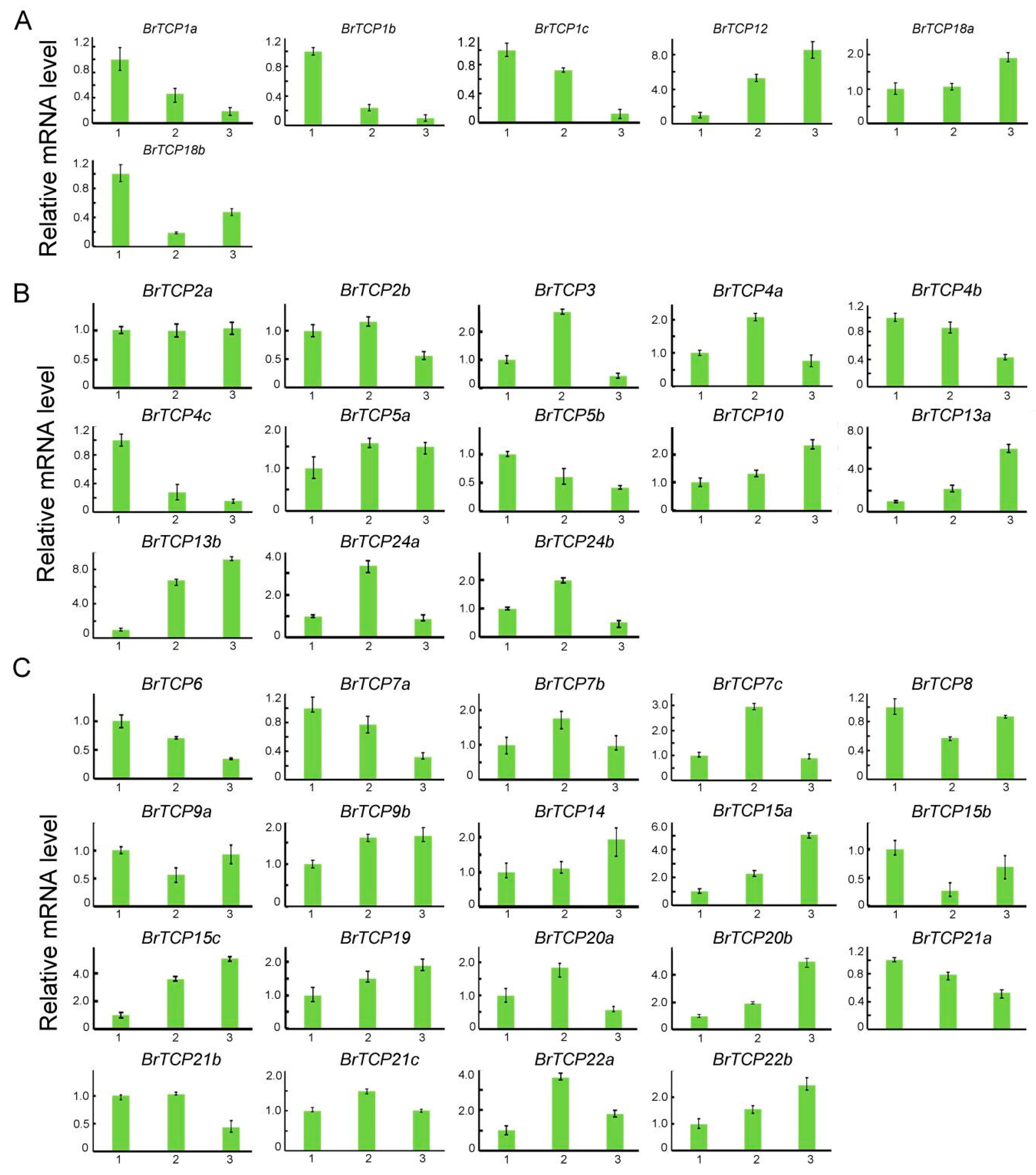
2.5. Expression Levels of TCP Genes in Chinese Cabbage
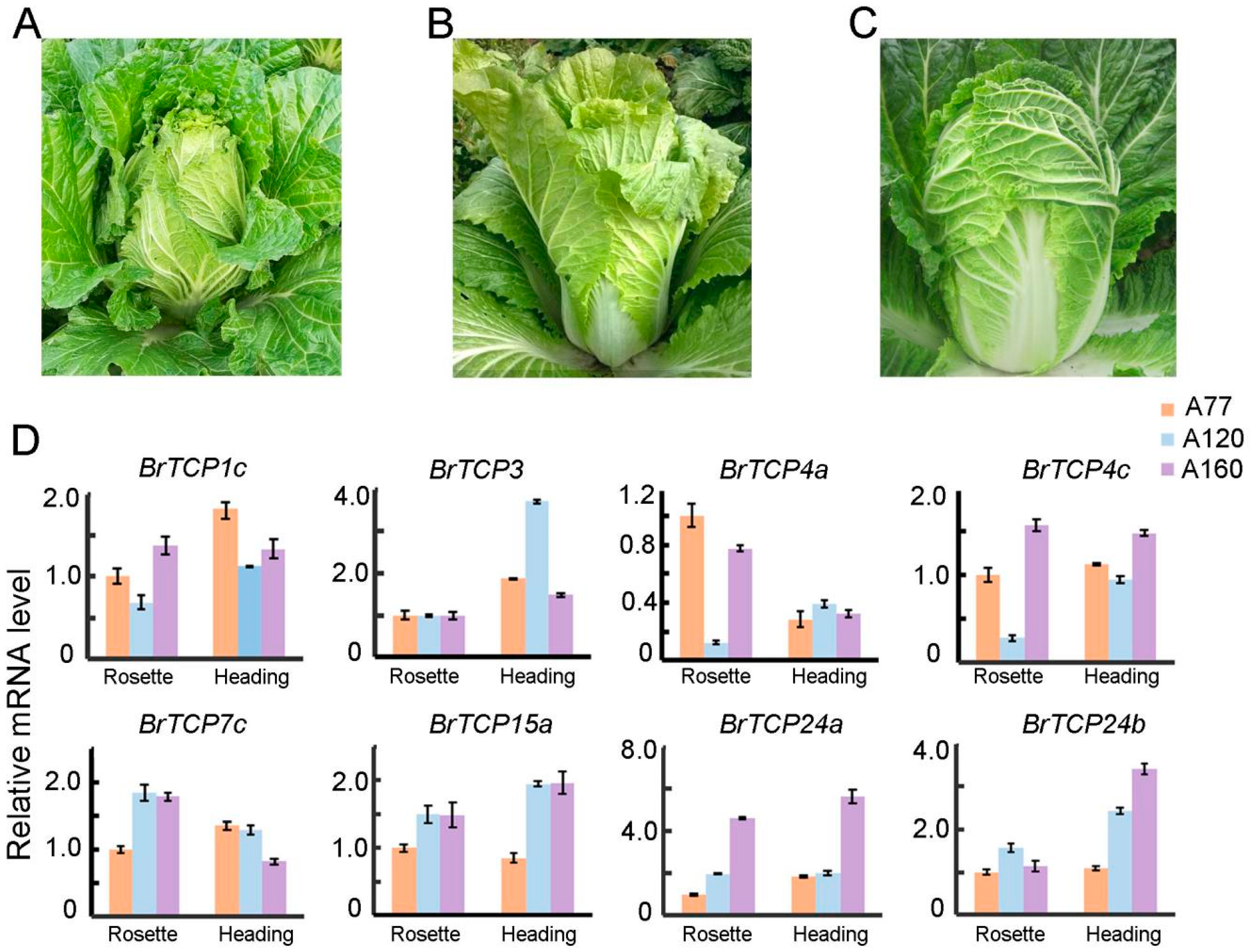
2.6. Associated Relationship between the Expression of BrTCPs and Heading of Chinese Cabbage
3. Discussion
4. Materials and Methods
4.1. Identification and Analysis of TCP Family Members in Chinese Cabbage
4.2. Chromosomal Location and Gene Structure
4.3. Phylogenetic Analysis and Analysis of Conserved Domains and Motif
4.4. Plant Materials
4.5. Total RNA Isolation and qRT-PCR Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Ohashi, Y. PCF1 and PCF2 specifically bind to cis elements in the rice proliferating cell nuclear antigen gene. Plant Cell 1997, 9, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. Cell Mol. Biol. 1999, 18, 215–222. [Google Scholar] [CrossRef]
- Luo, D.; Carpenter, R.; Copsey, L.; Vincent, C.; Clark, J.; Coen, E. Control of organ asymmetry in flowers of Antirrhinum. Cell 1999, 99, 367–376. [Google Scholar] [CrossRef]
- Cooper, A. A Case of Aneurism of the Carotid Artery. Med. Chir. Trans. 1809, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Ohashi, Y. DNA binding and dimerization specificity and potential targets for the TCP protein family. Plant J. Cell Mol. Biol. 2002, 30, 337–348. [Google Scholar] [CrossRef]
- Takeda, T.; Suwa, Y.; Suzuki, M.; Kitano, H.; Ueguchi-Tanaka, M.; Ashikari, M.; Matsuoka, M.; Ueguchi, C. The OsTB1 gene negatively regulates lateral branching in rice. Plant J. Cell Mol. Biol. 2003, 33, 513–520. [Google Scholar] [CrossRef]
- Vadde, B.V.L.; Challa, K.R.; Nath, U. The TCP4 transcription factor regulates trichome cell differentiation by directly activating GLABROUS INFLORESCENCE STEMS in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Wu, F.; Yu, X.; Bai, J.; Zhong, W.; He, Y. MicroRNA319a-targeted Brassica rapa ssp. pekinensis TCP genes modulate head shape in chinese cabbage by differential cell division arrest in leaf regions. Plant Physiol. 2014, 164, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Luo, Y.; Cai, Z.; Cao, X.; Hu, X.; Yang, J.; Luo, D. Functional diversity of CYCLOIDEA-like TCP genes in the control of zygomorphic flower development in Lotus japonicus. J. Integr. Plant Biol. 2013, 55, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Juntheikki-Palovaara, I.; Tahtiharju, S.; Lan, T.; Broholm, S.K.; Rijpkema, A.S.; Ruonala, R.; Kale, L.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Functional diversification of duplicated CYC2 clade genes in regulation of inflorescence development in Gerbera hybrida (Asteraceae). Plant J. Cell Mol. Biol. 2014, 79, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Nakabayashi, K.; Kamiya, Y.; Nambara, E. Transcription factor AtTCP14 regulates embryonic growth potential during seed germination in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2008, 53, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Balsemao-Pires, E.; Andrade, L.R.; Sachetto-Martins, G. Functional study of TCP23 in Arabidopsis thaliana during plant development. Plant Physiol. Biochem. 2013, 67, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Ng, S.; Carrie, C.; Duncan, O.; Low, J.; Lee, C.P.; Van Aken, O.; Millar, A.H.; Murcha, M.; Whelan, J. TCP transcription factors link the regulation of genes encoding mitochondrial proteins with the circadian clock in Arabidopsis thaliana. Plant Cell 2010, 22, 3921–3934. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Fujioka, S.; Blancaflor, E.B.; Miao, S.; Gou, X.; Li, J. TCP1 modulates brassinosteroid biosynthesis by regulating the expression of the key biosynthetic gene DWARF4 in Arabidopsis thaliana. Plant Cell 2010, 22, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Viola, I.L.; Uberti Manassero, N.G.; Ripoll, R.; Gonzalez, D.H. The Arabidopsis class I TCP transcription factor AtTCP11 is a developmental regulator with distinct DNA-binding properties due to the presence of a threonine residue at position 15 of the TCP domain. Biochem. J. 2011, 435, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, M.; Master, V.; Waites, R.; Davies, B. TCP14 and TCP15 affect internode length and leaf shape in Arabidopsis. Plant J. Cell Mol. Biol. 2011, 68, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.S.; Carvunis, A.R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.Z.; Chen, X.; Xiang, C.B.; Tang, N.; Zhang, Q.F.; Xiong, L.Z. Evaluation of seven function-known candidate genes for their effects on improving drought resistance of transgenic rice under field conditions. Mol. Plant 2009, 2, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Tyagi, A.K. OsTCP19 influences developmental and abiotic stress signaling by modulating ABI4-mediated pathways. Sci. Rep. 2015, 5, 9998. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.G.; Lee, G.H.; Kim, J.S.; Shim, E.J.; Park, Y.D. An insertional mutagenesis system for analyzing the Chinese cabbage genome using Agrobacterium T-DNA. Mol. Cells 2010, 29, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Kato, T. Studies on the head formation of Chinese cabbage. II. Relation between auxin and head formation. Agric. Hortic. 1958, 26, 771. (In Japanese) [Google Scholar]
- Gao, L.W.; Lyu, S.W.; Tang, J.; Zhou, D.Y.; Bonnema, G.; Xiao, D.; Hou, X.L.; Zhang, C.W. Genome-wide analysis of auxin transport genes identifies the hormone responsive patterns associated with leafy head formation in Chinese cabbage. Sci. Rep. 2017, 7, 42229. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, K.; Hasebe, M. Adaxial-abaxial polarity: The developmental basis of leaf shape diversity. Genesis 2014, 52, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liu, B.; Wu, J.; Cheng, F.; Wang, X. Genetic Variation and Divergence of Genes Involved in Leaf Adaxial-Abaxial Polarity Establishment in Brassica rapa. Front. Plant Sci. 2016, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. Brassica rapa Genome Sequencing Project, C. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Du, J.; Hu, S.; Yu, Q.; Wang, C.; Yang, Y.; Sun, H.; Yang, Y.; Sun, X. Genome-Wide Identification and Characterization of BrrTCP Transcription Factors in Brassica rapa ssp. rapa. Front. Plant Sci. 2017, 8, 1588. [Google Scholar] [CrossRef] [PubMed]
- Danisman, S. TCP Transcription Factors at the Interface between Environmental Challenges and the Plant’s Growth Responses. Front. Plant Sci. 2016, 7, 1930. [Google Scholar] [CrossRef] [PubMed]
- Li, S. The Arabidopsis thaliana TCP transcription factors: A broadening horizon beyond development. Plant Signal. Behav. 2015, 10, e1044192. [Google Scholar] [PubMed]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. A role of TCP1 in the longitudinal elongation of leaves in Arabidopsis. Biosci. Biotechnol. Biochem. 2010, 74, 2145–2147. [Google Scholar] [CrossRef] [PubMed]
- Bresso, E.G.; Chorostecki, U.; Rodriguez, R.E.; Palatnik, J.F.; Schommer, C. Spatial Control of Gene Expression by miR319-Regulated TCP Transcription Factors in Leaf Development. Plant Physiol. 2018, 176, 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mao, Y.; Yang, J.; He, Y. TCP24 modulates secondary cell wall thickening and anther endothecium development. Front. Plant Sci. 2015, 6, 436. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2014, 42, D1182–D1187. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome Location | CDS Length (bp) | Exon | Protein length (aa) | MW (Da) | PI |
|---|---|---|---|---|---|---|---|
| BrTCP1a | Bra004212 | A07:20728546–20729668 | 1026 | 2 | 341 | 38,786.3 | 6.5294 |
| BrTCP1b | Bra004097 | A07:20111066–20112206 | 1044 | 2 | 347 | 39,380 | 6.1198 |
| BrTCP1c | Bra034010 | A02:10223409–10224449 | 1041 | 1 | 346 | 38,511.8 | 5.477 |
| BrTCP2a | Bra013304 | A01:4882809–4883528 | 720 | 1 | 239 | 39,219.7 | 8.3134 |
| BrTCP2b | Bra012600 | A03:23315387–23316535 | 1149 | 1 | 382 | 42,262.9 | 7.6883 |
| BrTCP3 | Bra030952 | A08:1031146–1032183 | 1038 | 1 | 345 | 37,595.3 | 7.2706 |
| BrTCP4a | Bra027284 | A05:20829037–20830257 | 1221 | 1 | 406 | 44,035.4 | 7.5321 |
| BrTCP4b | Bra001579 | A03:17191721–17192944 | 1224 | 1 | 407 | 44,472.9 | 8.0355 |
| BrTCP4c | Bra021586 | A01:23505568–23506620 | 1053 | 1 | 350 | 38,203.2 | 7.3461 |
| BrTCP5a | Bra012990 | A03:21199353–21200453 | 1101 | 1 | 366 | 40,698.2 | 6.6787 |
| BrTCP5b | Bra029344 | A02:25211660–25212733 | 906 | 2 | 301 | 33,796.16 | 6.5 |
| BrTCP6 | Bra025528 | A04:8476440–8477114 | 675 | 1 | 224 | 24,724.6 | 7.816 |
| BrTCP7a | Bra029366 | A02:25051573–25052103 | 531 | 1 | 176 | 18,601 | 8.2872 |
| BrTCP7b | Bra009667 | A06:17073181–17073933 | 753 | 1 | 250 | 26,968.1 | 10.3355 |
| BrTCP7c | Bra026506 | A09:3299278–3299928 | 651 | 1 | 216 | 23,116.8 | 9.867 |
| BrTCP8 | Bra027886 | A09:10035227–10036411 | 1185 | 1 | 394 | 41,385.3 | 6.5677 |
| BrTCP9a | Bra004929 | A05:2539838–2540812 | 975 | 1 | 324 | 34,510.1 | 10.1796 |
| BrTCP9b | Bra000395 | A03:10937732–10938667 | 936 | 1 | 311 | 33,174.9 | 10.2854 |
| BrTCP10 | Bra018280 | A05:7148195–7149244 | 1050 | 1 | 349 | 38,727.4 | 6.8541 |
| BrTCP12 | Bra038350 | A02:10071368–10072479 | 1008 | 1 | 335 | 38,247.8 | 8.7635 |
| BrTCP13a | Bra039158 | A05:25073263–25074192 | 930 | 1 | 309 | 35,797.8 | 7.4173 |
| BrTCP13b | Bra001032 | A03:14476673–14479645 | 1617 | 4 | 538 | 59,180.4 | 7.2645 |
| BrTCP14 | Bra018126 | A06:10447686..10449086 | 1400 | 1 | 466 | 51,551 | 7.2085 |
| BrTCP15a | Bra004407 | A07:21777258–21778223 | 966 | 1 | 321 | 34,066.1 | 7.4651 |
| BrTCP15b | Bra003986 | A07:19409073–19409810 | 738 | 1 | 245 | 33,798.6 | 7.7239 |
| BrTCP15c | Bra007875 | A02:10484033–10484773 | 741 | 1 | 246 | 33,870.94 | 7.12 |
| BrTCP17 | Bra005967 | A03:1433460–1434200 | 741 | 1 | 246 | 27,367.4 | 7.0159 |
| BrTCP18a | Bra001710 | A03:18008822–18010603 | 1278 | 4 | 425 | 48,457.2 | 8.3237 |
| BrTCP18b | Bra037579 | A01:21234671–21236261 | 1284 | 4 | 428 | 48,910.3 | 7.1925 |
| BrTCP19 | Bra022568 | A02:7842256–7845540 | 894 | 2 | 297 | 30,182.5 | 5.419 |
| BrTCP20a | Bra025244 | A06:21530080–21531003 | 924 | 1 | 307 | 32,446.8 | 7.954 |
| BrTCP20b | Bra039096 | A09:1525195–1526031 | 837 | 1 | 278 | 29,671.6 | 6.5716 |
| BrTCP21a | Bra005984 | A03:1498581–1499285 | 705 | 1 | 234 | 24,263.2 | 10.8713 |
| BrTCP21b | Bra028662 | A02:1265168–1265857 | 690 | 1 | 229 | 23,957.7 | 10.2007 |
| BrTCP21c | Bra009338 | A10:14235005–14235715 | 711 | 1 | 236 | 24,733.4 | 9.2658 |
| BrTCP22a | Bra008001 | A02:11473562–11474686 | 1125 | 1 | 374 | 39,098.2 | 8.9956 |
| BrTCP22b | Bra016090 | A07:22761310–22762380 | 1071 | 1 | 356 | 37,206.3 | 7.4347 |
| BrTCP24a | Bra010789 | A08:15682126–15683052 | 927 | 1 | 308 | 34,410.5 | 7.1472 |
| BrTCP24b | Bra032365 | A09:22194917–22195873 | 957 | 1 | 318 | 35,327.5 | 8.1305 |
| PCF | CIN | CYC/TB1 | Total | |
|---|---|---|---|---|
| Arabidopsis thaliana | 13 | 8 | 3 | 24 |
| Brassica rapa (Chinese cabbage) | 19 | 14 | 6 | 39 |
| Brassica rapa (turnip) | 20 | 13 | 6 | 39 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Guan, X.; Liu, S.; Yang, M.; Ren, J.; Guo, M.; Huang, Z.; Zhang, Y. Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage. Int. J. Mol. Sci. 2018, 19, 847. https://doi.org/10.3390/ijms19030847
Liu Y, Guan X, Liu S, Yang M, Ren J, Guo M, Huang Z, Zhang Y. Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage. International Journal of Molecular Sciences. 2018; 19(3):847. https://doi.org/10.3390/ijms19030847
Chicago/Turabian StyleLiu, Yan, Xiaoyu Guan, Shengnan Liu, Meng Yang, Junhui Ren, Meng Guo, Zhihui Huang, and Yaowei Zhang. 2018. "Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage" International Journal of Molecular Sciences 19, no. 3: 847. https://doi.org/10.3390/ijms19030847
APA StyleLiu, Y., Guan, X., Liu, S., Yang, M., Ren, J., Guo, M., Huang, Z., & Zhang, Y. (2018). Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage. International Journal of Molecular Sciences, 19(3), 847. https://doi.org/10.3390/ijms19030847